Запутанная история возникновения цветового зрения
Все человекообразные и другие обезьяны Старого Света (африканские и азиатские) обладают трихроматическим цветовым зрением и имеют три гена опсинов, тогда как американские обезьяны, а также грызуны и другие млекопитающие обычно обладают дихроматическим зрением и имеют два гена опсинов. Глядя на рис. 4.4, можно сделать вывод, что полноценное цветовое зрение возникло у предка приматов Старого Света после разделения линий, обитающих в Старом и Новом Свете. Кроме того, поскольку приматы Старого Света имеют третий зрительный пигмент колбочек, соответствующий ген опсина также должен был появиться после этого разделения. Это говорит о том, что наша с вами способность воспринимать цвета появилась у нашего древнего предка в Старом Свете, а не возникла независимо в ходе более поздней эволюции гоминидов.
Наличие у других млекопитающих (белок, кошек, собак и др.) лишь двух опсинов и дихроматического зрения означает, что общий предшественник всех млекопитающих обладал дихроматическим зрением. Но, прежде чем мы сделаем вывод о том, что полноценное цветовое зрение приматов является их «уникальным усовершенствованием», нужно исследовать зрение других позвоночных. Тут‑то и возникает загвоздка. Прекрасным цветовым зрением обладают птицы, а также рептилии и многие рыбы, в частности золотые рыбки. У этих животных обнаружено не менее четырех генов опсинов. А у некоторых более примитивных позвоночных, таких как миноги, найдено пять генов опсинов. Это означает, что цветовое зрение возникло на очень ранних этапах эволюции позвоночных животных – до их разделения на челюстных и бесчелюстных. Поэтому, если рассматривать эволюционное древо позвоночных животных в целом, выясняется, что не относящиеся к приматам млекопитающие в какой‑то момент потеряли гены опсинов и свою способность цветового зрения. Из картины распределения этих признаков у позвоночных можно сделать вывод, что в ходе эволюции способность к цветовому зрению сначала была широко распространена, но потом у предков млекопитающих она исчезла и возродилась вновь у предков приматов Старого Света.
Возможно, вы удивитесь: если цветовое зрение играет такую важную роль, как же оно могло исчезнуть? Наиболее правдоподобное объяснение связано с ночным образом жизни млекопитающих. Первые млекопитающие были мелкими животными и вели скрытный, ночной образ жизни в экосистемах, хозяевами которых были более крупные животные, такие как динозавры. При ночном образе жизни не нужно различать цвета, а нужно иметь возможность видеть в полумраке и в темноте. В следующей главе мы рассмотрим несколько примеров того, как подобные изменения образа жизни приводят к потере генов, включая гены цветового зрения.
Мы достаточно точно можем указать, когда в ходе эволюции приматов появился наш третий ген опсина. Остается ответить на вопрос о том, как это произошло. Как этот новый ген смог расширить диапазон цветового зрения? Этапы эволюции цветового зрения можно проследить по последовательности гена этого опсина. Итак, рассмотрим последовательности опсинов, проанализируем отличия двух наших красно‑зеленых опсинов друг от друга и попробуем понять, какие из этих различий обеспечили их чувствительность к разным цветам. Общей закономерностью эволюции цветового зрения является «настройка» опсинов при адаптации к специфическим условиям. Сначала я расскажу вам, как настроены опсины человека, и представлю доказательства того, что они действительно полезны (адаптивны) для приматов. Затем я приведу несколько примеров того, как опсины разных видов организмов настроились на восприятие света с другой длиной волны при адаптации этих организмов к различным средам обитания и в ответ на различные стимулы.
Красное и зеленое
Крысы, мыши, белки, кролики, козы и другие млекопитающие имеют единственный MWS/LWS‑опсин с максимумом поглощения при длине волны 510–550 нм. Этот опсин кодируется единственным геном. Напротив, человек обладает двумя опсинами (MWS и LWS), которые кодируются двумя генами на X‑хромосоме, расположенными в тандеме «голова к хвосту». Последовательности ДНК этих двух опсинов совпадают на 98 %. Столь большое сходство и ближайшее соседство этих генов говорит о том, что они возникли в результате удвоения единственного гена опсина MWS/LWS у какого‑то примата‑предка. Удвоение генов – довольно распространенная форма изменения последовательности ДНК; многие наши гены в ходе эволюции приобрели по несколько копий. Рост числа копий гена увеличивает количество информации, на которую может влиять естественный отбор, и достаточно часто функции этих копий со временем начинают различаться. Именно это произошло с двумя олеинами на X‑хромосоме.
