Возрастные особенности спинного мозга
УЧЕНИЕ О НЕРВНОЙ СИСТЕМЕ — НЕВРОЛОГИЯ (NEUROLOGIA)
 Функцией нервной системы является управление деятельностью различных систем и аппаратов, составляющих целостный организм, координирование протекающих в нем процессов, установление взаимосвязей организма с внешней средой. Великий русский физиолог И. П. Павлов писал: «Деятельность нервной системы направляется, с одной стороны, на объединение, интеграцию работы всех частей организма, с другой — на связь организма с окружающей средой, на уравновешивание системы организма с внешними условиями».
Функцией нервной системы является управление деятельностью различных систем и аппаратов, составляющих целостный организм, координирование протекающих в нем процессов, установление взаимосвязей организма с внешней средой. Великий русский физиолог И. П. Павлов писал: «Деятельность нервной системы направляется, с одной стороны, на объединение, интеграцию работы всех частей организма, с другой — на связь организма с окружающей средой, на уравновешивание системы организма с внешними условиями».
Нервы проникают во все органы и ткани, образуют многочисленные разветвления, имеющие рецепторные (чувствительные) и эффекторные (двигательные, секреторные) окончания, и вместе с центральными отделами (головной и спинной мозг) обеспечивают объединение всех частей организма в единое целое. Нервная система регулирует функции движения, пищеварения, дыхания, выделения, кровообращения, лимфоотток, иммунные (защитные) и метаболические процессы (обмен веществ) и др.
Деятельность нервной системы, по словам И. М. Сеченова, носит рефлекторный характер. Рефлекс (лат. reflexus — отраженный) — это ответная реакция организма на то или иное раздражение (внешнее или внутреннее воздействие), которая происходит при участии центральной нервной системы (ЦНС). Человеческий организм, обитающий в окружающей его внешней среде, взаимодействует с ней. Среда влияет на организм, и организм в свою очередь соответствующим образом реагирует на эти влияния. Протекающие в самом организме процессы также вызывают ответную реакцию. Таким образом, нервная система обеспечивает взаимосвязь и единство организма и среды.
Структурно-функциональной единицей нервной системы является нейрон (нервная клетка, нейроцит). Нейрон состоит из тела и отростков. Отростки, проводящие к телу нервной клетки нервный импульс, получили название дендритов. От тела нейрона нервный импульс направляется к другой нервной клетке или к рабочей ткани по отростку, который называют аксоном, или нейритом. Нервная клетка динамически поляризована, т. е. способна пропускать нервный импульс только в одном направлении— от дендрита через тело клетки к аксону (нейриту).
Нейроны в нервной системе, вступая в контакт друг с другом, образуют цепи, по которым передаются (движутся) нерв-
ные импульсы. Передача нервного импульса от одного нейрона к другому происходит в местах их контактов и обеспечивается особого рода образованиями, получившими название межнейронных синапсов. Различают синапсы аксосоматические, когда окончания аксона одного нейрона образуют контакты с телом следующего, и аксодендритические, когда аксон вступает в контакт с дендритами другого нейрона. Контактный тип отношений в синапсе при различных физиологических состояниях может, очевидно, либо «создаваться», либо «разрушаться», обеспечивая возможность избирательной реакции на любое раздражение. Помимо этого, контактное построение цепочек нейронов создает возможность для проведения нервного импульса в определенном направлении. Благодаря наличию контактов в одних синапсах и разъединению в других проведение импульса может нарушаться.
В нервной цепочке различным нейронам присущи разные функции. В связи с этим выделяют три основных типа нейронов по их морфофункциональной характеристике.
1. Чувствительные, рецепторные, или афферентные, нейроны.
Тела этих нервных клеток лежат всегда вне головного или спин
ного мозга, в узлах (ганглиях) периферической нервной систе
мы. Один из отростков, отходящих от тела нервной клетки,
следует на периферию к тому или иному органу и заканчивается
там тем или иным чувствительным окончанием — рецептором,
который способен трансформировать энергию внешнего воздей
ствия (раздражения) в нервный импульс. Второй отросток на
правляется в ЦНС, спинной мозг или в стволовую часть голов
ного мозга в составе задних корешков спинномозговых нервов
или соответствующих черепных нервов.
Различают следующие виды рецепторов в зависимости от локализации:
1) экстероцепторы воспринимают раздражение из
внешней среды. Они расположены в наружных покровах тела,
в коже и слизистых оболочках, в органах чувств;
2) интероцепторы получают раздражение главным
образом при изменениях химического состава внутренней среды
организма и давления в тканях и органах;
3) проприоцепторы воспринимают раздражения в мыш
цах, сухожилиях, связках, фасциях, суставных капсулах.
Рецепцию, т. е. восприятие раздражения и начавшееся распространение нервного импульса по нервным проводникам к центрам, И. П. Павлов относил к началу процесса анализа.
2. Замыкательный, вставочный, ассоциативный, или кондук
торный, нейрон.Этот нейрон осуществляет передачу возбужде
ния с афферентного (чувствительного) нейрона на эфферентные.
Суть этого процесса заключается в передаче полученного аф
ферентным нейроном сигнала эфферентному нейрону для испол
нения в виде ответной реакции. И. П. Павлов определил это
действие как «явление нервного замыкания». Замыкательные
(вставочные) нейроны лежат в пределах ЦНС.
268
269
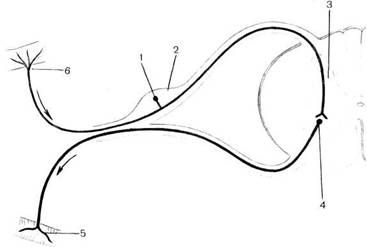
Рис. 108.Схема простейшей рефлекторной дуги.
1 — афферентный (чувствительный) нейрон; 2 - спинномозговой узел; 3 — серое вещество спинного мозга; 4 — эфферентный (двигательный) нейрон; 5 — двигательное нервное окончание в мышцах; 6 — чувствительное нервное окончание в коже.
3. Эффекторный, эфферентный (двигательный, или секреторный) нейрон.Тела этих нейронов находятся в ЦНС (или на периферии—в симпатических, парасимпатических узлах). Аксоны (нейриты) этих клеток продолжаются в виде нервных волокон к рабочим органам (произвольным — скелетным и непроизвольным— гладким мышцам, железам).
После этих общих замечаний рассмотрим более детально рефлекторную дугу и рефлекторный акт как основной принцип деятельности нервной системы. Рефлекторная дугапредставляет собой цепь нервных клеток, включающую афферентный (чувствительный) и эффекторный (двигательный, или секреторный) нейроны, по которым нервный импульс движется от места своего возникновения (от рецептора) к рабочему органу (эффектору). Большинство рефлексов осуществляется при участии рефлекторных дуг, которые образованы нейронами низших отделов ЦНС — нейронами спинного мозга.
Простейшая рефлекторная дуга (рис. 108) состоит только из двух нейронов — афферентного и эффекторного (эфферентного). Тело первого нейрона (рецепторного, афферентного), как отмечено, находится вне ЦНС. Обычно это псевдоуниполярный (униполярный) нейрон, тело которого расположено в спинномозговом узле (ganglion spindle) или чувствительном узле черепных нервов (ganglion sensoridle nn. cranidlii). Периферический отросток этой клетки следует в составе спинномоз-
говых нервов или имеющих чувствительные волокна черепных нервов и их ветвей и заканчивается рецептором, воспринимающим внешнее (из внешней среды) или внутреннее (в органах, тканях) раздражение. Это раздражение трансформируется рецептором в нервный импульс, который достигает тела нервной клетки, а затем по центральному отростку (совокупность таких отростков образует задние, или чувствительные, корешки спинномозговых нервов) направляется в спинной мозг или по соответствующим черепным нервам — в головной мозг. В сером веществе спинного мозга или в двигательном ядре головного мозга этот отросток чувствительной клетки образует синапс с телом второго нейрона (эфферентного, эффекторного). В межнейронном синапсе с помощью медиаторов происходит передача нервного возбуждения с чувствительного (афферентного) нейрона на двигательный (эфферентный) нейрон, отросток которого выходит из спинного мозга в составе передних корешков спинномозговых нервов или двигательных (секреторных) нервных волокон черепных нервов и направляется к рабочему органу, вызывая сокращение мышцы, либо торможение, либо усиление секреции железы.
