Уровня СМФП в области левой (s.) и правой (d.) миндалин (Amygd.) в период 25 страница
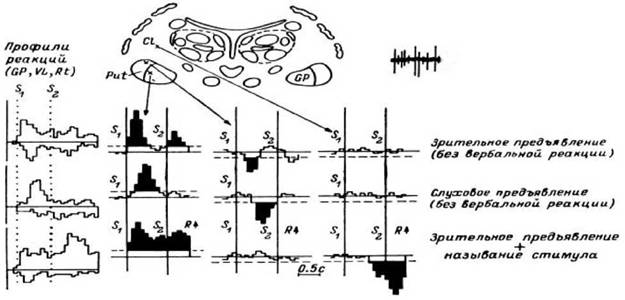
Рис. 33. Реакции нейронов мозга человека в ответ на предъявление зрительных и слуховых стимулов.
Профили реакции всех обследованных нейронных популяций (слева) и ПСГ, вычисленные для отдельных нейронных популяций подкорковых образований мозга в ответ на зрительное (верхний ряд), слуховое (средний ряд) предъявления цифр в отсутствие вербальной реакции и в случае, когда человек называет цифру вслух (нижний ряд). По оси ординат на профилях реакций - число нейронных популяций, в которых были обнаружены значимые (Р < 0.05) отличия частоты разряда в бине от средней фоновой частоты, на ПСГ - средняя частота разряда в бине; по оси абсцисс - время в бинах (бин - 100 мс). На гистограммах: S —предъявление информационного стимула (цифры); S - предъявление триггерного стимула, служащего сигналом для вербальной реакции испытуемого; R - название стимула. Остальные обозначения - как на рис. 32
Эти данные, однако, должны расцениваться пока как вероятность появления такого рода нейродинамики. Только длительные повторные исследования позволят уточнить, насколько именно вероятна та или другая из описанных ситуаций. Ивто же время уже можно говорить о более или менее селективной реакции на психологические пробы в различных нейронных популяциях.
4
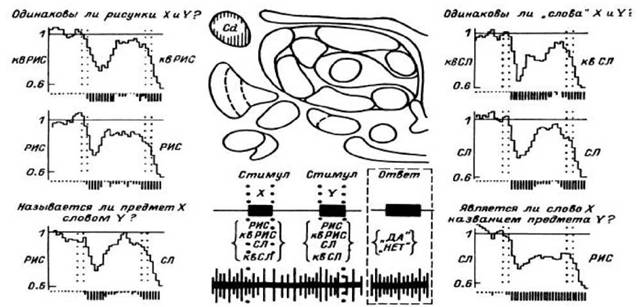
Рис. 34. ПСГ импульсной активности нейронной популяции в хвостатом ядре левого полушария при выполнении серии тестов, связанных со сравнением слов, квазислов, рисунков известных и неизвестных объектов.
|
|
|
Схематическое изображение структуры пробы дано в середине нижней части рисунка. В любой пробе предъявляются два стимула, каждый из которых может быть рисунком известного (РИС) или неизвестного (кв РИС) объекта, словом (СЛ) или квазисловом (кв СЛ). В зависимости от даваемой перед началом теста инструкции испытуемый в каждой пробе отвечает «Да» или «Нет» с учетом особенностей предъявленных стимулов
Большую или меньшую селективность реакций нейронных популяций удалось наблюдать и при предъявлении совокупностей тестов с опознанием слов, сравнением слов и квазислов, слов с соответствующими и не соответствующими им изображениями, сравнением изображений, имеющих и не имеющих смыслового значения, и т. п. Иллюстрацией сказанного может служить рис. 34 (Гоголицын и др., 19876).
Особую осторожность (опять с учетом динамичности нейронных коррелятов мышления) следует проявлять в этих исследованиях при оценке негативных эффектов
- когда нет значимых изменений импульсной активности при психологических пробах. При наличии перестроек на основе одного-двух тестов правильнее говорить, что данная зона, являясь звеном системы, может более или менее постоянно участвовать в обеспечении данного вида деятельности. При отсутствии перестроек для того, чтобы сказать, что данная зона мозга не может участвовать в обеспечении реализуемой деятельности, безусловно, необходимы повторные исследования (особенно это относится к случаям с высокой селективностью реакций). С учетом этих положений, может быть, следует осторожнее относиться к данным, полученным
|
|
|
в результате электрической стимуляции открытого мозга (Ojemann, 1983), на основе которых описывается высокая функциональная селективность в обеспечении мыслительных и преимущественно вербально-мнестических процессов.
