Эволюция гаметофитов у высших споровых и семенных. Предпосылки и появление семени
Спорофит - растение, образующее споры. Споры появляются в многоклеточных спорангиях в результате мейоза. Они распространяются ветром, водой, животными. У равноспоровых растений все споры одинаковые по размеру. Более высокоорганизованные растения разноспоровые: в микроспорангиях у них образуются многочисленные мелкие споры - микроспоры, а в мегаспорангиях - крупные мегаспоры. При прорастании спор образуются гаметофиты, поэтому гаметофиты гаплоидны.
Гаметофит - растение, образующее гаметы. Мужские гаметы - сперматозоиды образуются в мужских половых многоклеточных органах - антеридиях, похожих на мешочки, а женские яйцеклетки - в колбообразных архегониях (рис. 6.1). Оплодотворение яйцеклеток происходит при наличии капельно-жидкой среды, необходимой для движения сперматозоидов. После оплодотворения образуется дипло- идная зигота, формирующая многоклеточный зародыш. Зародыш со временем развивается в новый спорофит.
В жизненном цикле только у моховидных доминирует гаметофит, у всех остальных высших растений - спорофит. Гаметофит еще называют заростком. Он имеет вид небольшой пластинки (несколько миллиметров) или клубенька, без дифференциации на органы, прикрепляющегося к почве с помощью ризоидов. Вся эволюция высших растений была направлена на редукцию гаметофита и усовершенствование спорофита.
Рассмотрим процесс появления семени на примере группы растений, которая в современном мире является господствующей. Это представители отдела Покрытосеменные. Все они формируют цветок - важнейший генеративный орган. В его пестике располагается яйцеклетка, а пыльники тычинок содержат спермии. После процесса опыления, т.е. переноса пыльцы с пыльника тычинок на рыльце пестика, спермии по зародышевой трубке продвигаются в завязь тычинки, где и происходит процесс слияния гамет - оплодотворение. В результате формируется зародыш. При слиянии второго спермия с центральной зародышевой клеткой образуется запасное питательное вещество. Его еще называют эндоспермом. Завершает строение семени прочная наружная оболочка. Такая структура является основой для развития будущего растительного организма.
Внешнее строение семян
Как уже было сказано, снаружи семя покрыто кожурой. Она достаточно плотная, чтобы защитить зародыш, находящийся внутри, от механических повреждений, перепадов температур и проникновения вредных микроорганизмов. А вот цвет семян варьируется в широких пределах: от черного до ярко-красного. Такое строение семени легко объяснить. У одних растений цвет служит для маскировки. Например, чтобы птицы не смогли рассмотреть их в почве после посадки. Другие растения, наоборот, приспособлены к распространению семян при помощи различных животных. Вместе с непереваренными остатками пищи они выделяют их далеко за пределами ареала произрастания материнского растения.
Внутреннее строение семени
Основной частью любого семени является зародыш. Это и есть будущий организм. Поэтому он состоит из тех же частей, что и взрослое растение. Это зародышевый корешок, стебелек, листик и почечка. Строение семени разных растений может существенно отличаться. У большинства из них запасные питательные вещества накапливаются в эндосперме. Это оболочка, которая окружает зародыш вокруг, защищая и питая его в течение всего периода индивидуального развития. Но бывают случаи, когда во время процесса созревания и прорастания семени оно полностью расходует вещества эндосперма. Тогда они накапливаются в основном в мясистых частях зародыша. Они называются семядолями. Такое строение характерно, например, для тыквы или фасоли. А вот у пастушьей сумки запас веществ сконцентрирован в ткани зародышевого корешка. Отличаются и семена различных систематических групп растений.
Роль полимеризации и ароморфных надставок
13 А. Д. Заморским (19716) выделено два вида полимеризации; организмов — неполная и полная. Первая сопровождается увеличением количества частей тела, органов и органоидов в пределах организма, вторая — умножением числа самих организмов. У растений, ввиду нечеткости у них понятия индивид, границы между двумя видами полимеризации стираются. Поэтому^ у растений можно выделить еще две категории полимеризации:: количественную и качественную. Количественная полимеризация не приводит к образованию нового качества (простое размножение, образование примитивных колоний, увеличение количества частей тела). Качественная полимеризация ведет к образованию нового качества, например многоклеточных организмов из одноклеточных, сверхмногоклеточных из многоклеточных, новых органов на основе полимеризации (соцветий типа* псевданциев, синкарпной завязи и т. д.). В этом смысле качественная полимеризация является одной из форм интеграции ж невозможна без количественной. Переход последней в первую— есть отражение диалектического закона перехода количества* в качество. Переход количественной полимеризации первичных вирусоподобных биологических систем в качественную — это процесс, при котором за счет объединения вирусных частиц достигалась большая надежность системы. Необходимость в повышении надежности системы возрастала по мере истощения растворенных в воде полимеров. Следовательно, в эволюции низших растений были следующие этапы качественной полимеризации.
1. Образование нитчатых (простых и ветвистых) форм из одноклеточных.
2. Образование пластинчатых фом, представляющих собой пластинки из одного-двух слоев клеток как из нитчатых, так и из одноклеточных. ,
3. Трехмерные формы, являющиеся как бы совокупностью нескольких пластинок. Пример плодовитых тел высших грибов показывает, что они могли образоваться и непосредственно благодаря полимеризации нитей, но все же не отдельных клеток. Вольвоксовые водоросли, колонии которых бывают шарообразными, имеют живые клетки только по поверхности и развиваются как пластинчатые образования, становящиеся все более вогнутыми.