Как правило, рефлекторная дуга состоит не из двух нейронов, ;i устроена гораздо сложнее. Между двумя нейронами — рецеп-торным (афферентным) и эффекторным (афферентным)—имеется один или несколько замыкательных (вставочных) нейронов. В этом случае возбуждение от рецепторного нейрона по его центральному отростку передается не прямо эффекторной нервной клетке, а одному или нескольким вставочным нейронам. Роль вставочных нейронов в спинном мозге выполняют клетки, лежащие в сером веществе задних столбов. Часть этих клеток имеет аксон (нейрит), который направляется к двигательным клеткам передних рогов спинного мозга того же уровня и замыкает рефлекторную дугу на уровне данного сегмента спинного мозга. Аксон других клеток может в спинном мозге предварительно Т-образно делиться на нисходящую и восходящую ветви, которые направляются к двигательным нервным клеткам передних рогов соседних, выше- или нижележащих сегментов. На пути следования каждая из отмеченных восходящих или нисходящих ветвей может отдавать коллатерали к двигательным клеткам этих и других соседних сегментов. В связи с этим становится понятным, что раздражение даже самого минимального числа рецепторов может передаваться не только нервным клеткам какого-то определенного сегмента спинного мозга, но и распространяться на клетки нескольких соседних сегментов. В результате ответная реакция представляет собой сокращение не одной мышцы и даже не одной группы мышц, а сразу нескольких групп. Так, в ответ на раздражение возникает сложное рефлекторное движение. Это и есть одна из ответных реакций организма (рефлекс) в ответ на внешнее или внутреннее раздражение.
270
271
И. М. Сеченов в своем труде «Рефлексы головного мозга» выдвинул идею причинности (детерминизма), отмечая, что каждое явление в организме имеет свою причину, и рефлекторный эффект является ответом на эту причину. Эти идеи получили дальнейшее творческое развитие в трудах С. П. Боткина и И. П. Павлова, которые являются основоположниками учения о нервизме. Огромная заслуга И. П. Павлова состоит в том, что он распространил учение о рефлексе на всю нервную систему, начиная от низших отделов и кончая самыми высшими ее отделами, и экспериментально доказал рефлекторную природу всех без исключения форм жизнедеятельности организма. По мнению И. П. Павлова, простая форма деятельности нервной системы, которая является постоянной, прирожденной, видовой и для формирования структурных предпосылок которой не требуется социальных условий, должна обозначаться как безусловный рефлекс.
Кроме этого, существуют приобретаемые в течение индивидуальной жизни временные связи с окружающей средой. Возможность приобретения временных связей позволяет организму устанавливать многообразнейшие и сложнейшие отношения с внешней средой. Эту форму рефлекторной деятельности И. П. Павлов назвал условнорефлекторной (в отличие от без-условнорефлекторной). Местом замыкания условных рефлексов является кора большого мозга. Головной мозг и его кора -основа высшей нервной деятельности.
П. К. Анохин и его школа экспериментально подтвердили наличие так называемой обратной связи рабочего органа с нервными центрами — «обратную афферентацию». В тот момент, когда из центров нервной системы эфферентные импульсы достигают исполнительных органов, в них вырабатывается ответная реакция (движение или секреция). Этот рабочий эффект раздражает рецепторы самого исполнительного органа. Возникшие в результате этих процессов импульсы по афферентным путям направляются обратно в центры спинного или головного мозга в виде информации о выполнении органом определенного действия в каждый данный момент. Таким образом создается возможность точного учета правильности исполнения команд в виде нервных импульсов, поступающих к рабочим органам из нервных центров, и постоянной их коррекции. Существование двусторонней сигнализации по замкнутым, круговым или кольцевым рефлекторным нервным цепочкам «обратной афферентации» позволяет производить постоянные, непрерывные, ежемоментные коррекции любых реакций организма на любые изменения условий внутренней и внешней среды. Без механизмов обратной связи немыслимо приспособление живых организмов к окружающей среде. Так, на смену старым представлениям о том, что в основе деятельности нервной системы лежит «разомкнутая» (незамкнутая) рефлекторная дуга, пришло представление о замкнутой, кольцевой, цепи рефлексов.
Нервную систему человека условно подразделяют по топографическому принципу на центральную и периферическую.
К центральной нервной системе (ЦНС)относят спинной мозг и головной мозг, которые состоят из серого и белого вещества. Серое вещество спинного и головного мозга — это скопления нервных клеток вместе с ближайшими разветвлениями их отростков. Белое вещество — это нервные волокна, отростки нервных клеток, имеющие миелиновую оболочку (отсюда белый цвет волокон). Нервные волокна образуют проводящие пути спинного и головного мозга и связывают различные отделы ЦНС и различные ядра (нервные центры) между собой.
Периферическую нервную системусоставляют корешки, спинномозговые и черепные нервы, их ветви, сплетения и узлы, лежащие в различных отделах тела человека.
По другой, анатомо-функциональной, классификации единую нервную систему также условно подразделяют на две части: соматическую и автономную, или вегетативную. Соматическая нервная системаобеспечивает иннервацию главным образом тела— сомы, а именно кожи, скелетных (произвольных) мышц. Этот отдел нервной системы выполняет функции связи организма с внешней средой при помощи кожной чувствительности и органов чувств.
Автономная (вегетативная) нервная системаиннервирует все внутренности, железы, в том числе и эндокринные, непроизвольную мускулатуру органов, кожи, сосудов, сердца, а также регулирует обменные процессы во всех органах и тканях.
Автономная нервная система в свою очередь подразделяется на парасимпатическую часть, pars parasympdthica, и симпатическую часть, pars sytnpathica. В каждой из этих частей, как и в соматической нервной системе, выделяют центральный и периферический отделы.
Такое деление нервной системы, несмотря на его условность, сложилось традиционно и представляется достаточно удобным для изучения нервной системы в целом и ее отдельных частей. В связи с этим в дальнейшем мы также будем в изложении материала придерживаться этой классификации.
Нервная система в филогенезе
Любой живой организм, находящийся в определенной среде обитания, постоянно взаимодействует с ней. Из внешней среды живой организм получает необходимые для жизнедеятельности продукты питания, во внешнюю среду происходит выделение ненужных для организма веществ. Внешняя среда оказывает на организм благоприятное или неблагоприятное воздействие. На эти воздействия и изменения внешней среды живой организм реагирует путем изменения своего внутреннего состояния. Реакция живого организма может проявляться в виде роста, усиле-
272
273
ния или ослабления обменных процессов, движений или секреции.
Простейшие одноклеточные организмы не имеют нервной системы, и все отмеченные реакции у них являются проявлением деятельности одной клетки.
Воздействия внешней среды многоклеточные организмы воспринимают наружными эктодермальными клетками. Такие клетки специализируются на восприятии раздражения, трансформации его в биоэлектрические потенциалы и проведении возбуждения. Из эктодермальных клеток, погружающихся в глубь тела, в последующем возникает примитивно устроенная нервная система многоклеточных организмов.
У многоклеточных организмов нервная система состоит из клеток, которые связаны друг с другом отростками, способными воспринимать раздражение от любых участков поверхности тела и посылать импульсы другим клеткам, регулируя их деятельность. Такая наиболее просто образованная сетевидная, или диффузная, нервная система имеется у кишечнополостных, например, у гидры. У таких животных различают два вида клеток. Одни из них — рецепторные клетки — расположены между клетками эктодермы. Другие — эффекторные клетки — находятся в глубине организма, связаны друг с другом и с клетками, обеспечивающими ответную реакцию. Раздражение любого участка поверхности тела гидры приводит к возбуждению глубжележа-щих клеток, в результате чего живой многоклеточный организм проявляет двигательную активность, захватывает пищу или уходит от противника.
Дальнейшая эволюция нервной системы характеризуется концентрацией нервных клеток, которые формируют нервные центры, или нервные узлы (ганглии), с отходящими от них нервными стволами. На этом этапе возникает узловая форма нервной системы. У представителей сегментированных животных (например, у кольчатых червей) узлы расположены вентральнее пищеварительной трубки и соединяются поперечными и продольными нервными стволами. От этих узлов отходят нервы, разветвления которых заканчиваются также в пределах данного сегмента. Посегментно расположенные ганглии служат рефлекторными центрами соответствующих сегментов тела у этих животных. Продольные нервные стволы соединяют друг с другом узлы разных сегментов на одной половине тела и образуют две продольные брюшные цепочки. В головном конце тела, дорсальнее глотки, расположена одна пара более крупных надглоточных узлов, которая окологлоточным кольцом нервов соединяется с первой парой узлов брюшной цепочки. Эти узлы развиты более других и являются прообразом головного мозга позвоночных животных. Такое сегментарное строение нервной системы позволяет при раздражении определенных участков поверхности тела животного не вовлекать в ответную реакцию все нервные клетки тела, а использовать только клетки данного сегмента.
Следующий этап развития нервной системы состоит в том, что нервные клетки расположены уже не в виде отдельных узлов, а формируют продолговатый непрерывный нервный тяж, внутри которого имеется полость. На этой стадии нервная система называется трубчатой нервной системой. Строение нервной системы в виде нервной трубки характерно для всех представителей хордовых — от наиболее просто устроенных бесчерепных до млекопитающих животных и человека.
В соответствии с метамерностью тела хордовых животных единая трубчатая нервная система состоит из ряда однотипных повторяющихся структур, или сегментов. Отростки нейронов, входящих в состав данного нервного сегмента, разветвляются, как правило, в определенном, соответствующем данному сегменту участке тела и его мускулатуре.