Исследования импульсной активности нейронных популяций подтвердили многие механизмы мозга, показанные или предсказанные нами ранее. Эти работы безусловно явились и существенно новой ступенью в изучении различных мозговых механизмов. Это, в частности, относится к динамичности мозговых коррелятов мыслительной деятельности, которая была, во-первых, гораздо полнее изучена, доказана и, во- вторых, рассмотрена более полно в плане внутренних механизмов, определяющих динамичность нейрофизиологических коррелятов мыслительных процессов.
|
|
|
Ранее нами (Бехтерева и др. 1970) было показано, как могут изменяться свойства зоны мозга в зависимости от состояния различных видов биохимической медиации. Позднее В. А. Илюхиной (1977) была обнаружена зависимость свойств мозговых зон от уровня так называемого постоянного потенциала, наиболее стабильной составляющей СМФП - омега-потенциала (по: Илюхина, 19826). При одновременном отведении и исследовании импульсной активности нейронных популяций менее стабильных СМФП была обнаружена зависимость функциональной характеристики зоны мозга и от этого показателя.
В исследованиях С. В. Медведева и сотрудников (Медведев, Белов, 1985, 1986; Медведев и др., 1987) регистрировалась динамика импульсной активности нейронов (ИАН) при одновременном отведении с той же точки мозга, с того же электрода (что определялось возможностями полиэлектронейрографа, - Данько, Каминский, 1982) СМФП в декасекундном диапазоне (тау-волн, по В. А. Илюхиной). Больному предъявлялись психологические пробы на обобщение зрительного стимула.
В зонах мозга, расположенных в области различных ядер таламуса, стриопаллидарной системы была обнаружена зависимость динамики ИАН по ходу реализации психологического теста от фазовых характеристик тау-волн в той же зоне (рис. 35). Особенно интересной была в этом случае тонкая модуляция функциональных свойств мозговых зон.
|
|
|
Как представлено в работах В. А. Илюхиной (1977, 1986), при изменениях наиболее стабильной составляющей СМФП наблюдались перестройки функциональных свойств зон мозга, которые можно было характеризовать типом «да-нет», то есть свойство либо исчезало, либо появлялось. При изменениях фазы тауволн могло наблюдаться уменьшение или увеличение изменений ИАН в различные фазы психологического теста. Так, например, в зоне, расположенной в области заднего вентроорального ядра таламуса, при реализации психологического теста на восходящей и частично нисходящей фазе тау-волны существенные изменения ИАН обнаруживались в фазе принятия решения. В той же зоне при реализации того же теста на нисходящей-восходящей (рис. 35) фазе тау-волны изменения частоты ИАН в
фазу принятия решения отсутствовали.
При предъявлении в тесте двух абсолютно одинаковых сигналов, из которых только первый нес информационную нагрузку, а второй был в контексте теста незначим, на восходящей- нисходящей фазе тау-волны обнаруживалась реакция только на первый из двух сигналов. А при предъявлении их на нисходящей- восходящей фазе селективность реакции зоны мозга исчезала - зона реагировала одинаково на оба сигнала.
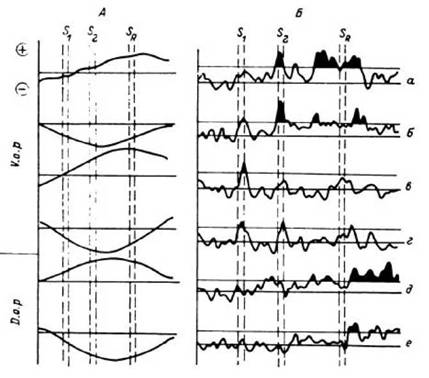
Рис. 35. Зависимость
динамики импульсной активности нейронов от фазовых характеристик тау-волн тех же зон мозга при мыслительной деятельности.