Следовательно, растения прошли шесть ступеней качественной полимеризалии: одноклеточную, нитчатую, пластинчатую, талломную, предпобеговую (таллитную) ипобеговую (см. рис. 1). В эволюции жизненных форм семенных полибластических растений можно различить следующие этапы полимеризации: образование кустовидных форм из одноосных, клонообразующих из кустовидных. Уменьшение степени полимеризации возможно путем редукции средних, а не конечных или начальных ступеней (этапов).
Этапы качественной полимеризации являются этапами и мерилом прогресса. Они запечатлены в геологической летописи и основном биогенетическом законе, в этапах онтогенетического развития. Онто- и филогенез осуществляются прибавлением к предыдущим, менее полимеризованным стадиям последующих, все более и более полимеризованных. Основной, ароморфный, путь эволюции онтогенезов — это путь надставок, анаболии.
Количественная полимеризация, помимо увеличения степени надежности, ведет к грандизации, увеличению размеров системы. Это, как и повышение энергетического уровня, имеет немаловажное значение в борьбе за существование. Качественная полимеризация так же приводит к грандизации, но не столь явно. Известно, например, что некоторые многоклеточные (коловратки) мельче многих одноклеточных, псилофиты были значительно мельче, чем талломы крупных водорослей. Встречаются цветковые эфемеры, которые намного мельче мхов. Основной, ароморфный, путь эволюции онтогенезов — это путь надставок, анаболии.
14. Высшие растения, или Наземные растения, или Эмбриофиты (лат.Streptophyta, включают группу без ранга лат. Embryophyta) — тип зелёных растений, которым свойственна дифференциация тканей, в отличие от низших растений — водорослей. К высшим растениям относятся мхи и сосудистые растения (папоротникообразные, псилотовые, хвощевидные, плауновидные, голосеменные и покрытосеменные).
Высшие растения в некоторых системах классификации рассматривают как таксон ранга подцарство.
Эволюция высших растений тесно связана с выходом на сушу и завоеванием наземных ниш.
Развитие специализированных тканей было важным условием для выхода растений на сушу. Для комфортного существования в воздушной среде растениям было необходимо развить как минимум эпидермис с устьицами для защиты от высыхания и теплообмена и проводящие ткани для обмена минеральных и органических веществ. Результатом выхода растений на сушу также стало разделение организма растения на корень, стебель и лист.
1. Мхи
Среди высших растений наиболее примитивным строением обладают Мохообразные (Bryophyta sensu lato) — у них отсутствует корень (есть ризоиды), а у маршанциевых, антоцеротовых и некоторых юнгерманниевых мхов отсутствует деление на лист и стебель — они являются слоевищными, как водоросли или лишайники. Устичный аппарат крайне примитивен, проводящая система не развита, проводящие функции выполняет паренхима.
2. Сосудистые споровые
.
Так называемые Сосудистые споровые, к которым относятся папоротники, хвощи, плауны и псилотовые, имеют уже довольно развитую проводящую систему, всегда выражены стебель, лист и корень. Однако они все еще сильно связаны с водной средой, так как имеют свободноживущий гаметофит и половое размножение у них происходит с участием жгутиконосных сперматозоидов, которые не могут существовать вне водной среды.
Если Мохообразные и Сосудистые споровые рассматривают как единую группу, к ней иногда применяют собирательный термин «высшие споровые растения»[2].
3. Семенные растения
Важным эволюционным прорывом растений на пути завоевания суши стало появление семени и оболочки пыльцевого зерна. Благодаря тому, что отныне гаметофит (теперь состоящий всего из нескольких клеток) стал полностью помещаться внутри влагоудерживающей оболочки, растения смогли освоить пустынные и холодные области.
У некоторых голосеменных и практически у всех цветковых растений в проводящих структурах появляются сосуды и ситовидные трубки — полые проводящие элементы, состоящие из стенок отмерших клеток, благодаря чему их проводящие системы действуют чрезвычайно эффективно.
К высшим относят растения следующих отделов:
· Моховидные (Bryophyta)
· Плауновидные (Lycopodiophyta)
· Хвощевидные (Equsetophyta)
· Папоротниковидные (Polypodiophyta)
· Голосеменные (Gymnospermae или Pinophyta)
Покрытосеменные или Цветковые (Angiospermae или Magnoliophyta) Сосу́дистые расте́ния (лат. Tracheophyta, Plantae Vasculares, Tracheobionta) — обширная группа растений, органы которых снабжены сосудисто-волокнистыми пучками. К этой группе относятся все высшие растения за исключением мохообразных[1]. Остальные растения, не имеющие сосудистых пучков, называются клеточными (лат.Plantae Cellulares). Это деление предложено Огюстеном Пирамом Декандолем в 1813 году. К сосудистым растениям относятся как высшие споровые (Папоротниковидные и Плауновидные), так и все семенные растения (Гинкговидные, Гнетовидные, Саговниковидные, Хвойные и Покрытосеменные). Отдельные группы сосудистых растений отличаются друг от друга особенностями диплоидных и гаплоидных фаз онтогенеза.