Таким образом, совершенствование форм движения животных (от перистальтического способа у простейших многоклеточных до передвижения с помощью конечностей) привело к необходимости совершенствования строения нервной системы. У хордовых туловищный отдел нервной трубки — это спинной мозг. В спинном мозге и в стволовой части формирующегося головного мозга у хордовых в вентральных отделах нервной трубки располагаются «двигательные» клетки, аксоны которых формируют передние («двигательные») корешки, а в дорсальных-нервные клетки, с которыми вступают в связь аксоны «чувствительных» клеток, расположенных в спинномозговых узлах.
В головном конце нервной трубки в связи с развивающимися в передних отделах туловища органами чувств и наличием здесь жаберного аппарата сегментарное строение нервной трубки хотя и сохраняется, однако претерпевает значительные изменения. Эти отделы нервной трубки являются зачатком, из которого развивается головной мозг. Утолщение передних отделов нервной трубки и расширение ее полости — это начальные этапы дифференциации головного мозга. Такие процессы наблюдаются уже у круглоротых. На ранних стадиях эмбриогенеза почти у всех черепных животных головной конец нервной трубки состоит из трех первичных нервных пузырьков: ромбовидного, rhom-bencephalon, расположенного ближе всех к спинному мозгу, среднего, mesencephalon, и переднего, prosencephalon.
Развитие головного мозга происходит параллельно с усовершенствованием спинного мозга, причем появление новых центров в головном мозге ставит как бы в подчиненное положение уже существующие центры спинного мозга. В тех участках головного мозга, которые относятся к заднему мозговому пузырю (ромбовидному мозгу), происходит развитие ядер жаберных нервов (X пара — п. vagus), возникают центры, регулирующие процессы дыхания, пищеварения, кровообращения. Несомненное влияние на развитие заднего мозга оказывают появляющиеся уже у низших рыб рецепторы статики и акустики (VIII пара). В связи с этим на данном этапе развития головного мозга преобла-
274
275
дающим над другими отделами является задний мозг (мозжечок и мост мозга). Появление и совершенствование рецепторов зрения и слуха обусловливают развитие среднего мозга, где закладываются центры, отвечающие за зрительную и слуховую функции. Все эти процессы происходят в связи с приспособляемостью организма животных к водной среде обитания.
При переходе животных к новой среде обитания — из водной в воздушную — происходит дальнейшая перестройка как организма в целом, так и его нервной системы. Развитие обонятельного анализатора вызывает дал-ьнейшую перестройку переднего конца нервной трубки (переднего мозгового пузыря), где закладываются центры, регулирующие функцию обоняния, появляется так называемый обонятельный мозг, rhinencep-halon.
Из трех первичных пузырей за счет дальнейшей дифферен-цировки переднего и ромбовидного мозга выделяются следующие 5 отделов (мозговые пузыри): конечный мозг, telen-cephalon; промежуточный мозг, diencephalon; средний мозг, mesencephalon; задний мозг, metencephalon, и продолговатый мозг, medulla oblongdta (bulbus). Центральный канал спинного мозга в головном конце нервной трубки превращается в систему сообщающихся друг с другом полостей мозговых пузырей, получивших название желудочков головного мозга. Дальнейшее развитие нервной системы связано с прогрессивным развитием переднего мозга и возникновением новых нервных центров, которые на каждом последующем этапе занимают положение, все более близкое по отношению к головному концу, и подчиняют своему влиянию ранее существовавшие центры.
Более старые нервные центры, сформировавшиеся на ранних этапах эволюции, не исчезают, а сохраняются, занимая подчиненное положение по отношению к более новым. Так, наряду с впервые возникшими в заднем мозге центрами слуха (ядрами) на более поздних этапах центры слуха появляются в среднем, а затем и в конечном мозгу. У амфибий в переднем мозге уже формируется зачаток будущих полушарий, однако, как и у рептилий, почти все их отделы относятся к обонятельному мозгу. В переднем (конечном) мозге у амфибий, рептилий и птиц различают подкорковые центры (ядра полосатого тела) и кору, которая имеет примитивное строение. Последующее развитие головного мозга связано с возникновением новых рецепторных и эффекторных центров в коре, которые подчиняют себе нервные центры низшего порядка (в стволовой части головного мозга и спинном мозге), координируют их деятельность, объединяя нервную систему в структурное и функциональное целое. Этот процесс получил название кортиколизации функций. Усиленное развитие конечного мозга у высших позвоночных животных (млекопитающих) приводит к тому, что этот отдел преобладает над всеми остальными и покрывает все отделы в виде плаща
(pallium) или коры большого мозга (cortex cerebralis). На смену
древней коре, paleocortex, а затем старой, archeocortex,
занимающей у рептилий дорсальную и дорсолатеральную по
верхности полушарий, приходит новая кора, neocortex. Ста
рые отделы оттесняются на нижнюю (вентральную) поверхность
полушарий и в глубину, как бы свертываются, превращаются
в гиппокамп (аммонов рог) и в прилежащие к нему отделы
мозга. . .
Одновременно с этими процессами происходят дифференци-ровка и усложнение всех других отделов мозга: промежуточного, среднего и заднего, перестройка как восходящих (чувствительных, рецепторных), так и нисходящих (двигательных, эффектор-ных) путей. Так, у высших млекопитающих по сравнению с низшими нарастает масса волокон пирамидных путей, связывающих центры коры большого мозга с двигательными клетками -передних рогов спинного мозга и двигательными ядрами стволовых отделов головного мозга. Увеличение массы волокон проводящих путей (в филогенезе) непосредственно связано с увеличением объема плаща и формированием коры большого мозга.
Наибольшего развития кора большого мозга достигает у человека, что объясняется трудовой деятельностью человека и возникновением речи как средства общения между людьми. И. П. Павлов, создавший учение о второй сигнальной системе, материальным субстратом последней считал сложн.0 устроенную кору полушарий большого мозга — новую кору. Эволюция мозжечка, ял спинного мозга тесно связана с изменением способа перемещения животного в пространстве. Так, у пресмыкающихся, не имеющих конечностей и перемещающихся с помощью движений туловища, спинной мозг не имеет утолщений и состоит из примерно одинаковой величины сегментов. У животных, передвигающихся с помощью конечностей, в спинном мозге появляются утолщения, степень развития которых зависит от функциональной значимости конечностей. Если сильнее развиты передние конечности, например у птиц, то более выражено шейное утолщение спинного мозга. В мозжечке у птиц имеются боковые выпячивания — клочок, flocculus, самая древняя часть полушарий мозжечка. Формируются полушария мозжечка, высокой степени развития достигает червь мозжечка, vermis сеге-ЬеШ.
Если преобладающими являются функции задних конечностей, например у кенгуру, то значительнее выражено поясничное утолщение. У человека диаметр шейного утолщения спинного мозга больше, чем поясничного. Это объясняется тем, что рука, являющаяся органом труда, способна производить более сложные и разнообразные движения, чем нижняя конечность.
В процессе эволюции в связи с развитием высших центров управления деятельностью всего организма в головном мозге спинной мозг попадает в подчиненное положение. В нем сохраняется более старый сегментарный аппарат собственных связей
276
277
спинного мозга и развивается надсегментарный аппарат двусторонних связей с головным мозгом. Эволюция головного мозга проявилась в развитии и совершенствовании рецепторного аппарата, усовершенствовании механизмов приспособления организма к окружающей среде путем изменения обмена веществ, кортиколизации функций. У человека вследствие прямохожде-ния и в связи с усовершенствованием движений верхних конечностей в процессе трудовой деятельности полушария мозжечка развиты гораздо сильнее, чем у животных.
Кора большого мозга является совокупностью корковых концов всех видов анализаторов и представляет собой материальный субстрат конкретно-наглядного мышления (по И. П. Павлову—первая сигнальная система действительности). Дальнейшее развитие мозга у человека определяется его сознательным использованием орудий труда, что позволило человеку не только приспосабливаться к меняющимся условиям среды, как это делают животные, но и самому влиять на внешнюю среду. В процессе общественного труда возникла речь как необходимое средство общения между людьми. Ф. Энгельс в своем труде «Диалектика природы» писал: «Сначала труд, а затем и вместе с ним членораздельная речь явились двумя самыми главными стимулами, под влиянием которых мозг обезьяны постепенно превратился в человеческий мозг, который, при всем своем сходстве с обезьяньим, далеко превосходит его по величине и совершенству»'.
Так у человека появилась способность к абстрактному мышлению и сформировалась система восприятия слова, или сигнала,—вторая сигнальная система, по И. П. Павлову, материальным субстратом которой является новая кора большого мозга.
Развитие нервной системы человека
Нервная система человека развивается из наружного зародышевого листка — эктодермы. В дорсальных отделах туловища зародыша дифференцирующиеся эктодермальные клетки образуют медуллярную (нервную) пластинку (рис. 109). Последняя вначале состоит из одного слоя клеток, которые в дальнейшем дифференцируются на спонгиобласты (из них развивается опорная ткань — нейроглия) и нейробласты (из них развиваются нервные клетки). В связи с тем что интенсивность размножения клеток в различных участках медуллярной пластинки неодинакова, последняя прогибается и постепенно приобретает вид бороздки или желобка. Рост боковых отделов этой нервной (медуллярной) бороздки приводит к тому, что ее края вначале сближаются, а затем срастаются. Таким образом нервная бороздка, замыкаясь в своих дорсальных отделах, превращается в нервную трубку. Сращение первоначально происходит в пе-
 1 Маркс К , Энгельс Ф. Соч. 2-е изд., т. 20. с. 490.