Наша пара опсинов, как и опсины других приматов с три‑хроматическим зрением, в наибольшей степени стимулируется светом с длиной волны 530 нм (зеленый) и 560 нм (красный) – это их максимумы поглощения. Изучение функциональных свойств опсинов показало, что их спектр поглощения достаточно легко изменить путем замены определенных аминокислотных остатков. То, что у всех приматов с трихроматическим зрением максимумы поглощения белков сохранились на длине волны 530 и 560 нм, означает, что это свойство белков поддерживается естественным отбором.
Последовательности зеленого и красного пигментов различаются всего 15 аминокислотными остатками. Заменяя одну аминокислоту на другую и анализируя результат, ученые смогли установить, какие именно аминокислотные остатки отвечают за особые свойства каждого из пигментов.
По‑видимому, основной вклад в различие в максимумах поглощения красного и зеленого пигментов вносят аминокислоты в положениях 180, 277 и 285. Аминокислоты, находящиеся в этих позициях в двух пигментах, а также их вклад в сдвиг максимума поглощения, представлены в табл. 4.1.
Таблица 4.1. Аминокислоты в ключевых позициях человеческих опсинов

Исследования показывают, что после удвоения гена пигмента MWS/LWS у нашего далекого предка две образовавшиеся копии стали функционировать по‑разному (одна настроилась на восприятие света с диной волны 530 нм, другая – 560 нм) главным образом в результате изменения аминокислотных остатков в этих трех ключевых позициях (рис. 4.5).

Рис. 4.5. Удвоение и тонкая настройка гена опсина у человекообразных обезьян. У общего предшественника обезьян и человекообразных обезьян Старого Света произошло удвоение гена опсина. Со временем в двух копиях возникли мутации, которые привели к настройке двух опсинов на поглощение красного или зеленого цвета. Сохранению этих мутаций способствовал естественный отбор. Рисунок Лианн Олдс.
Удвоение гена красно‑зеленого зрительного пигмента произошло после разделения приматов Старого и Нового Света. По‑видимому, это случилось 30 млн или 40 млн лет назад, вскоре после разделения африканского и южноамериканского континентов. По‑видимому, последовавшие за удвоением гена эволюционные изменения аминокислот в трех ключевых позициях предоставляли существенные преимущества. Теперь в Азии и Африке живут обезьяны только с трихроматическим зрением. Если в период возникновения цветового зрения рядом с этими обезьянами обитали и другие, не обладавшие такой способностью (что кажется вполне вероятным), то они и их потомки уже вымерли.
Конечно, мы не были свидетелями этих событий, произошедших 30 млн или 40 млн лет назад, и кто‑то может возразить, что все сказанное выше – всего лишь гипотезы. Однако важная роль цветового зрения у приматов – неоспоримый факт. В частности, у диких обезьян очень редко встречается дальтонизм. У людей цветовая слепота – достаточно распространенное явление: около 8 % мужчин белой расы не различают некоторых цветов из‑за аномалии генов красно‑зеленых опсинов на X‑хромосоме, однако в дикой природе это большая редкость. При исследовании 3153 макак дальтонизм был обнаружен лишь у трех особей (менее 0,1 %). Высокий уровень распространения дальтонизма у людей (у которых цветовое зрение, по крайней мере в настоящее время, находится под очень слабым влиянием естественного отбора) и низкий – у диких макак говорит о том, что естественный отбор поддерживает способность цветового восприятия у этих обезьян и у других видов, обладающих трихроматическим зрением.
Второе свидетельство в пользу большой экологической значимости трихроматического зрения у приматов дали наблюдения за тем, как выбирают пищу ди‑ и трихроматические приматы в природных условиях. Питер Лукас из Университета Гонконга, Натаниэль Домини (теперь работает в Университете Калифорнии в Санта‑Крузе) и их коллеги предприняли подробное исследование пищевых привычек и предпочтений колобусов и шимпанзе в Уганде, лемуров на Мадагаскаре и паукообразных обезьян в Коста‑Рике. Ученые обнаружили, что животные, обладающие трихроматическим зрением, отдают явное предпочтение более нежным красноватым листьям, которые содержат больше белка. Большинство исследованных приматов питались еще и фруктами, и цвет фруктов также имел для них значение. Однако Лукас и Домини считают, что полноценное цветовое зрение играет более важную роль в выборе листьев, особенно когда фруктов мало или они еще не поспели.