15.
Отдел Моховидные. В этот отдел входит более 25 тыс. видов. Корней нет. Спорофит самостоятельно не существует, развивается и всегда находится гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий на ножке. Отдел включает класс листостебельные мхи, класс печеночники и антоцеротовые.
Отдел Плауновидные – очень древняя группа, к которой относятся как ископаемые, так и ныне живущие растения. Имеют длинные стелющиеся дихотомически ветвящиеся побеги, густо усаженные жесткими мелкими листьями.
Отдел хвощевидные. Многолетние травы лугов, болот, лесов и полей. От корневищ отходят придаточные корни и надземные побеги, имеющие типичное членистое строение. От узлов отходят мутовки бурых чешуйчатых листьев, сросшихся в трубчатое влагалище, и мутовки боковых побегов.
Отдел папоротниковидные. Преобладание долговечного листостебельного спорофита над эфемерным примитивным гаметофитом, наличие у спорофита крупных, обычно перисто-рассеченных, сложно устроенных листьев – вай, расположение спорангиев группами (сорусами) на нижней стороне листьев
16.
Надотдел Bryomorphae ( Bryophyta) – мохообразные
Мохообразные – уникальная группа высших растений, в жизненном цикле которых господствует гаметофит, спорофит живет на материнском гаметофите и питается за его счет.
Характерно: - слабое развитие кутикулы ( или она отстуствует), мхи свободно поглощают влагу всей поверхностью тела и легко ее отдают - проводящая система мхов не имеет ситовидных клеток и трахеид - корней нет( имеются ризойды) - по этой причине мохообразные сохраняют древний пассивный способ поглощения воды за счет капиллярности и гигроскопичности ( т.е. они пойкилогидридные растения), настоящих типичных устьиц нет - листья отсутствуют и их функцию выполняет таллом или филлоиды (листоподобные структуры энационного происхождения) - особый замедленный тип фотосинтеза; его низкая продуктивность обусловлена мелкими хлоропластами с пониженным содержанием хлорофилла а по сравнению с хлорофиллом b - способность легко впадать в состояние анабиоза и пережидать неблагоприятные условия
Bryomorphae – древняя группа высших растений, известная с корнца девона – начала карбона, их изучает бриология - в настоящее время насчитывается от 16 до 22 тыс. видов мхов, максимальное их многообразие в тропиках, но доминируют в растительном покрове они также и в горных и приполярных районах, - предполагается, что разные группы мохообразных произошли от различных предковых форм, или от зеленых водорослей – обладателей фрагмобластемы, или от древних форм риниевых; т.е. мохообразные – полифилетическая группа высших растений.
Особенности строения гаметофита: - гаметофит мохообразны – самый сложно организованный среди других высших растений; - взаимоотношение материнского гаметофита и спорофита не паразитического характера, а напоминают взаимодействие организма матери и плода у плацентарных млекопитающих - начальная стадия развития гаметофита называется протонемой, строение которой специфично для разичных групп мохообразных - развитие протонемы связано с делением единственной инициальной ( апикальной) клетки, которая м.б. различного строения и делится по-разному у различных групп мохообразных.
Классификация мохообразных: подразделяются на 4 класса ( отдела) - отдел печёночники – Divisio Hepaticae или Marschantiophyta - отдел Антоцеротовые мхи – Divisio Anthocerotophyta - отдел Такакиевые мхи – Divisio Takakiophyta - отдел листостебельные мхи – Divisio Bryophyta или Musci
17.
Отдел печёночники – Divisio Hepaticae или Marschantiophyta насчитывает около 6-8 видов, большинство обитает в тропиках. Характерные особенности отдела: - колоссальное морфологическое разнообразие гаметофитов ( есть талломные и листостебельные формы) и значительное сходство спорофитов - дорзовентральность в строении тела гаметофита: на брюшной стороне имеются ризойды( у толломных форм) или брюшные чешуйки- амфигастрии ( у листостебельных) - листья всегда однослойные и состоят из однородных клеток - в коробочках спорогонов имеются элатеры ( разрыхлители) или питательные клетки - протонема слабо развита и часто мало отличается от гаметофита - в отличии от других мхов, у всех печеночников имеется симбиотическая микориза, а иногда и симбиотические цеанобактерии - печеночники – монофилитическая группа, однако их предки до сих пор не установлены.
Систематика печеночников: - класс Сферокарповые – Classic Sphaerocarpopsida - класс Маршанциевые – Classic Marschantiopsida - класс Моноклеевые - Classic Monocleopsida - класс Мецгериевые – Classic Metzgeriopsida - класс Калобриевые – Classic Calobriopsida - класс Трейбиевые – Classic Treubiopsida - класс Юнгермсанниевые – Classic Jungermanniopsida
1 класс – Сферокарповые чуть более 30 видов, микроскопические по размерам, наземные и водные печеночники с побеговой структурой, обитают в районах со средиземноморским климатом и проходят ускоренный цикл развития за период теплой зимы или быстро пересыхающих тропических водоемах с солоноватой водом
2 класс –Маршанциевые 13 семейств, примерно с 400 видов, тело представлено талломом различного строения, который однако из-за упорядоченного расположения брюшных чешуек можно рассмотреть и как уплощенный побег, большинство одно- и многолетние растения, преимущественно гелиофиты, обитают на различных субстратах, в том числе на скалах, могут легко переносить засуху ( криптобиоз или в виде спор).