А - результаты усреднения СМФП в декасекундном диапазоне по положительным (б, г, е) и отрицательным (а, в, д) группам знакового ключа; Б - ПСГ текущей частоты импульсной активности нейрона: а, б, д, е - смысловые группы; в, г - «пустышки»; а- г - регистрация в области заднего вентроорального ядра таламуса (V. о. р); д, е - регистрация в области заднего дорсоорального ядра таламуса (D. о, р). Вертикальными штриховыми линиями обозначены моменты предъявления I (S ), II (S ) стимулов и сигнала, разрешающего речевой ответ больному (SR)
Такого рода модулирующее влияние тау-волн обнаружено не во всех исследованных зонах и не во всех зонах с одинаковой выраженностью. Обнаруженная зависимость должна изучаться и, может быть, через некоторое время удастся уточнить, какова степень общности данного модулирующего механизма мозга. В данном случае именно выявление его в каких-то зонах мозга должно расцениваться как импульс к дальнейшему поиску, а не как отсутствие их и, следовательно, как
повод к «закрытию проблемы».
Исследования нейрофизиологических механизмов мыслительной деятельности, несомненно, целесообразно продолжить, чтобы шаг за шагом получать все более полные характеристики вклада различных мозговых зон коры и глубоких структур мозга в обеспечение мыслительной деятельности. Одновременно важно дальнейшее выяснение механизмов мозга, их общего и частного значения. Мы очень ждем от работающих вместе с нами представителей точных наук методов исследования, все более адекватных тому, как мозг взрослого человека решает простые, сложные и сложнейшие задачи. Надо также надеяться, что в самом ближайшем будущем изучение местных и системных процессов в мозгу человека сольется в единое целое. Вместе с психологами физиологи должны попытаться понять мозговые механизмы не только мыслительной деятельности, обеспечивающей ежедневную жизнь человека, но и тех процессов, которые служат прорыву в неизвестное, то есть механизмы творчества. Однако какими бы интригующими ни были результаты будущего, они не явятся поводом для третьего издания данной книги. В задачи книги входило наряду с детализацией ряда данных прежде всего изложение общих подходов к изучению здорового и больного мозга и общих принципов его работы.
Глава шестая
Нейрофизиология памяти человека и общие механизмы его мозга
И все это может сделать одна только физиология, так как она одна держит в своих руках ключ к истинно научному анализу психических явлений.
И. М. Сеченов
Давно и широким фронтом ведется настойчивое многоплановое изучение проблемы памяти. Опыт показал, что достаточно полноценная расшифровка ее механизмов возможна лишь на основе интегративного подхода, но это не исключает отдельных, разных по значимости аналитических этапов ее изучения.
Физиологическое изучение памяти базируется главным образом на исследовании тех изменений различных физиологических показателей в мозгу, которые происходят в ходе запоминания (обучения), динамики следовых явлений и отражения процесса считывания из долгосрочной памяти. Ценные данные о механизмах памяти у человека были получены, как известно, при клинико-анатомических сопоставлениях (Бехтерев, 1900, и многие другие), в процессе развития нейропсихологии (Лурия, 1977; Хомская,
1977) и при анализе результатов электрических воздействий на кору и различные глубокие образования мозга (Ojemann, 1978a; Смирнов, 1976). Физиологический аспект проблемы широко разрабатывается в экспериментальных исследованиях различных лабораторий, результаты которых обобщены в монографиях и сборниках
(Talland, 1965; John, 1967; Механизмы модуляции памяти, 1976; Механизмы управления памятью, 1979; Вартанян, Пирогов, 1987; Механизмы памяти, 1987, и др.). Стройная биоэлектрическая гипотеза памяти предложена М. Н. Ливановым (1977).
Данные о физиологических коррелятах памяти получены не только в эксперименте на животных, ной в исследованиях, проводимых у человека (Гречин, 1975; Раева, Ливанов, 1975; Кропотов, Гречин, 1976; John, 1977; Кропотов, 1979а, и др.).
Интерес к проблеме памяти все возрастает. При этом совершенствование методики исследований, формулирование и разработка новых концепций о механизмах мозга будут способствовать изучению процессов памяти. Широкие запросы педагогики и клиники остро ставят задачу изыскания путей управления процессом памяти, его различными свойствами. Поток обрушивающихся на человека сведений все увеличивается, поэтому возникла необходимость создать способы запоминания иногда очень большого количества сведений одновременно. И. М. Сеченов писал: «Память - сила, которая лежит в основе всего психического развития. Не будь в самом деле этой силы психическое развитие было бы невозможностью» (Сеченов, 1952а, с. 99).