Таким образом, способность воспринимать красный и зеленый цвет, по‑видимому, дает определенные преимущества. Однако красный и зеленый – это только часть видимого спектра, наиболее важная для обитателей лесов. Но животные населяют самые разные уголки планеты, в том числе моря, где способность различать красный и зеленый цвета абсолютно бесполезна.
В глубоком синем море
На большие морские глубины солнечный свет проникает сквозь толщу воды, поэтому там темно. За зрение в слабом рассеянном свете отвечают фоторецепторы палочек, а опсин этих клеток называется родопсином. Родопсин человека и большинства наземных млекопитающих настроен таким образом, чтобы максимально поглощать свет с длиной волны около 500 нм.
На глубину около 200 м проникает лишь свет из узкого синего диапазона с длиной волны около 480 нм. Интересно, что родопсины глубоководных рыб и дельфинов сдвинуты в синюю область, то есть их максимум поглощения на 10–20 нм ниже максимума поглощения наземных млекопитающих. Ученые активно изучали тонкую настройку родопсина таких океанских жителей, как афалина, обыкновенный дельфин, черный дельфин, атлантический ремнезуб, солнечник и различные угри. Чтобы установить влияние отдельных аминокислотных остатков на различия между родопсинами разных видов животных, одни аминокислоты или их группы заменяли другими, обнаруженными у других видов. В частности, у афалины за сдвиг максимума поглощения родопсина на 10 нм в синюю область по сравнению с родопсинами наземных млекопитающих отвечают аминокислоты в положениях 83, 292 и 299 (рис. 4.6). Максимум поглощения родопсина ремнезуба еще сильнее сдвинут в синюю область (до 484 нм), и этот родопсин отличается от родопсина дельфина только аминокислотой в положении 299.
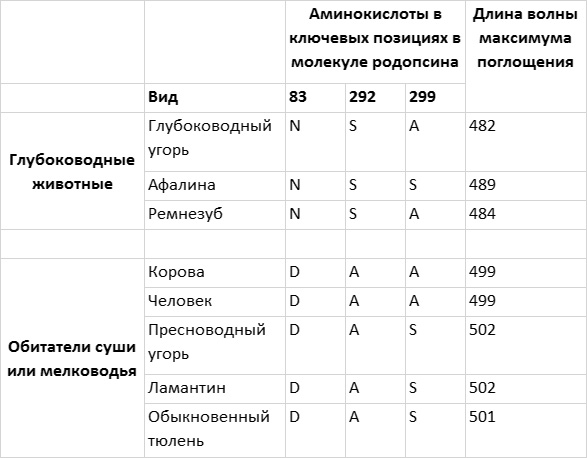
Рис. 4.6. Настройка родопсина зависит от глубины, на которой обитают морские животные. Глубоководные рыбы и китообразные имеют родопсин, максимум поглощения которого сдвинут в синюю область спектра по сравнению с родопсинами животных, обитающих на суше или на мелководье. У животных, обитающих на одинаковой глубине, часто в ключевых позициях в последовательности родопсина стоят одни и те же аминокислоты. Рисунок Джейми Кэрролл.
Связь «синего сдвига» родопсина с адаптацией к жизни в морских глубинах подкрепляется данными анализа родопсинов глубоководных и пресноводных угрей. Родопсин угря, обитающего на большой глубине, характеризуется сдвигом максимума поглощения в синюю область и в ключевых положениях имеет точно те же аминокислотные остатки, что и родопсин ремнезуба. Родопсин пресноводного угря имеет максимум поглощения, близкий к таковому у наземных млекопитающих, а в ключевых позициях в его последовательности находятся те же аминокислотные остатки, что и у ламантина и тюленя – двух обитающих на мелководье млекопитающих с типичным вариантом родопсина.