3 класс – Моноклеевые представлен одним родом – Monoclea, в котором различают 1-2 вида, морфолого-анатомические и биохимические признаки несут черты, характерные для других групп печеночных мхов, обитает в тенистых сырых местах, по берегам ручеёв и мелких луж на влажной почве и гриющих остатках
4 класс - Мецгериевые относясятся к классу около 650 видов, встречаются как талломные так и побеговые форсы, характерны симбиотические взаимосвязи с грибами и цианеями
5 класс – калобриевые относится один род Haptomitrium, и 15 видов, побеговые формы с оригинальным строением, наличие корневищ без ризойдов, каралловидный облик микотрофность, протонема клубневидная, листья гомологичны слизевым волоскам, архегонии и антеридии примитивного типа, характерна двудомность, на спорофите имеются небольшие придаточные зеленые листочки (парафиллы), большинство обитают в горах экваториального пояса
6 класс – Трейбиевые 7 видов Treubia рода, и2 вида Apotreubia рода, тело гаметофита может быть рассмотрена как побеговая структура, а может быть как уплощенный стеблеобразный таллом с боковыми листовидными и спинными чешуевидными выростами, листья в онтогенезе возникают из клеток, формирующих слизевые волоски, т.е. гомологичны им, гаметофиты двудомные , многочисленные гаметангии располагаются группой в пазухе спинной доли листа, на спорофитек имеются небольшие зеленые листочки, обитают на влажной хорошо гумусированной почве, распространены на островах Тихого океана
7 класс – Юнгерманниевые 5 – 7 тыс.видов, около 300 родов, гаметофит представлен листостебельными стелющимися формами с двумя рядами боковых и одним рядом брюшных листьев или амфигастриев, листья тонкие, однослойные, энационного происхождения, иногда они редуцированы и побег состоит из одного уплощенного стебля, протонема нитчатая, разнонитчатая или пластинчатая, встречаются неотеническиеформы(гаметофит гетеротрихальный)Э, нередко размножаются вегетативно, антеридии и архегонии верхушечные, архегонии имеют специальные защитные образования (перихеции и периантии ), могут быть одно- двух- и мн6огодомные растения, Спорофит состоит из коробочки ножки и гаустории (гаустория может прорастать сквозь гаметофит и питаться самостоятельно за счет собств. Ризойдов), у большинства коробочка вскрывается 4-мя створками, имеются микрофильные симбиотические формы, а также эпифильные полупаразиты, с эволюционной точки зрения мхи более специализированные и диверсифицированные среди печеночников, распространены по всему миру, но много в тропиках.
18.
Антоцеротовые мхи – Divisio Anthocerotophyta отдел представлен одним классом Anthocerotopsida примерно 100 – 150 видов, Таллом гаметофиты пластинчатый, большей частью розетковидный формы, краевая меристема формирует лопасти, которые налегают друг на друга, Хлоропласты, содержащиеся в верхних слоях клеток таллома, как правило одиночные и с особой архитектурной тилакоидной системы, не встреающейся у других высших растений, содержат периноиды, протонема слабо развита и представлена одиночной нитью, гаметофит сособен к регенерации , иногда в клетках гаметофита встречается микориз0а и симбиотические цианобактерии, спорофит очень освоеобразен: состоит из гаустории и длинной узкой коробочки без ножки ,
нарастающей интеркалярно; эпидермис без хлоропластов, но есть устьица с замыкающими хлорофильными клетками, хлорофиллоносны и клетки субэпидермиса, гаустория иногда проростает сквозь пластинку гаметофита, внедряется в почву, развивая на своей поверхности ризойды(предполагали, что когда-то спорофит антоцеротовых мхов вел свободный образ жизни), однако считать спорофит вероятным предком высших споровые нельзя . произрастают во всех зонах.
19.