Изыскивая предпосылки для улучшения запоминания, надо одновременно искать и противоядие - способы улучшения выделения значимости информации (эта задача в общем виде рассматривается техническими специалистами как проблема сигнал - шум), а также пути дезактивации, если не полного «стирания» памяти.
Одновременная постановка этих задач важна и для обучения здорового мозга, и для переобучения мозга больного человека путем коррекции матрицы памяти, поддерживающей устойчивое патологическое состояние.
Обнаружение и исследование динамики физиологических изменений в различные фазы памяти оказываются возможными потому, что процесс запоминания в ходе обучения чаще всего происходит не мгновенно, а имеет развертку во времени. В то же время литературные данные и наблюдавшиеся нами при электрической стимуляции мозга явления типа импринтинга (запечатления) свидетельствуют о том, что мгновенное запоминание, запечатление возможно не только на самых ранних этапах онтогенеза (Бехтерева, Смирнов, 1975). Биологически, по-видимому, для организма выгодно иметь разные формы памяти. Реализация основных, важнейших для сохранения вида процессов организма и человека, и животных определяется генетической памятью, не развивающейся, а скорее проявляющейся в онтогенезе. Индивидуально приобретенная память определяет лишь адекватные коррекции протекающих процессов.
Для осуществления функций, важнейших для сохранения индивида как представителя вида, используется механизм мгновенной памяти - запечатление.
И, наконец, все другие виды деятельности, определяющие жизнь индивида в меняющейся среде и особенно жизнь человека в специально человеческой среде, базируются на более или менее быстро, но, как правило, не мгновенно
формирующейся памяти. Скорость ее формирования является функцией значимости события и, таким образом, соответствующей мотивации. Она зависит от эмоционально активных или эмоционально нейтральных активационных механизмов и, следовательно, от функционального фона мозга. Этот процесс связан с генетическими особенностями мозга и целым рядом других, не всегда легко учитываемых факторов.
Биологической выгодой не мгновенного запоминания в процессе обучения является, по-видимому, возможность «отбора на запись», фильтрации информации и, что очень важно, одновременно идущего ее упорядочивания. В случае психонервной памяти примером упорядочивания является соотнесение ее с различными смысловыми и другими полями со всеми вытекающими отсюда последствиями иерархии дальнейшего ассоциативного поиска.
Генетически у человека имеются высокосовершенные предпосылки к психонервной памяти, реализующиеся в процессе обучения, индивидуального и особенно социально обогащенного развития. Так, не только обучение смысловому значению слов, но и использование их как важнейших единиц мыслительной деятельности осуществляется в ходе индивидуального развития человека в условиях общения с особями данного биологического вида. Именно в процессе обучения идет формирование и накопление базиса долговременной психонервной памяти при одновременной тренировке ее последующего использования в форме считывания, перевода из долгосрочной памяти в оперативную.
Как указывалось выше, связь мыслительных операций с процессами памяти неразрывна. Выделение в исследованиях собственно мыслительных процессов или собственно процесса памяти всегда имеет искусственный характер и определяет скорее расстановку акцентов, чем их сущность. Действительно, уже простейшие умозаключения немыслимы без активации и использования соответствующего базиса долгосрочной памяти вербального и невербального типов. В то же самое время по ходу тех же и всех других процессов все время происходит не только пополнение базиса памяти, но и трансформация ассоциаций и их иерархии. В полном соответствии со сказанным данные, полученные в процессе изучения тонких нейрофизиологических коррелятов слов и мыслительных процессов (Бехтерева и др., 1977а), оказались существенными для понимания целого ряда закономерностей нейропсихологии, нейролингвистики, физиологии анализаторных систем, искусственного интеллекта и ряда других направлений научных исследований. Они вместе с данными электрических воздействий на мозг оказались важнейшими для комплексно решаемой научной проблемы - проблемы памяти.