Эта поразительная связь между «синим сдвигом» родопсина и жизнью на большой глубине полна глубокого смысла. Резонно предположить, что естественный отбор способствовал настройке родопсина в соответствии со средой обитания. Однако существует еще более важная причина подозревать влияние естественного отбора, и она становится ясна при рассмотрении эволюционных связей между видами животных, перечисленных на рис. 4.6. Дельфины и ремнезубы относятся к отряду китообразных – млекопитающих, которые произошли от сухопутного предка, вернувшегося в водную стихию. Это может показаться удивительным, но ближайшими современными родственниками китообразных являются гиппопотамы, олени, коровы, свиньи и верблюды. Это следует из результатов анализа последовательностей SINE и LINE, а также других последовательностей ДНК. Поскольку родопсины наземных родственников китообразных настроены на длину волны 500 нм, можно с полным основанием заключить, что родопсин дельфинов и китов изменился после того, как их эволюционная ветвь отделилась от ветви других млекопитающих.
Однако угри – это рыбы, и их эволюционная ветвь отделилась от ветвей других позвоночных несколько сотен миллионов лет назад. Это означает, что одинаковые по своей сути различия между глубоководными и пресноводными угрями и морскими и наземными млекопитающими возникли независимо. Если в одинаковых позициях в каком‑то белке у двух или нескольких видов животных при адаптации к определенным условиям появляются одни и те же аминокислотные остатки (как у китообразных и глубоководного угря в нашем примере), это говорит о работе естественного отбора. Замечу, что пример эволюции родопсина – лишь один из множества случаев, когда эволюция воспроизводит саму себя (мы подробнее поговорим об этом в шестой главе).
Теперь перейдем от красного, зеленого и синего к невидимым для нас цветам – и к целому миру взаимоотношений между животными, связанных с ультрафиолетовым зрением.
За пределами радуги
Примерно через 12 лет после выхода в свет книги «О происхождении видов» Дарвин опубликовал «Происхождение человека и половой отбор». Это был первый труд Дарвина, посвященный эволюции человека, но, возможно, еще важнее то, что Дарвин впервые обратил внимание на различную роль полов в эволюции признаков; он называл этот процесс «половым отбором». И сам половой отбор, и развитая Дарвином теория полового отбора известны широкой публике гораздо хуже, чем его теория естественного отбора, однако биологи считают половой отбор одним из наиболее важных и интересных механизмов в эволюции животных. Половой отбор – это арена, на которой «наилучшая приспособленность» оказывается напрямую связана с успешностью спаривания.
Дарвин был увлечен разнообразием птичьей окраски – он посвятил много времени изучению этого удивительного явления и оставил многостраничные описания роскошного оперения самых разных видов птиц. Особенно его интересовало, каким образом предпочтения самок могут привести к эволюции причудливого оперения самцов, такого как павлиний хвост. Еще со времен Дарвина птицы остаются излюбленным объектом биологов, изучающих половой отбор. Но до недавних пор подобные исследования имели один и тот же изъян: окраску птиц оценивали люди. Я уже говорил о том, что мы видим мир совсем не таким, каким его видят другие млекопитающие, и не таким, каким его или друг друга видят птицы. Многие птицы способны видеть в ультрафиолетовом свете и различать цвета, не существующие для нас, и эта способность играет важную роль в спаривании, поиске пищи и даже вскармливании птенцов.
У многих птиц опсины настроены на восприятие ультрафиолетового света, и на их оперении есть отметины, отражающие свет в ультрафиолетовой части спектра (рис. D‑K).


Длина волны ультрафиолетового света меньше 400 нм, то есть меньше длины волны фиолетового света, и этот свет регистрируется коротковолновым опсином. Человеческий опсин SWS настроен на длину волны 417 нм. У некоторых видов птиц опсины SWS настроены на длину волны около 370 нм, поэтому эти птицы видят в ультрафиолетовом свете. Другие птицы имеют коротковолновые опсины, настроенные на фиолетовую область спектра, с максимумом поглощения около 405 нм, и эти птицы, как и люди, не могут воспринимать ультрафиолетовый свет. Лабораторные исследования помогли установить, какие изменения опсинов SWS отвечают за чувствительность птиц к ультрафиолету.
Одна специфическая позиция, а именно позиция 90 в коротковолновом опсине, напрямую связана со способностью птиц воспринимать фиолетовый или ультрафиолетовый свет. Те птицы, у которых в этом положении находится аминокислота серин, видят в фиолетовой области спектра, а те, у которых в этом месте располагается цистеин, видят в ультрафиолетовом диапазоне (рис. 4.7).

Рис. 4.7. Эволюция способности птиц к восприятию ультрафиолетового света. Восприятие птицами фиолетового или ультрафиолетового света в значительной степени зависит от того, какая аминокислота – серин (S) или цистеин (C) – находится в позиции 90 в молекуле коротковолнового опсина. Замена аминокислоты в этой позиции в разных отрядах птиц происходила не менее четырех раз. Рисунок Джейми Кэрролл.