1 Маркс К , Энгельс Ф. Соч. 2-е изд., т. 20. с. 490.
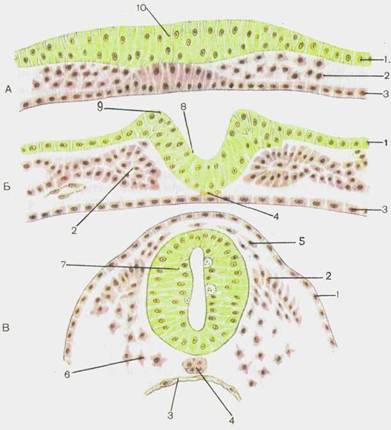
Рис. 109.Ранние стадии развития нервной системы человека. Формирование нервной трубки.
А — нервная пластинка. Б - нервный желобок. В — нервная трубка. 1 -эктодерма; 2 — мезодерма; 3 -энтодерма; 4 - хорда; 5 — ганглиозная пластинка; 6 —мезенхима; 7 — нервная трубка; 8 — нервный желобок; 9 — нерв ный валик; 10 — нервная пластинка.
реднем отделе, несколько отступя от переднего края нервной трубки. Затем срастаются задние, каудальные, ее отделы. На переднем и заднем концах нервной трубки остаются небольшие несращенные участки — нейропоры. После сращения дорсальных отделов нервная трубка отшнуровывается от эктодермы и погружается в мезодерму.
В период образования нервная трубка состоит из трех слоев. Из внутреннего слоя в дальнейшем развивается эпенди-мальная выстилка полостей желудочков мозга и центрального канала спинного мозга, из с р е д н е г о («плащевого») слоя-серое вещество мозга. Наружный слой, почти лишенный
278
279
клеток, превращается в белое вещество. Вначале все стенки нервной трубки имеют одинаковую толщину. Впоследствии более интенсивно развиваются боковые отделы трубки, которые все более утолщаются. Вентральная и дорсальная стенки отстают в росте и постепенно погружаются между интенсивно развивающимися боковыми отделами. В результате такого погружения образуются вентральная и дорсальная продольные срединные борозды будущего спинного и продолговатого мозга.
Со стороны полости трубки на внутренней поверхности каждой из боковых стенок формируются неглубокие продольные пограничные бороздки, которые подразделяют боковые отделы трубки на вентральную основную и дорсальную крыльную пластинки.
Основная пластинка служит зачатком, из которого формируются передние столбы серого вещества и прилежащее к ним белое вещество. Отростки развивающихся в передних столбах нейронов выходяд1 (прорастают) из спинного мозга, образуют передний (двигательный) корешок..Из крыльной пластинки развиваются задние столбы серого вещества и примыкающее к ним белое вещество. Еще на стадии нервной бороздки в латеральных отделах ее выделяются клеточные тяжи, получившие название медуллярных гребешков. В период образования нервной трубки два гребешка, срастаясь, образуют ганглиозную пластинку, располагающуюся дорсальнее нервной трубки, между последней и эктодермой. Впоследствии ганглиозная пластинка вторично делится на два симметричных ганглиозных валика, каждый из которых смещается на боковую поверхность нервной трубки. Затем ганглиозные валики превращаются в соответствующие каждому сегменту туловища спинномозговые узлы, ganglia spindlia, и чувствительные узлы черепных нервов, ganglia sensoridlia nn. cranialium. Клетки, выселившиеся из ганглиозных валиков, служат зачатками и для развития периферических отделов вегетативной нервной системы.
Вслед за обособлением ганглиозной пластинки нервная трубка в головном конце заметно утолщается. Эта расширенная часть служит зачатком головного мозга. Остальные отделы нервной трубки в дальнейшем превращаются в спинной мозг. Нейро-бласты, расположенные в формирующемся спинномозговом узле, имеют форму биполярных клеток. В процессе дальнейшей диф-ференцировки нейробластов расположенные в непосредственной близости к телу клетки участки двух ее отростков сливаются в один Т-образно делящийся затем отросток. Так, клетки спинномозговых узлов становятся по своей форме псевдоуниполярными. Центральные отростки этих клеток направляются в спинной мозг и образуют задний (чувствительный) корешок. Другие отростки псевдоуниполярных клеток растут от узлов к периферии, где имеют рецепторы различных типов.
На ранних стадиях развития эмбриона нервная трубка простирается по всей длине тела. В связи с редукцией каудальных
отделов нервной трубки нижний конец будущего спинного мозга постепенно суживается, образуя терминальную (концевую) нить, filum terminate. Примерно в течение 3 мес внутриутробного развития длина спинного мозга равна длине позвоночного канала. В дальнейшем рост позвоночного столба происходит более интенсивно. В связи с фиксацией головного мозга в полости черепа наиболее заметное отставание в росте нервной трубки наблюдается в ее каудальных отделах. Несоответствие в росте позвоночного столба и спинного мозга приводит как бы к «восхождению» нижнего конца последнего. Так, у новорожденного нижний конец спинного мозга расположен на уровне III поясничного позвонка, а у взрослого — на уровне I—II поясничных позвонков. Корешки спинного мозга и спинномозговые узлы формируются достаточно рано, поэтому «восхождение» спинного мозга приводит к тому, что корешки удлиняются и изменяют свое направление из горизонтального на косое и даже вертикальное (продольное по отношению к спинному мозгу). Вертикально идущие к крестцовым отверстиям корешки каудальных (нижних) сегментов спинного мозга формируют вокруг концевой нити пучок корешков— так называемый конский хвост, cauda equ-ina.
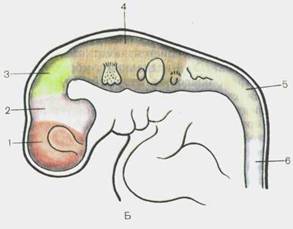
Головной отдел нервной трубки является зачатком, из которого развивается головной мозг. У 4-недельных эмбрионов головной мозг состоит из трех мозговых пузырей, отделенных друг от друга небольшими сужениями стенок нервной трубки. Это prosencephalon — передний мозг,mesencephalon — средний мозги rhomb encephalon— ромбовидный(задний) мозг(рис. 110). К концу 4-й недели появляются признаки дифференциации переднего мозгового пузыря на будущий конечный мозг— telen-cephalon и промежуточный— diencephalon. Вскоре после этого rhomb encephalon подразделяется на задний мозг,metencephalon, и продолговатый мозг,medulla oblongdta, s. bulbus.
Одновременно с формированием пяти мозговых пузырей нервная трубка в головном отделе образует несколько изгибов в сагиттальной плоскости (рис. 111). Ранее других появляется теменной изгиб, направленный выпуклостью в дорсальную сторону и располагающийся в области среднего мозгового пузыря. Затем на границе заднего мозгового пузыря и зачатка спинного мозга выделяется затылочный изгиб, направленный выпуклостью также в дорсальную сторону. Третий изгиб — мостовой, обращенный вентрально, появляется между двумя предыдущими в области заднего мозга. Этот последний изгиб подразделяет ромбовидный мозг на указанные выше два отдела (пузыря): myelencephalon и metencephalon, состоящего из моста, pans, и дорсально расположенного мозжечка, cerebellum. Общая полость ромбовидного мозга преобразуется в IV желудочек, который в задних своих отделах сообщается с центральным каналом спинного мозга и с межоболочечным пространством. Над тонкой однослойной крышей формирующегося IV желудочка прорастают крове-
280
281
|
|
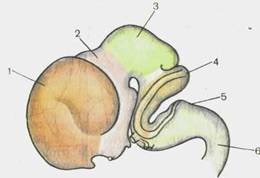
| Рис. 111.Головной мозг эмбриона человека, 8 нед. 1 — telencephalon; 2 — dien-cephalon; 3—mesencephalon; 4 — metencephalon; 5 — myelen-cephalon; 6 — medulla spinalis. |
|
|
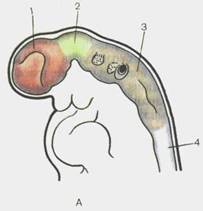
Рис. 110. Головной мозг эмбриона человека на стадиях трех (А) и пяти (Б) мозговых пузырей.
А — З'1/2 нед: 1 — prosencep-halon; 2 — mesencephalon; 3 — rhombencephalon; 4 — medulla spinalis. Б — 4 нед: 1 — telen-cephalon; 2 — diencephalon; 3 — mesencephalon; 4 — rnetencepha-lon; 5 — myelencephalon; 6 — medulla spinalis.