Отдел Листостебельные мхи – Divisio Bryophyta или Musci самый многочисленный отдел мохообразных: насчитывает 10-12000 видов, объединенных в 800 родов и 100 семейств. - характерна листостебельность, радиальная симметрия; ризойды всегда многоклеточны. - листья ( филлоиды) однослойны (Shagnum) или многослойны энационного происхождения. - протонема резко отличается от взрослого гаметофита, имеет вид ветвящейся нити или пластинки. - коробочка спорогона имеет сложное строение – состоит из урночки и крышечки, которая отбрасывается и способствует активному распространению спор, элатера отсутствует. - подавляющее большинство листостебельных мхов в отличии от других мохообразных приурочены не к тропическим , а умеренным и холодным облостям, имеются вторичноводные и много олиготрофов. Классификация: Класс Сфагновые – Sphagnopsida Класс андреэевые – Andreaepsida Класс Эдиподиевые – Oedipodiopsida Класс Тетрафисовые – Tetraphidopsida Класс Политриховые – Polytrichopsida Класс Буксбаумиевые – Buxbaumiopsida Класс Бриевые – Bryopsida
Сфагновые мхи ( Sphagnopsida) – представлены одним порядком (Sphagnales) одним семейством (Sphagnaceae), одним родом (Sphagnum), в роде свыше 300 видов ( В России 42), которые морфологически трудно различить, в основном отличаются анатомией листьев и стеблей - широко распространены в умеренной зонеСеверного полушария - характеризуются высочайшей степенью специализациигаметофита, быстро поглащающего воды и прочно удерживающего её, механизм поглащения – за счет гигроскопичности, капиллярности и набухания 9пассивный принцип) – как и юнгерманиевые имеет место тонкая специализация на базе невысокого уровня эволюционного развития. Сфагнум — болотный мох, впитывает воду всем телом; ризоидов нет. Для него характерны особые водозапасающие клетки на листьях и стебле (прозрачные, мёртвые, полые с отверстиями); клеточная стенка укреплена утолщениями. Водозапасающие клетки окружены более мелкими зелёными фотосинтезирующими клетками, которые объединены в единую сеть[1]. Имеются ножка и коробочка со спорами. Тело сфагнума содержит карболовую кислоту, которая является антисептиком, убивающим бактерии. В связи с этим мох почти не гниёт и образует торф (по 1—2 мм в год). За счёт роста сфагнума и других водных растений происходит заболачивание лесов и зарастание водоёмов: озёра превращаются в болота. Из-за малой теплопроводности употребляется в строительном деле как изоляционный материал в виде пластинок, порошка, изготовляемого из этого торфа; также дезодорирующее средство. Сфагнум применяется в цветоводстве в качестве наполнителя при составлении земляных смесей.Перевязочный материал, Верхние части растения используют в качестве лекарственного сырья. Сфагнум содержит фенольное соединение сфагнол и другие фенольные и тритерпеновые вещества. Встречаются в Волгоградской области: Sphagnum centrale (Сфагнум центральный) Sphagnum compactum (Сфагнум компактный) Sphagnum fallax (Сфагнум обманчивый) Sphagnum squarrosum (Сфагнум оттопыренный) Sphagnum platyphyllum (Сфагнум плосколистный) Sphagnum girgensohnii (Сфагнум Гиргензона)
Андреевые мхи (Andreaepsida) – представлены одним семейством ( Andreaeceae) с двумя родами – Andreaea(70-120 ыидов) и Andreaeobryum (1 вид) -споры прорастают под экзиной; листья однослойные, вырастют верхушкой - коробочка оригинального строения , вскрывается 4-мя щелями - виды Andreaea обитают в умеренных и холодных зонах Сев. И Южн полушарий, в тропиках(горы), на силикатных скалах. - вид Andreaeobryum произрастает на Аляске и в Зап. Канаде (известянковые скалы) -древняя группа мхов, проявляющая филогенетические связи с печеночниками, сфановыми и такакиевыми.
20.
Отдел ЛИСТОСТЕБЕЛЬНЫЕ МХИ, или собственно МХИ (BRYOPHÝTA). Класс ПОЛИТРИХОВЫЕ МХИ. Строение, биология, экология. Цикл развития зеленого мха кукушкина льна. Хозяйственное значение политриховых мхов. Представители класса, обитающие в Волгоградской области. Общая характеристика Класса БУКСБАУМИЕВЫЕ МХИ.
Отдел Листостебельные мхи - самый многочисленный отдел мохообразных. Характерна листостебельность, радиальная симметрия, ризоиды всегда многоклеточны. Листья (филлоиды) однослойны (сфагнум) или многослойны, энационного происхождения. Протонема резко отличается от взрослого гаметофита, имеет вид ветвящейся нити или пластинки. Коробочка спорогона имеет сложное строение – состоит из урночки и крышечки, которая отбрасывается и способствует активному распространению семян. Элатеры отсутствуют. Подавляющее большинство листостебельных мохообразных, приурочены не к тропическим, а к умеренным и холодным областям (леса, тундры), имеются вторично водные виды: много олиготрофов. Отдел включает классы: Сфагновые, Андреэевые, Эдиподиевые, Тетрафиеовые, Политриховые, Буксбаумиевые, Бриевые.
Класс Политрихиевые. По-видимому, представителей этого класса следует считать одной из самых продвинутых в эволюционном отношении группой листостебельных. Имеют высокоспециализированное анатомическое строение побегов (стеблей, а особенно корневищ), сравнимое в таковым у высших сосудистых растений. Спорогон и коробочка устроены очень сложно: перистом крайне оригинален и высоко эффективен. Широко распространенная политопная группа, занимающее разнообразные места обитания. Способны создавать доминантные сообщества, играют заметную роль в специфических сообществах. Возникли, вероятно, в палеозое во влаж.местах Пангеи. В их эволюции большую роль играли анаболия и протогенез (связан с освоением пионерских местообитаний)
ЖЦ Кукушкиного льна. На верхушке стебелька мха весной образуются половые органы: на одних особях мужские — антеридии, на других женские — архегонии.
В антеридиях формируются в большом числе двужгутиковые сперматозоиды. В архегониях образуется одна яйцеклетка. В сырую погоду, во время дождя, росы, антеридий вскрывается на верхушке и сперматозоиды выплывают наружу. К моменту созревания яйцеклетки архегоний также вскрывается на верхушке. Сперматозоиды, подплывая к архегонию, проникают к яйцеклетке, и один из них сливается с ней, производя оплодотворение. Оплодотворенная яйцеклетка (зигота) начинает делиться, и в дальнейшем формируется новое растенье, представленое коробочкой, сидящей на ножке. Ножка своим основанием внедряется в верхушку стебля, всасывая необходимые для развития коробочки питательные вещества. В таком виде растение уходит под снег. Следующей весной ножка сильно удлиняется и выносит коробочку высоко над покровными листьями. Взрослая коробочка имеет следующее строение: сверху она прикрыта крышечкой, средняя часть называется урночкой, нижняя, слегка расширенная после перетяжки, называется апофизой (шейкой коробочки). Внутри коробочки, в центре ее, проходит тяж бесплодных клеток — колонка, наверху расширяющаяся в эпифрагму. Со всех сторон вокруг колонки висит на нитях спорангий, в котором формируются споры.