Седзо Йокояма, один из ведущих специалистов в области цветового зрения, и его коллеги из Университета Эмори (Джорджия) напрямую показали, что замена серина на цистеин делает пигмент, чувствительный к фиолетовому свету, чувствительным к ультрафиолетовому свету, а замена цистеина на серин в пигменте, чувствительном к ультрафиолету, делает его восприимчивым к фиолетовому свету. Разница в одной‑единственной аминокислоте обеспечивает сдвиг максимума поглощения на 35–38 нм, что чрезвычайно много. Эти исследования показывают, что одна замена способна изменить функцию опсина, и следовательно, эволюция опсина, чувствительного к фиолетовому или ультрафиолетовому свету, может быть относительно простым, одностадийным процессом.
Птицы, обладающие способностью видеть в ультрафиолетовом свете, относятся к девяти семействам из четырех разных отрядов. На основании их родственных связей можно сделать вывод, что у птиц способность видеть в ультрафиолетовом свете возникала в ходе эволюции независимым образом не менее четырех раз. Во всех отрядах, к которым относятся птицы, способные воспринимать ультрафиолет, есть также птицы с фиолетовым типом SWS‑опсина. Это означает, что мутация, заменившая серин на цистеин в молекуле фиолетового пигмента, происходила в ходе эволюции несколько раз.
В ситуации с ультрафиолетовым зрением эволюция вновь повторяет саму себя. Это убедительное доказательство того, что на гены опсина оказал воздействие отбор. Однако в данном случае это мог быть половой отбор, поскольку многочисленные исследования показывают, что на выбор партнера у птиц, способных видеть в ультрафиолетовой области спектра, влияет окраска, различимая только в ультрафиолетовом диапазоне (рисунки D‑K цветной вкладки).
Например, самки скворцов выбирают себе партнера по окраске оперения, видимой только в ультрафиолетовом свете, а не в видимой области спектра, которую воспринимает человек. Это открытие было сделано в ходе эксперимента, когда птиц помещали в специальные камеры, позволяющие фильтровать свет с определенной длиной волны. При наличии ультрафиолетового света самки по‑другому оценивали самцов, причем явно отдавали предпочтение тем, у которых кончики перьев на груди несут ультрафиолетовые отметины.
Аналогичным образом самцы лазоревки (синей синицы) отличаются от самок тем, что голубая шапочка у них на голове отражает свет в ультрафиолетовом диапазоне (рисунки D‑G цветной вкладки). В лабораторных экспериментах самки отдают предпочтение тем самцам, шапочка которых наиболее сильно отражает ультрафиолет. Именно поэтому ученые из Бристольского университета, проводившие это исследование, назвали свою статью «Синие синицы на самом деле ультрафиолетовые».
Способность видеть в ультрафиолетовом свете помогает не только в выборе партнера. Недавно было доказано, что у восьми видов птиц рты птенцов, особенно края клювов, отражают свет в ультрафиолетовом диапазоне. Это означает, что вернувшиеся к гнезду родители даже в темноте могут видеть рты своих детенышей. Кроме того, сильные птенцы, по‑видимому, сильнее отражают ультрафиолетовый свет, так что в конкуренции между птенцами наиболее приспособленными оказываются те, чьи клювы заметнее для кормящих родителей.
Есть данные и о том, как способность воспринимать ультрафиолетовый свет используется для охоты. Лазоревки используют свое ультрафиолетовое зрение для поиска гусениц, имеющих маскирующую окраску в видимой области спектра. Хищная птица пустельга охотится на мышей‑полевок, отслеживая участки наибольшего скопления мышей по их экскрементам, которые отражают ультрафиолетовые лучи.
Способностью видеть в ультрафиолетовом свете обладают, конечно же, не только птицы. Известны некоторые виды рыб, амфибий, рептилий и млекопитающих (таких как летучие мыши), которые видят в ультрафиолетовом свете, и у каждого из этих видов животных есть опсин SWS, настроенный на длину волны 360–370 нм. Широкое распространение этой способности говорит о том, что она подходит для самых разных целей. Это один из общих принципов эволюции: одно новшество создает возможности для эволюции дополнительных новшеств. В завершении данной главы я вновь вернусь к обезьянам колобусам и расскажу о том дополнительном наборе новшеств, которые появились у этих животных вдобавок к умению находить самые питательные листья. Это еще один яркий пример развития новых способностей на основе «старых» генов.