носные сосуды. Вместе с верхней стенкой IV желудочка, состоящей лишь из одного слоя эпендимальных клеток, они образуют сосудистое сплетение IV желудочка,plexus choroideus uentriculi qudrti. В передних отделах в полость IV желудочка открывается водопровод среднего мозга,aqueductus mesencephali (cerebri), который является полостью среднего мозга. Стенки нервной трубки в области среднего мозгового пузыря утолщаются более равномерно. Из вентральных отделов нервной трубки здесь развиваются ножки мозга,pedunculi cerebri, а из дорсальных отделов — пластинка крыши среднего мозга,lamina iecti [tectalisj mesencephali. Наиболее сложные превращения в процессе развития претерпевает передний мозговой пузырь (prosencephalon). В промежуточном мозге (задней его части) наибольшего развития достигают латеральные стенки, которые значительно утол-
щаются и образуют зрительные бугры (таламусы). Из боковых стенок промежуточного мозга путем выпячивания в латеральные стороны образуются глазные пузырьки, каждый из которых впоследствии превращается в сетчатку (сетчатую оболочку) глазного яблока и зрительный нерв. Тонкая дорсальная стенка промежуточного мозга срастается с сосудистой оболочкой, образуя крышу III желудочка, содержащую сосудистое сплетение,plexus choroideus ventriculi tertii. В дорсальной стенке также появляется слепой непарный вырост, который впоследствии превращается в шишковидное тело, или эпифиз,corpus pineale. В области тонкой нижней стенки образуется еще одно непарное выпячивание, превращающееся в серый бугор,tuber cinereum, воронку,infundibulum. и заднюю долю гипофиза,neurohypophysis.
Полость промежуточного мозга образует III желудочек мозга, который посредством водопровода среднего мозга сообщается с IV желудочком.
Конечный мозг, telencephalon, состоящий на ранних этапах развития из непарного мозгового пузыря, впоследствии за счет преобладающего развития боковых отделов превращается в два пузыря — будущие полушария большого мозга. Непарная вначале полость конечного мозга также подразделяется на две части, каждая из которых сообщается с помощью межжелудочкового отверстия с полостью III желудочка. Полости развивающихся полушарий большого мозга преобразуются в имеющие сложную конфигурацию боковые желудочки мозга (рис. 112).
Интенсивный рост полушарий приводит к тому, что они постепенно покрывают сверху и с боков не только промежуточный и средний мозг, но и мозжечок. На внутренней поверхности стенок формирующихся правого и левого полушарий, в области их основания, образуется выступ (утолщение стенки), в толще которого развиваются узлы основания головного мозга — базаль-ные (центральные) ядра, nuclei basdles. Тонкая медиальная стенка пузыря каждого полушария вворачивается внутрь полости бокового желудочка вместе с сосудистой оболочкой и обра-
282
283
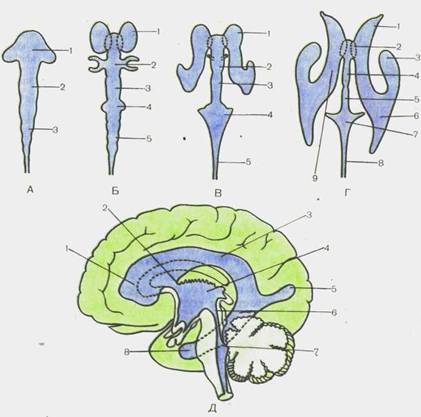
Рис. 112.Развитие желудочков головного мозга у человека (схема). А — полости головного мозга на стадии трех мозговых пузырей (вид сверху): 1 — prosencephalon; 2 — mesencephalon; 3 — rhombencephalon. Б — полости головного мозга на стадий пяти мозговых пузырей (вид сверху): 1 —telencep-halon; 2—diencephalon, 3 — mesencephalon; 4 — metencephalon; 5 — myelen-cephalon. В — образование желудочков головного мозга (вид.сверху): 1—ven-triculus lateralis; 2 — ventriculus tertius; 3 — aqueductus mesencephali; 4 — ventriculus quartus; 5 — canalis centralis. Г — желудочки головного мозга взрослого человека (вид сверху): 1—cornu frontale; 2 — for. interventriculare; 3 — cornu temporale; 4— ventriculus tertius; 5 — aqueductus mesencephali; 6 — cornu occipitale; 7 — ventriculus quartus; 8 — canalis centralis; 9 — pars centralis (ventriculi lateralis). Д — желудочки головного мозга взрослого человека (вид сбоку): 1 —cornu frontale; 2—for. interventriculare; 3 — pars centralis; 4 — ventriculus tertius; 5 — cornu occipitale; 6 —aqueductus mesencephali; 7-ventriculus quartus; 8 — cornu temporale.
зует сосудистое сплетение бокового желудочка. В области тонкой передней стенки, представляющей продолжение терминальной (пограничной) пластинки, развивается утолщение, которое впоследствии превращается в мозолистое тело и переднюю спайку мозга, связывающие друг с другом оба полушария. Неравномерный и интенсивный рост стенок пузырей полушарий приводит
к тому, что вначале на гладкой их наружной поверхности в определенных местах появляются углубления, образующие борозды полушарий большого мозга. Раньше других появляются глубокие постоянные борозды, и первой среди них формируется латеральная (сильвиева) борозда. При помощи таких глубоких борозд каждое полушарие оказывается разделенным на доли, которые более мелкими бороздами подразделяются на выпячивания — извилины большого мозга.
Наружные слои стенок пузырей полушарий образованы развивающимся здесь серым веществом — корой большого мозга. Борозды и извилины значительно увеличивают поверхность коры большого мозга. К моменту рождения ребенка полушария его большого мозга имеют все основные борозды и извилины. После рождения в различных долях полушарий появляются мелкие непостоянные борозды, не имеющие названий. Их количество и место появления определяют многообразие вариантов и сложность рельефа полушарий головного мозга.
Вопросы дляповторения
1. Назовите виды нейронов
2. Что такое рефлекторная дуга? Дайте ее определение.
3. Как классифицируется нервная система анатомически, топографически и но
функциональным признакам?
4. Как называются части головного мозга на стадии пяти мозговых пузырей?
Что развивается из каждого пузыря?
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
СПИННОЙ МОЗГ
Спинной мозг, medulla spinalis, по внешнему виду представляет собой длинный, цилиндрической формы, уплощенный спереди назад тяж (рис. 113). В связи с этим поперечный диаметр спинного мозга больше переднезаднего.
Спинной мозг располагается в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг. В этом месте из спинного мозга (верхняя его граница) выходят корешки, образующие правый и левый спинномозговые нервы. Нижняя граница спинного мозга соответствует уровню I — II поясничных позвонков. Ниже этого уровня верхушка мозгового конуса спинного мозга продолжается в тонкую терминальную (концевую) нить (рис. 114). Терминальная нить, filum terminate, в своих верхних отделах еще содержит нервную ткань и представляет собой рудимент каудального конца спинного мозга. Эта часть терминальной нити, получив-
284
285
|
|
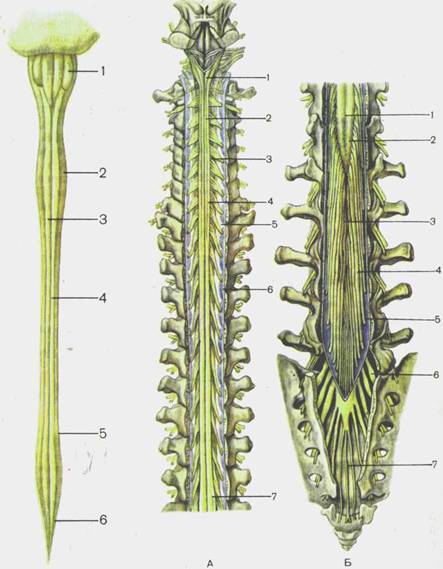
Рис. 113.Спинной мозг, medulla spinalis; передняя поверхность.
1 — medulla oblongata; 2 — intumescentia cervicalis; 3 — fissura mediana ventralis [anterior]; 4 — sul. ventrolateralis [anterolateralis]; 5 — intumescentia lumbosacralis; 6 — conus medullaris.
Рис. 114. Спинной мозг (позвоночный канал вскрыт); вид сзади.
А__ шейно-грудной отдел: 1 — medulla oblongata; 2 — sul. medianus poste
rior; 3 — intumescentia cervicalis; 4 — sul. dorsolateralis [posterolateralis]; 5 —
lig.'denticulatum; 6 — dura mater spinalis; 7 — intumescentia lumbosacralis. Б —
пояснично-крестцовый отдел: 1 —sul. medianus dorsalis [posterior); 2 — conus
medullaris; 3 — filum terminale; 4 — cauda equina; 5 — dura mater spinalis, 6 —
gangl. spiriale; 7 — filum spinale [terminale externum].
шая название внутренней, окружена корешками поясничных и крестцовых спинномозговых нервов и вместе с ними находится в слепо заканчивающемся мешке, образованном твердой оболочкой спинного мозга. У взрослого человека внутренняя часть терминальной нити имеет, длину около 15 см. Ниже уровня П крестцового позвонка терминальная нить представляет собой соединительнотканное образование, являющееся продолжением всех трех оболочек спинного мозга и получившее название наружной части терминальной нити. Длина этой части около 8 см. Заканчивается она на уровне тела II копчикового позвонка, срастаясь с его надкостницей.