По созревании спор (начало — середина лета) колонка и стенка спорангия разрушаются, и споры лежат в полости урны. Затем сбрасывается крышечка и по верхнему краю урночки обнаруживается так называемый перистом, состоящий из зубцов, расположенных в один ряд. Вход в урночку закрыт пленкой (эпифрагмой). Зубцы перистома очень гигроскопичны и меняют свое положение в зависимости от влажности воздуха. Во влажную погоду они загибаются внутрь, прижимая пленку к краям урны, что препятствует проникновению воды в коробочку. В сухую погоду зубцы перистома отгибаются наружу, пленка ссыхается и споры высеиваются наружу через отверстия между зубцами.
Спора, попав на сырую землю, прорастает и образует обильно разветвленную нить — протонему, на которой возникают почки, развивающие затем листостебельные растеньица мха.
Таким образом, у мхов имеется «смена поколений». Листостебельное растение (сам мох), образующее половые органы (гаметофит), сменяется коробочкой на ножке, внутри которой образуется спорангий со спорами (спорофит), причем это новое поколение не порывает связи с гаметофитом. Редукционное деление происходит при образовании спор в спорангии коробочки.
В цикле развития преобладает гаметофит (сам мох). Спорофит слабо развит и имеет небольшие размеры и кратковременное существование. У некоторых мхов на коробочке развивается ассимиляционная ткань с устьицами, но основное питание спорофит получает от гаметофита.
Кукушкин лён применяют при возведении деревянных домов как межвенцовый утеплитель. В народной медицине кукушкин лен используется, как мочегонное и слабительное.
Класс Буксбаумиевые мхи. 2 небольших семейства: монотипное Buxbaumiaceae (10 видов) и олиготипное Diphyseiaceae (3вода, 20 видов). Протонема гетеротрихиальная (многолетняя), способна размножаться вегетативно за счет распада на отдельные клетки. Гаметофит (особенно мужск) предельно редуцирован, питается за счет развития вторичной протонемы. Спорофит имеет сложно дифференцированную коробочку с в той или или иной степени редуцированным перистомом и оригинальным механизмом выброса спор («спринцовка»). Род Buxbaumia – в северн.полушарии (бореальн.леса с океанич.климатом). Diphyseiaceae – в горных океанич.районах Ю.-В Азии, Юж и Сев. Америки, 2 вида – на востоке и западе Евразии. Достаточно продвинутая специализированная группа, в эволюции которой большую роль играл протогенез (формирование органов и их системна все более ранних стадиях онтогенеза), что связано с освоением пионерных местообитаний, где условия жизни неблагоприятны и осоаблена конкуренция. Родственно и эволлюционно связаны с Политриховыми мхами.
21.
Отдел ЛИСТОСТЕБЕЛЬНЫЕ МХИ, или собственно МХИ (BRYOPHÝTA). Класс БРИЕВЫЕ МХИ. Особенности строения, биологии и экологии. Бриевые мхи Волгоградской области.
Отдел Листостебельные мхи – См. предыдущий вопрос. Класс Бриевые. Самый крупный класс мхов (10-12 тыс видов и около 800 родов). Традиционно делятся на 2 группы – верно- и бокоплодных мхов. Протонема гетеротрихиальная, одно-, двумерная (редко трехмерная), ее клетки часто специализированы. Стебли с проводящими пучками, листья одно-, многослойные, характерны парафиллы и псевдопарафиллы. Размножаются преимущественно спорами, но могут и вегетативно. Спорофит и коробочка устроены очень сложно, строение перистома имеет важное таксономическое значение. Космополиты, освоили все возможные местообитания. Высоко специализированная группа – вершина эволюции мохообразных. Листья большей частью с жилкой. У большинства родов коробочка вскрывается посредством отпадения крышечки, а по краю образовавшегося в урночке отверстия развивается перистом. Распространены по всему миру от тундры и лесотундры до степей и пустынь. В Волгоградской области: Брий, или Бриум серебристый. Ортотрихум бледноватый. Гигроамблистегиум разнообразный
22.
Низшие TRACHEOPHÝTA – сосудистые споровые растения. Общая характеристика PROTRACHEOPHÝTA – древнейших ископаемых сосудистых споровых растений. Характеристика важнейших представителей, их эволюционное значение.
Группа Tracheophyta объединяет весьма разнообразные растения как минимум трех отделов. Считается, что у всех Tracheophyta в жизненном цикле преобладает спорофаза и всегда самостоятельно существующий спорофит имеет более сложное строение, чем гаметофит. Такое соотношение поколений вероятнее всего было присуще и подавляющему большинству ископаемых форм Tracheophyta. У примитивных представителей автотрофные спорофиты имели теломную организацию, а у продвинутых форм есть побеги и корни. Большинство форм имеет устьица, но некоторые вторично их утратили. Спорофитам свойственна сложная гистологическая дифференциация, лишь у немногих видов утраченная вторично в связи с водным или паразитическим образом жизни.