Жвачные обезьяны
В отличие от большинства приматов, которые употребляют в пищу в основном фрукты и насекомых, обезьяны колобусы питаются листьями. Переваривать листья колобусу, как и другим жвачным животным, помогают бактерии, населяющие передний отдел кишечника – один из отделов многокамерной пищеварительной системы. Подобно коровам и другим жвачным животным колобус извлекает питательные вещества, расщепляя бактерии с помощью различных ферментов. Один из важных ферментов, рибонуклеаза, образуется в поджелудочной железе и секретируется в тонкую кишку, где расщепляет РНК. Этот фермент помогает высвободить большое количество азота, заключенного в РНК ферментирующих бактерий. В поджелудочной железе колобуса и обычных жвачных животных содержится больше рибонуклеазы, чем у других млекопитающих, поэтому ученые заинтересовались эволюцией этого пищеварительного фермента.
Выяснилось, что большинство млекопитающих, в том числе обезьяны, имеют лишь одну копию гена панкреатической рибонуклеазы, тогда как у колобуса таких генов три. В ходе эволюции этих обезьян ген рибонуклеазы удвоился. Жианши Цанг из Университета Мичигана подробно изучил рибонуклеазы и кодирующие их гены и пришел к выводу, что одна копия гена кодирует фермент, фактически идентичный ферменту других видов обезьян, но две другие, «новые» копии изменились и подстроились под нужды пищеварительной системы колобуса.
Всего в последовательностях двух новых копий рибонуклеазы обнаружено соответственно десять и тринадцать аминокислотных замен по сравнению с традиционной версией фермента. Одно из наиболее важных отличительных свойств новых белков состоит в том, что они обладают повышенной активностью в более кислой среде. Эта особенность согласуется с различиями в устройстве системы пищеварения колобусов и других приматов.
Но это открытие – не единственное доказательство участия естественного отбора в эволюции рибонуклеазы. Еще более весомые доказательства получены в результате анализа последовательности генов и белков новых рибонуклеаз. Вспомните, в предыдущей главе мы говорили о том, что большинство изменений в ДНК являются синонимичными. Однако в двух новых версиях гена рибонуклеазы соотношение несинонимичных и синонимичных изменений очень высокое (примерно 4:1, тогда как для других белков это соотношение составляет 1:5). Это является прекрасным доказательством того, что специфические изменения белка были поддержаны естественным отбором.
Изобретаем и адаптируемся
Эволюция цветового зрения и пищеварительных ферментов – лишь два из огромного числа примеров увеличения объема генетической информации и ее настройки в ходе приспособления видов к новым экологическим нишам. Удвоение генов и их тонкая настройка под действием естественного отбора происходят повсеместно. Большинство наших генов относятся к семействам генов, расширившимся в ходе эволюции. Случайное удвоение генов или групп генов происходит достаточно часто. Многие люди различаются между собой по количеству копий генов.
Поскольку в момент своего появления новые копии генов являются избыточными, ненужными, со временем лишь часть их сохраняется и подвергается функциональным изменениям, о которых я рассказывал на примере опсинов и рибонуклеазы. Сохранение и настройка генов – это видоспецифический процесс, который зависит от вероятности, отбора и времени. Разная судьба генов приводит к разнице в количестве генов у разных видов, а также, что еще более важно, к физиологическим и другим различиям между видами. Мы увидели, что смена образа жизни, например освоение океанских глубин или переход на лиственную диету, сопровождается характерными изменениями генов, задействованных в этих процессах.
Однако приобретение и тонкая настройка генов – лишь одна сторона эволюционной адаптации. У этого процесса есть вторая сторона, которая также отражена в ДНК. Когда образ жизни вида изменяется по сравнению с образом жизни его предков, какие‑то функции генов становятся ненужными, и такие гены начинают разрушаться. Если мы застаем ген в таком состоянии, он может многое рассказать об эволюционном изменении вида. Об этом мы и поговорим в следующей главе.

Латимерия. Фотография сделана с помощью глубоководного аппарата IAGO, сконструированного Юргеном Шоером и Хансом Фрике.
Глава 5
Дата добавления: 2018-09-22; просмотров: 306; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!