Длина спинного мозга у взрослого человека в среднем 43 см (у мужчин 45 см, у женщин 41—42 см), масса — около 34—38 г, что составляет примерно 2 % от массы головного мозга.
В шейном и пояснично-крестцовом отделах спинного мозга обнаруживаются два заметных утолщения: шейное утолщение, intumescentia ceruicalis, и пояснично-крестцовое утолщение, intumesceritia lumbosacralis. Образование утолщений объясняется тем, что от шейного и пояснично-крестцового отделов спинного мозга осуществляется иннервация соответственно верхних и нижних конечностей. В этих отделах в спинном мозге имеется большее по сравнению с другими отделами количество нервных клеток и волокон. В нижних отделах спинной мозг постепенно суживается и образует мозговой конус,conus medulldris.
На передней поверхности спинного мозга видна передняя срединная щель,fissura medidna anterior, которая вдается в ткань спинного мозга глубже, чем задняя срединная борозда,sulcus medidnus posterior. Эти борозды являются границами, разделяющими спинной мозг на две симметричные половины. В глубине задней срединной борозды имеется проникающая почти во всю толщу белого вещества глиальная задняя срединная перегородка, septum medidnum posterius. Эта перегородка доходит до задней поверхности серого вещества спинного мозга.
На передней поверхности спинного мозга, с каждой стороны от передней щели, проходит переднелатеральная борозда,sulcus anterolaterаlis. Она является местом выхода из спинного мозга передних (двигательных) корешков спинномозговых нервов и границей на поверхности спинного мозга между передним и боковым канатиками. На задней поверхности на каждой половине спинного мозга имеется заднелатеральная борозда,sulcus poste-rolaterdlis, место проникновения в спинной мозг задних чувствительных корешков спинномозговых нервов. Эта борозда служит границей между боковым и задним канатиками.
Передний корешок,radix anterior, состоит из отростков двигательных (моторных) нервных клеток, расположенных в переднем роге серого вещества спинного мозга. Задний корешок,radix, posterior, — чувствительный, представлен совокупностью проникающих в спинной мозг центральных отростков псевдо-
287
|
|
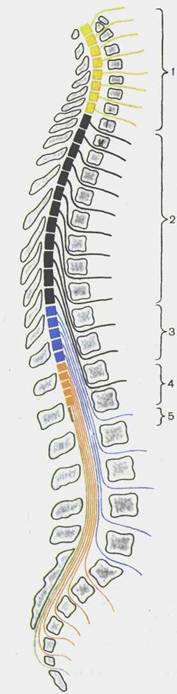
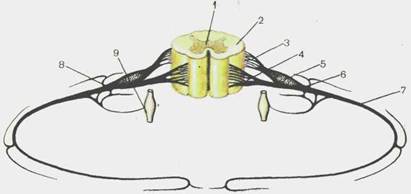
Рис. 115.Сегмент спинного мозга.
1 — substantia grisea; 2 — substantia alba; 3 — radix dorsalis [posterior]; 4- radix ventralis [anterior]; 5 — gangl. spinale; 6 — n. spinalis; 7 — r. ventra-lis [anterior]; 8 - r. dorsalis [posterior]; 9 —gangl. sympathicum.
униполярных клеток, тела которых образуют спинномозговой узел, ganglion spindle, лежащий у места соединения заднего корешка с передним. На всем протяжении спинного мозга с каждой его стороны отходит 31 пара корешков. Передний и задний корешки у внутреннего края межпозвоночного отверстия сближаются, сливаются Друг с другом и образуют спинномозговой нерв, nervus spinalis.
Таким образом, из корешков образуется 31 пара спинномозговых нервов. Участок спинного мозга. соответствующий двум парам корешков (два передних и два задних), называют сег-ментом (рис 115). Соответственно 31 паре спинномозговых нервов у спинного мозга выделяют 31 сегмент: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1—3 копчиковых сегмента. Каждому сегменту спинного мозга соответствует определенный участок тела, получающий иннервацию от данного сегмента. Обозначают сегменты начальными буквами, указывающими на область (часть) спинного мозга, и цифрами, соответствующими порядковому номеру сегмента: шейные сегменты, segmenta cervicdlia, Ci—СVIII; грудные сегменты, segmenta thoracica, Thi -- Thxn; поясничные сегменты, segmenta lumbalia [lumbaria], Li — Lx; крестцовые сегменты, segmenta sacrdlia, Si — Sv; копчиковые сегменты, segmenta coccygea, Coi—CoIII.
Для врача очень важно знать топографические взаимоотношения сегментов спинного мозга с позвоночным столбом (скеле-тотопия сегментов). Протяженность спинного мозга значительно меньше длины позвоночного столба, поэтому порядковый номер какого-либо сегмента спинного мозга и уровень его положения, начиная с нижнего шейного отдела, не соответствует порядке-
Рис. 116. Топография сегментов спинного мозга.
1 — pars cervicalis (Q—Cvm); 2 — pars thoracica (Th,—ThxII); 3 — pars lumbalis (L,— LV); 4 — pars sacralis (S,—SV); 5 — pars coccy-gea (Co,—Com).
вому номеру одноименного позвонка (рис. 116). Положение сегментов по отношению к позвонкам можно определить следующим образом. Верхние шейные сегменты расположены на уровне соответствующих их порядковому номеру тел позвонков. Нижние шейные и верхние грудные сегменты лежат на один позвонок выше, чем тела соответствующих позвонков. В среднем грудном отделе эта разница между соответствующим сегментом спинного мозга и телом позвонка увеличивается уже на 2 позвонка, в нижнем грудном — на 3. Поясничные сегменты спинного мозга лежат в позвоночном канале на уровне тел X, XI грудных позвонков, крестцовые и копчиковый сегменты — на уровне XII грудного и I поясничного позвонков.
Спинной мозг состоит из нервных клеток и волокон серого вещества, имеющего на поперечном срезе вид буквы Н или бабочки с расправленными крыльями. На периферии от серого вещества находится белое вещество, образованное только нервными волокнами (рис. 117).
В сером веществе спинного мозга имеется центральный канал,candlis centrdlis. Он является остатком полости нервной трубки и содержит спинномозговую жидкость. Верхний конец канала сообщается с IV желудочком, а нижний, несколько расширяясь, образует слепо заканчивающийся концевой желудочек, ventriculus terminаlis. Стенки центрального канала спинного мозга выстланы эпендимой, вокруг которой находится центральное студенистое(серое) веще-
288
289
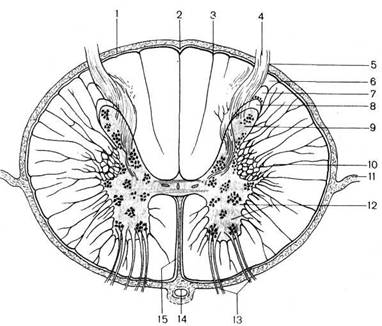
Рис. 117.'Поперечный разрез спинного мозга.
1 — pia mater spinalis; 2 — sul. medianus dorsalis [posterior]; 3 — sul. inter-medius dorsalis (posterior); 4 — radix dorsalis [posterior]; 5 — sul. dorsolatera-lis [posterolateralis]; 6 — zona terminalis (BNA); 7 — zona spongiosa (BNA); 8 — substantia gelatinosa; 9 — cornu dorsale [posterius]; 10 — cornu laterale; 11 - lig. denticulatum; 12 — cornu ventrale [anterius]; 13 — radix ventralis [anterior]; 14 — a. spinalis anterior; 15 — fissura mediana ventralis [anterior].
CTBO, substantia gelatinosa centrdlis. У взрослого человека центральный канал в различных отделах спинного мозга, а иногда и на всем протяжении зарастает.
Серое вещество,substantia grisea, на протяжении спинного мозга справа и слева от центрального канала образует симметричные серые столбы,columnae griseae. Кпереди и кзади от центрального канала спинного мозга эти серые столбы связаны друг с другом тонкими пластинками серого вещества, получившими название передней и задней спаек.
В каждом столбе серого вещества различают переднюю его часть — передний столб, columna ventralis [anterior], и заднюю часть — задний столб, columna dorsalis [posterior]. На уровне нижнего шейного, всех грудных и двух верхних поясничных сегментов (от СVII до Li—LII) спинного мозга серое вещество с каждой стороны образует боковое выпячивание — боковой столб,columna laterdlis. В других отделах спинного мозга (выше VIII шейного и ниже II поясничного сегментов) боковые столбы отсутствуют.
На поперечном срезе спинного мозга столбы серого вещества с каждой стороны имеют вид рогов. Выделяют более широкий передний рог, cornu ventrdle [anterius], и узкий задний рог, cornu dorsdle [posterius], соответствующие переднему и заднему столбам. Боковой рог, cornu, laterdle, соответствует боковому промежуточному столбу (автономному) серого вещества.