Спорофиты Tracheophyta способны синтезировать суберин. Лигнин откладывается в клеточной стенке. На базе этой способности спорофиты сосудистых растений выработали специализированные механические элементы — склереиды разных типов и волокна, представленные в теле растения идиобластами или массивами особой ткани — склеренхимы. Лигнификация клеточных стенок лежит и в основе образования специфических для этой группы растений водопроводящих трахеальных элементов: трахеид и производных от них члеников сосудов, что и отражено в названии группы. Гаметофиты сосудистых растений находятся на талломном уровне организации или сильно редуцированы. Тело гаметофитов, как правило, сложено эпидермой и гомогенной паренхимой. Почти никогда не бывает устьиц. Антеридии сосудистых растений чаще всего сидячие, а архегонии погружены брюшком в ткань гаметофита. Сильно редуцированные гаметофиты не образуют гаметангиев. Сперматозоиды сосудистых растений в большинстве случаев имеют многочисленные располагающиеся по спирали жгутики. Почти все современные сосудистые растения гомойогидридны,
и только у немногих их представителей спорофиты вторично пойкилогидридны. Гаметофиты, по-видимому, всегда гомойогидридны, хотя и не обладают столь же эффективными механизмами поддержания водного баланса, как спорофиты тех же видов.
Protracheophyta — полностью вымершая группа самых примитивных сосудистых растений, история которых прослеживается с верхнего силура. Уже в нижнем девоне представители данной группы были достаточно разнообразны и встречались в постоянно влажных биотопах по всей Земле, но к концу верхнего девона вымерли. Это были низкорослые растения, тело которых состояло из разветвленных до 2—5 порядков, как правило, однотипных теломов. Лишь у немногих форм (Taeniocnida, Aglaорћўlon) теломы были дифференцированы на спороносные и вегетативные. Ветвление теломов обычно описывают как апикальное дихотомическое, варьировавшее от изотомии у более примитивных форм до резкой анизотомии у наиболее продвинутых. В центре теломов располагался центральный тяж проводящих элементов, который обычно описывают как гаплостелу. Большинство Protracheophyta были гелофитами или гигрофитами, населявшими болотистые места. Некоторые виды были гидатофитами, скорее всего, вторично вселившимися вводную среду.












23.
Отдел ПЛАУНОВИДНЫЕ. Класс ЗОСТЕРОФИЛЛОВЫЕ. Особенности строения, эволюционное значение. Класс ПЛАУНОВЫЕ. Особенности строения, биологии и цикла развития равноспоровых плауновидных. Равноспоровые представители плауновидных в составе флоры
региона. Краткая характеристика порядков и их представителей. Эволюционное значение порядка ДРЕПАНОФИКУСОВЫЕ.
Отдел Плауновидные. Время возникновения – нижний девон. Расцвет в карбоне, в перми гигантские Лекоподиофиты начинают вымирать. Современные представители – около 1000-1200 видов. Включает талломные и побеговые (преобладают) формы. Большинство последних – первично гоморизные растения с придаточными корнями побегового или ризофорного происхождения. Листья микрофильные, энационного происхождения. Спорангии короткие, вскрывающиесы наверху двумя створками. Тип стелы – прото-, актино- или плектостела. Протоксилема всегда экзархная или почти экзархная. Нет камбия, либо очень слабо развит. Экология: От гигрофитов (селягинеллы) и гидрофитов (полушники) до мезофитов и ксеромезофитов (плауны, баранцы). Распространение: тропики, субтропики, реже – в умеренной зоне, многие – обитатели высокогорных и приморских областей.
Очень редко они поднимают свои побеги, не имеющие камбия, выше чем на один метр. С помощью стелющихся побегов плауновидные хорошо и быстро вегетативно размножаются.
Побеги большинства растений этой группы покрыты спирально расположенными мелкими листьями – микрофиллами. Характерно для плауновидных также дихотомическое или вильчатое ветвление побегов. Подземные части этих растений обычно представлены корневищем с придаточными корнями и видоизмененными листьями. У некоторых видов подземные части побегов образуют своеобразный орган, несущий расположенные по спирали корни и называемый ризофором. Надземные и подземные побеги плауновидных нарастают ограниченное время, поскольку со временем клетки верхушечных меристем теряют способность к делению.
ЖЦ, размножение.Спороносные листочки (спорофиллы) плауновидных по форме, размерам и цвету могут быть похожи на вегетативные листья (трофофиллы) или в той или иной степени отличаться от них. Чередуясь со стерильными листьями, спорофиллы образуют на протяжении стебля спороносные зоны или собраны в расположенные на концах ветвей компактные образования – стробилы.