В предыдущей главе рассматривались результаты исследования перестроек импульсной активности, соотносимых с восприятием, удержанием в памяти и воспроизведением слов и некоторыми мыслительными операциями. Было показано, что в процессе восприятия слов в импульсной активности нейронных популяций
формируется паттерн, характеризующийся изменениями частоты и структуры импульсного потока, формы разрядов нейронов и их групп внутри нейронных популяций и между дистантно расположенными популяциями. Уже в этом паттерне при использовании в психологических тестах слов, имеющих смысловую общность, обнаруживались нейрофизиологические корреляты этой общности в виде характерных перестроек взаимодействия нейронных группировок. Далее были выявлены перестройки структуры импульсного потока в виде очень динамичных определенных последовательностей интервалов и мало динамичных, но менее определенных изменений интервалов между импульсами. Общие перестройки структуры импульсных потоков, появляющиеся в разных зонах мозга (см. выше), рассматривались нами как фактор, способствующий организации мозговой системы. Общие последовательности интервалов, обнаруженные в коррелятах слов, соотносимых по смыслу, отражали переведенный из латентной формы в активную системообразующий фактор долгосрочной памяти. Они явились биоэлектрическим выражением активации ассоциативных связей, образовавшихся в результате индивидуального опыта и хранимых в долгосрочной памяти.
При инструкции запомнить слово и произнести его по команде и относительно небольших изменениях интервала между заданием и ответом можно видеть настраивание мозга на тот интервал в форме появления вначале слабо выраженных, а затем вполне отчетливых перестроек импульсной активности в виде появления паттерна сигнала, являющегося «оригинальной копией» первого развернутого паттерна, названного управляющим (Бехтерева и др., 19756). Термин «оригинальная копия» заимствован из искусства. Если он и не очень удачен, то отражает и сходство управляющего паттерна с первым, возникшим при восприятии слова, и отличие от него. Отличие формально характеризуется его соотносимостью с акустическими (а вероятно, и моторными) характеристиками ответа больного. По существу, различие определяется тем, что второй паттерн в тестах с предъявлениями известных слов является результатом активации долгосрочной памяти лица, выполняющего тест. Подчеркивание в этих случаях значения активации долгосрочной памяти основано на сопоставлении нейрофизиологической динамики в тестах на краткосрочную память с динамикой при выполнении мыслительных операций. Если в первом случае соотношения относительно просты: полный первичный паттерн-компрессированный паттерн (паттерны)-полный управляющий паттерн, соотносимые с одним и тем же словом, то при различных входном и выходном сигналах, где входной является лишь одной из детерминант выходного - ответа, видно, как мозг формирует ответ при наличии соответствующего базиса долгосрочной памяти. Влияние последнего на динамику паттерна-кода в импульсной активности нейронных популяций прослеживается при варьировании наиболее простых тестов - тестов на краткосрочную память. При этом обнаруживается, что выполнение тестов, формально составленных одинаково, но с использованием слов, встречающихся в речи с разной частотой, известных и неизвестных, может осуществляться на различной нейрофизиологической основе.
Действительно, и при предъявлении редко и часто встречающихся в речи известных слов, и при предъявлении неизвестных слов по команде «Повторите» корректно или ошибочно воспроизводится то, что было в задании. Однако после восприятия известных слов наблюдается указанная выше нейрофизиологическая динамика, а при удержании в памяти неизвестного слова возникший при его восприятии паттерн сохраняется в импульсной активности, хотя и в дискретной форме, до речевого ответа. Сохранение паттерна связано в данных конкретных тестах и с тем, что больному предлагалось запомнить предъявленные сигналы. Однако идентичность этого условия при различии характера предъявляемых сигналов и соответственно этому различие мозговой нейродинамики позволили нам предположить, что второй вариант (сохранение паттерна) связан с отсутствием тормозящих влияний базиса долгосрочной памяти при неопознании субъективно нового сигнала. Нейрофизиологическим механизмом стабильности паттерна могут быть явления, протекающие по типу не до конца еще прослеженного и изученного процесса так называемой реверберации (Lorente de No, 1938; Беритов, 1960). Отношение циркуляции возбуждения по замкнутым нейронным цепочкам, спустя некоторое время после окончания действия какого-либо стимула к процессу краткосрочной памяти, недавно получило прямое экспериментальное доказательство на животных при исследовании взаимодействий функционально связанных нейронов (Вартанян и др., 1986). Стабильность, сохранение паттерна создают предпосылки к оптимизации в этих условиях формирования соответствующего биохимического базиса долгосрочной памяти. Это объяснение почти напрашивается, и мы пока не можем предложить лучшего, хотя, естественно, можно было бы предположить, что дело не в отсутствии тормозящих влияний, а в самом факторе новизны, активации соответствующих детекторов. Но что такое, если вдуматься в физиологическую сторону вопроса, новизна?
Дата добавления: 2018-08-06; просмотров: 172; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!