В передних рогах расположены крупные нервные корешковые клетки — двигательные (эфферентные) нейроны. Эти нейроны образуют 5 ядер: два латеральных (передне- и заднелатеральное), два медиальных (передне- и заднемедиальное) и центральное ядро. Задние рога спинного мозга представлены преимущественно более мелкими клетками. В составе задних, или чувствительных, корешков находятся центральные отростки псевдоуниполярных клеток, расположенных в спинномозговых (чувствительных) узлах.
Серое вещество задних рогов спинного мозга неоднородно. Основная масса нервных клеток заднего рога образует собственное его ядро. В белом веществе, непосредственно примыкающем к верхушке заднего рога, apex cornus dorsdlis [posterioris], серого вещества, выделяют пограничную зону. Кпереди от последней в сером веществе расположена губчатая зона, которая получила свое название в связи с наличием в этом отделе крупнопетлистой глиальной сети, содержащей нервные клетки. Еще более кпереди выделяется студенистое вещество, substdntia gela-tinosa, состоящее из мелких нервных клеток. Отростки нервных клеток студенистого вещества, губчатой зоны и диффузно рассеянных во всем сером веществе пучковых клеток осуществляют связь с несколькими соседними сегментами. Как правило, они заканчиваются синапсами с нейронами, расположенными в передних рогах своего сегмента, а также выше- и нижележащих сегментов. Направляясь от задних рогов серого вещества к передним рогам, отростки этих клеток располагаются по периферии серого вещества, образуя возле него узкую каемку белого вещества. Эти пучки нервных волокон получили название передних, латеральных и задних собственных пучков, fasciculi proprii, ventrdles [anteriores], laterdles et dorsdles [posteriores]. Клетки всех ядер задних рогов серого вещества — это, как правило, вставочные (промежуточные, или кондукторные) нейроны. Нейриты, отходящие от нервных клеток, совокупность которых составляет центральное и грудное ядра задних рогов, направляются в белом веществе спинного мозга к головному мозгу.
Промежуточная зона серого вещества спинного мозга расположена между передним и задним рогами. Здесь на протяжении с VIII шейного по II поясничный сегмент имеется выступ серого вещества — боковой рог.
В медиальной части основания бокового рога заметно хорошо очерченное прослойкой белого вещества грудное ядро,nucleus thordcicus, состоящее из крупных нервных клеток. Это ядро
290
291
тянется вдоль всего заднего столба серого вещества в виде клеточного тяжа (ядро Кларка). Наибольший диаметр этого ядра на уровне от XI грудного до I поясничного сегмента. В боковых рогах находятся центры симпатической части всгетативнои нерв ной системы в виде нескольких групп мелких нервных клеток" объединенных в латеральное промежуточное (серое) вещество, substantia (grisea) intermedia laterаlis. Аксоны этих клеток проходят через передний рог и выходят из спинного мозга в составе передних корешков.
В промежуточной зоне расположено центральное промежуточное (серое) вещество, substdntia (grisea) intermedia centrаlis, отростки клеток которого участвуют в образовании спинно-моз-жечкового пути. На уровне шейных сегментов спинного мозга между передним и задним рогами, а на уровне верхнегрудных сегментов — между боковыми и задним рогами в белом веществе, примыкающем к серому, расположена ретикулярная формация, formdtio reticulаris. Ретикулярная формация имеет здесь вид тонких перекладин серого вещества, пересекающихся в различных направлениях, и состоит из нервных клеток с большим количеством отростков.
Серое вещество спинного мозга с задними и передними ко-. решками спинномозговых нервов и собственного пучками белого вещества, окаймляющими серое вещество, образует собственный, или сегментарный, аппарат.-.спинного мозга. Основное на-значение сегментарного аппарата как филогенетически наиболее старой части спинного мозга — осуществление врожденных реакций (рефлексов) в ответ на раздражение (внутреннее или внешнее). И. П. Павлов определил этот вид деятельности сегментарного аппарата спинного мозга термином «безусловные рефлексы».
Белое вещество, substdntia alba, как отмечалось, локализуется кнаружи от серого вещества. Борозды спинного мозга разделяют белое вещество на симметрично расположенные справа и слева три канатика. Передний канатик, funiculus ven-trdlis [anterior], находится между передней срединной щелью и передней латеральной бороздой. В белом веществе кзади от передней срединной щели различают переднюю белую спайку, commissura alba, которая соединяет передние канатики правой и левой сторон. Задний канатик, funiculus dorsdlis {posterior} , находится между задней срединной и задней латеральной бороздами. Боковой канатик, funiculus laterdlis, — это участок белого вещества между передней и задней латеральными бороздами.
Белое вещество спинного мозга представлено отростками нервных клеток. Совокупность этих отростков в канатиках спинного мозга составляют три системы пучков (тракты, или проводящие пути) спинного мозга (рис. 118):
1) короткие пучки ассоциативных волокон, связывающие сегменты спинного мозга, расположенные на различных уровнях;
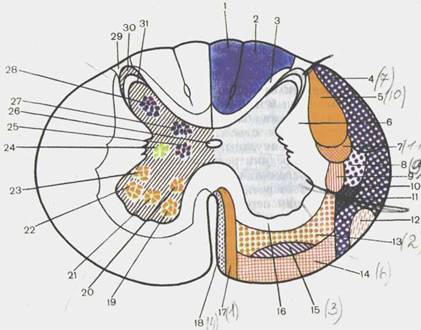
Рис. 118. Проводящие пути белого вещества (1 — 18) и расположение ядер серого вещества (19—28) в спинном мозге; поперечный разрез (схема).
1 — fasc. gracilis; 2 — fasc. cuneatus; 3 — fasc. proprius dorsalis (posterior); 4 — tr. spinocerebellaris dorsaiis (posterior); 5 — tr. corticospinalis (pyramida-lis) lateralis; 6 — fasc. proprius lateralis; 7 — tr. rubrospinalis; 8— tr. spinothala-micus lateralis; 9 — tr. vestibulospinalis dorsalis (posterior] (BNA); 10—tr. spinocerebellaris ventralis [anterior]; 11 — tr. spinotectalis; 12—tr. olivospinalis; 13—tr. reticulospinalis ventralis [anterior); 14 - tr. vestibulospinalis; 15 — tr. spinothalamicus ventralis [anterior]; 16 —fasc. proprius ventralis [anterior]; 17 —tr. corticospinalis (pyramidalis) ventralis (anterior); 18 — tr. tectospinalis; 19— nucl. ventromedialis; 20 — nucl. dorsomedialis; 21 — nucl. centralis; 22-nucl. ventrolateralis; 23 — nucl. dorsolateralis; 24 — columna intermediolateralis (autonomica); 25 — nucl. intermediomedialis (BNA); 26 —canalis centralis; 27-columna thoracica; 28 — nucl. proprius cornu posterior (BNA); 29 — zona termi-nalis (BNA); 30 — zona spongiosa (BNA); 31 - substantia gelatinosa.
2) восходящие (афферентные, чувствительные) пучки, на
правляющиеся к центрам большого мозга и мозжечка;
3) нисходящие (эфферентные, двигательные) пучки, идущие
от головного мозга к клеткам передних рогов спинного мозга.
Две последние системы пучков образуют новый (в отличие от филогенетически более старого сегментарного аппарата) над-сегментарный проводниковый аппарат двусторонних связей спинного и головного мозга. В белом веществе передних канатиков находятся преимущественно нисходящие проводящие пути, в боковых канатиках — и восходящие, и нисходящие проводящие
 292
292
293
пути, в задних канатиках располагаются восходящие проводящие пути.
Передний канатик, funiculus ventrdlis [anterior], включает следующие проводящие пути:
1. Передний корково-спинномозговой (пирамидный) путь, tractus corticospindlis (pyramiddlis) ventrdlis [anterior],— двигательный, содержит отростки гигантских пирамидных клеток (гигантопирамидальный нейрон). Пучок нервных волокон, образующих этот путь, лежит вблизи передней срединной щели, занимая переднемедиальные отделы переднего канатика. Пршюдя-щий путь передает импульсы двигательных реакций от корьТ большого мозга к передним рогам спинного~мозга_.
2. Ретикулярно-спинномозговой путь, tractus reticulospindlis, проводит импульсы от ретикулярной формации головного мозга к двигательным ядрам переднего рога спинного мозга. Он располагается в центральной части переднего канатика, латераль-нее корково-спинномозгового пути.
3. Передний спинно-таламический путь, tractus spinothaldmi-
cus ventrdlis [anterior], находится несколько кпереди от ретику-
лярно-спинномозгового пути. Проводит импульсы тактильной
чувствительности (осязание и давление).
4. Покрышечно-спинномозговой путь, tractus tectospindlis,
связывает подкорковые центры зрения (верхние холмики крыши
среднего мозга) и слуха (нижние холмики) с двигательными
ядрами передних рогов спинного мозга. Он расположен медиаль-
нее переднего корково-спинномозгового (пирамидного) пути.
Пучок этих волокон непосредственно примыкает к передней сре
динной щели. Наличие этого тракта позволяет осуществлять реф
лекторные защитные движения при зрительных и слуховых раз
дражениях.
5. Между передним корково-спинномозговым (пирамидным)
путем спереди и передней серой спайкой сзади расположен зад
ний продольный пучок, fasciculus longitudindlis dorsdlis [poste
rior] .
Этот пучок тянется из ствола мозга до верхних сегментов спинного мозга. Волокна этого пучка проводят нервные импульсы, координирующие, в частности, работу мышц глазного яблока и мышц шеи.
6. Преддверно-спинномозговой путь, tractus vestibulospindlis,
расположен на границе переднего канатика с боковым. Этот
путь занимает место в поверхностных слоях белого вещества
переднего канатика спинного мозга, непосредственно возле его
передней латеральной борозды. Волокна этого пути идут от вес
тибулярных ядер VIII пары черепных нервов, расположенных
в продолговатом мозге, к двигательным клеткам передних рогов
спинного мозга.
Боковой канатик,funiculus laterdlis, спинного мозга содержит следующие проводящие пути: 1. Задний спинно-мозжечковый путь, tractus spinocerebelldris
294
dorsdlis [posterior] (пучок Флексига), проводит импульсы про-приоцептивной чувствительности, занимает заднелатеральные отделы бокового канатика возле задней латеральной борозды. Медиально пучок волокон этого проводящего пути прилежит к латеральному корково-спинномозговому (пирамидному) пути, красноядерно-спинномозговому и латеральному спинно-талами-ческому путям. Впереди задний спинно-мозжечковый путь соприкасается с одноименным передним путем.
2. Передний спинно-мозжечковый путь, trdctus spinocerebelld-
ris ventrdlis [anterior] (пучок Говерса), также несущий про-
приоцептивные импульсы в мозжечок, расположен в переднела-
теральных отделах бокового канатика. Впереди примыкает к
передней латеральной борозде спинного мозга, граничит с оливо-
спинномозговым путем. Медиально передний спинно-мозжечко
вый путь прилежит к латеральному спинно-таламическому и
спинно-покрышечному путям.
3. Латеральный спинно-таламический путь, trdctus spinotha-
lamicus laterdlis, локализуется в передних отделах бокового ка
натика, между передним и задним спинно-мозжечковыми путями
с" латеральной стороны, красноядерно-спинномозговым и пред-
дверно-спинномозговым проводящими путями с медиальной сто
роны. Проводит-импульсы болевой и температурной чувстви
тельности.
К нисходящим системам волокон бокового канатика относятся латеральный корково-спинномозговой (пирамидный) и экстрапирамидный кра-сноядерно-спинномозговой проводящие пути.
4. Латеральный корково-спинномозговой (пирамидный) путь,
trdctus corticospindlis (pyramiddlis) laterdlis, проводит двига
тельные импульсы от коры большого мозга к передним рогам
спинного мозга. Пучок волокон этого пути, являющихся отрост
ками гигантских пирамидных клеток, лежит медиальнее заднего
спинно-мозжечкового пути и занимает значительную часть пло
щади бокового канатика, особенно в верхних сегментах спин
ного мозга. Впереди этого пути находится красноядерно-спинно-
мозговой проводящий путь. В нижних сегментах он на срезах
занимает все меньшую и меньшую площадь.
5. Красноядерно-спинномозговой путь, trdctus rubrospindlis,
расположен кпереди от латерального корково-спинномозгового
(пирамидного) пути. Латерально к нему на узком участке при
лежат задний спинно-мозжечковый путь (его передние отделы) и
латеральный спинно-таламический путь. Красноядерно-спинно-
мозговой путь является проводником импульсов автоматического
(подсознательного) управления движениями и тонусом скелет
ных мышц к передним рогам спинного мозга.
В боковых канатиках спинного мозга проходят также пучки нервных волокон, образующих и другие проводящие пути (например, спинно-покрышечный, оливоспинномозговой и т. д.).
Задний канатик,funiculus dorsdlis [posterior], на уровне шейных и верхних грудных сегментов спинного мозга задней
295
промежуточной бороздой делится на два пучка. Медиальный непосредственно прилежит к задней продольной борозде — это тонкий пучок (пучок Голля), fasciculus gracilis. Латераль-нее его примыкает с медиальной стороны к заднему рогу клиновидный пучок (пучок Бурдаха), fasciculus cunedtus. Тонкий пучок состоит из более длинных проводников, идущих от нижних отделов туловища и нижних конечностей соответствующей стороны к продолговатому мозгу. В него входят волокна, вступающие в состав задних корешков 19 нижних сегментов спинного мозга и занимающие в заднем канатике более медиальную его часть. За счет вхождения в 12 верхних сегментов спинного мозга волокон, принадлежащих нейронам, иннервирующим верхние конечности и верхнюю часть туловища, формируется клиновидный пучок, занимающий латеральное положение в заднем канатике спинного мозга. Тонкий иклиновидный пучки —
это ПРОВОДНИКИ прприоцептивной чувствительности суставно---
мышечное чувство), которые несут в кору полушарий большого мозга информацию о положении тела и его частей в пространстве.
В различных отделах спинного мозга соотношения площадей (на горизонтальных срезах), занятых серым и белым веществом, неодинаковы. Так, в нижних сегментах, в частности в области поясничного утолщения, серое вещество на срезе занимает большую часть. Изменения количественных соотношений серого и белого вещества объясняются тем, что в нижних отделах спинного мозга значительно уменьшается число волокон нисходящих путей, следующих от головного мозга, и только начинают формироваться восходящие пути. Количество волокон, образующих восходящие тракты, постепенно нарастает от нижних сегментов к верхним. На поперечных срезах средних грудных и верхних шейных сегментов спинного мозга площадь белого вещества больше. В области шейного и поясничного утолщений площадь, занимаемая серым веществом, больше, чем в других отделах спинного мозга.
Возрастные особенности спинного мозга
Спинной мозг новорожденного имеет длину 14 см (13,6—14,8), нижняя граница мозга находится на уровне нижнего края II поясничного позвонка. К двум годам длина спинного мозга достигает 20 см, а к 10 годам по сравнению с периодом новорожден-ности удваивается. Наиболее быстро растут грудные сегменты спинного мозга. Масса спинного мозга у новорожденного — около 5,5 г, у детей 1 года — около 10 г. К 3 годам масса спинного мозга превышает 13 г, а в 7 лет равна примерно 19 г.
На поперечном срезе вид спинного мозга такой же, как у взрослого человека. У новорожденного шейное и поясничное утолщения выражены хорошо, центральный канал шире, чем у взрослого. Уменьшение просвета центрального канала происхо-
дит главным образом в течение 1—2 лет, а также в более поздние возрастные периоды, когда увеличивается масса серого и белого вещества. Объем белого вещества возрастает быстрее, особенно за счет собственных пучков сегментарного аппарата, который формируется раньше, чем проводящие пути, соединяющие спинной мозг с головным.
ОБОЛОЧКИ СПИННОГО МОЗГА
Спинной мозг окружен тремя оболочками мезенхимного происхождения (рис. 119). Наружная — твердая оболочка спинного мозга. За ней лежит средняя — паутинная оболочка спинного мозга, которая отделена от предыдущей субдуральным пространством. Непосредственно к спинному мозгу прилежит внутренняя мягкая оболочка спинного мозга. Внутренняя оболочка отделена от паутинной субарахноидальным пространством. В неврологии принято эти две последние, в противоположность твердой мозговой оболочке, называть мягкой оболочкой.
Твердая оболочка спинного мозга, dura mater spindlis, представляет собой продолговатый мешок с довольно прочными и толстыми (по сравнению с другими оболочками) стенками, расположенный в позвоночном канале и содержащий спинной мозг с передними и задними корешками спинномозговых нервов и остальными оболочками. Наружная поверхность твердой мозговой оболочки отделена от надкостницы, выстилающей изнутри позвоночный канал, надоболочечным эпидуральным пространством, cdvitas epidurdlis [spdtium epidurale]. Последнее заполнено жировой клетчаткой и содержит внутреннее позвоночное венозное сплетение. Вверху, в области большого затылочного от-верстия твердая оболочка спинного мозга прочно срастается с краями большого затылочного отверстия и продолжается в твер-дую оболочку головного мозга. В позвоночном канале твердая оболочГка укреплена при помощи отростков, продолжающихся в периневральные оболочки спинномозговых нервов, срастающихся с надкостницей в каждом межпозвоночном отверстии. Помимо этого, твердую оболочку спинного мозга укрепляют многочисленные фиброзные пучки, направляющегося от оболочки к задней продольной связке позвоночного столба
Внутренняя поверхность твердой оболочки спинного мозга отделена от паутинной узким щелевидным субдуральным пространством, которое пронизано большим количеством тонких пучков соединительнотканных волокон. В верхних отделах позвоночного канала субдуральное пространство спинного мозга свободно сообщается с аналогичным пространством в полости черепа. Внизу его пространство заканчивается слепо на уровне II крестцового позвонка. Ниже пучки волокон, принадлежащие твердой оболочке спинного мозга, продолжаются в терминальную (наружную) нить.
Дата добавления: 2018-05-12; просмотров: 1456; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!