Среди плауновидных есть равно- и разноспоровые растения, гаметофиты (заростки) которых, соответственно, либо обоеполые (т.е. на них образуются как мужские, так и женские половые клетки), либо однополые. Есть между ними и другие отличия. Заростки современных равноспоровых (представителей класса плауновых) созревают в течение 1–15 лет, достигая в длину от 2 до 20 мм. При этом они развиваются в подземных или полуподземных условиях, и питаются в значительной степени или исключительно за счет образования союза с почвенными грибами. Заростки разноспоровых плауновидных (к ним относятся все представители класса полушниковых) развиваются обычно в течение нескольких недель. Они вообще лишены хлорофилла и живут за счет питательных веществ, содержащихся в споре. Поэтому они очень мелкие и даже в зрелом виде лишь слегка выступают за пределы оболочки споры.
Половые органы, развивающиеся на заростках плауновидных, представлены антеридиями, в которых развиваются сперматозоиды с двумя или многими жгутиками, и архегониями, в которых развиваются яйцеклетки. Для того чтобы сперматозоид мог достичь яйцеклетки, необходимо наличие капельно-жидкой воды. Оплодотворенная яйцеклетка у плауновидных сразу же, не впадая в состояние покоя, развивается в растение нового бесполого поколения – спорофит.
Спорангии — органы спороношения — образуются в пазухах листьев или на их внутренних поверхностях. Споры могут созревать довольно долго, высыпаясь, они прорастают и дают начало гаметофитам только на влажных почвах.

Цикл развития плауна булавовидного: А — спорофит; Б — гаметофит; 1 — ползучий побег с придаточными корнями; 2 — восходящие побеги; 3 — ножка спороносных колосков; 4 — листочки: восходящего побега (а) и ножки спороносныхколосков (б); 5 — спороносные колоски; 6 — споролистики: вид с брюшной (в) и спинной (г) сторон; 7 — спорангии; 8 — споры; 9 — прорастающая спора; 10 — архегоний; 11 — антеридий; 12 — оплодотворение; 13 — оплодотворенная яйцеклетка; 14 — развитие нового спорофита на гаметофите
Плауновидные, являясь обязательными компонентами многих экосистем, нуждаются в охране и защите.
В волгоградской области – Плаун Булавовидный (не точно).
Класс Зостерофилловые. Древние вымершие примитивные плауновидные (7-8 родов). Сущ.в девоне, в начвле перми вымирают. Имели теломную организацию тела (плагиотропные разветвленные ризомоиды и ортотропные слабо разветвленные или неветвящиеся спороносные талломы 20-30 см выс.(система синтеломов – стеблей нет). Телом и ризомоиды могли быть уплощены. Имелись эмергенцы – вероятные будущие зачатки энациев. И в ризомоидах и в теломах – протостела (гаплостела). Спорангии латеральные, вскрывались двумя створками верхушечной щелью, располагались или рассеянно или скучанно, формирую терминальные стробилы. Достоверные гаметофиты известны. Экология: Гидрофиты, гелофиты, гигрофиты.
Зостерофилловые считаются родственниками плауновидных, в их строении присутствуют несколько важных общих признаков: экзархная протостела и боковые почковидные спорангии. В отличие от плаунов, зостерофилловые не имели листьев, вместо них у этих растений имелись различного типа боковые отростки стебля, по-видимому, служившие для увеличения фотосинтезирующей поверхности.Зостерофилловые занимали промежуточное положение между риниофитами и плауновидными. По мнению некоторых учёных, эти растения являлись предками плауновидных, другие же палеоботаники считают её парафилетичной группой либо сестринской группой по отношению к ним.
Класс Плауновые.Известны с нижнего девона, существуют до настоящего времени. Главного корня нет, имеются лишь придаточные корни. Ветвление побегов дихотомическое. Листья (микрофиллы) энационного происхождения, без лигул. И в стебле, и в корне протостела (актино- и плектостела) с экзархной или мезархной протоксилемой. Нет камбия. Трахеиды с окаймленными порами (более совершенные). Спорангии с колонками внутри (более совершенные). Появляются спорофиллы с одночными спорангиями на адаксиальной (верхней) поверхности. Равноспоровые растения. Гаметофиты небольшие по размерам и известны только у современных представителей. Систематика: Класс Lycopodiopsida: Порядок Дрепанофикусовые (Drepanophycales) - Вымершая группа примитивных равноспоровых плауновых. Порядок Плауновые (Lycopodiales) – современные равноспоровые плауновые.
Порядок Дрепанофикусовые. Вымершие примитивные плауновые (3 рода). Жили вместе и параллельно с Zosterophullopsida в девоне. Невысокие (20-40 см, 50-60 см) с дихотомически ветвящимися плагиоортотропными или ортотропными побегами и плагиотропными ризомоидами. Корни придаточные (были не у всех), ризоидов не было, симбиотрофная микориза. В ризоидах – гаплостела, в стеблях – актиностела. Камбия нет, на стебле – настоящие устьица. Хорошо выражена трехслойная кора (кортекс), наружный слой (экзодерма) – хлоренхима. Листья (филлоиды) – васкуляризированные эмергенцы, переходящие а энации. Спорангии располагались поочередно в верхней части стеблей по одному в пазухах спорофиллов на их адаксиальной поверхности. Спорофиллы ничем не отличались от трофиллов. Экология: произрастали на берегах водоемов и болот. Были галофиты (т.к. присутствовала аэренхима). Drepanophycales, вероятно, являются прямыми предками современных Lycopodiales.
Порядок Плауновые. Ныне живущие равноспоровые плауновые (7 родов, около 200 видов, 18 – в России). Многолетнее травянистое растение с дихотомическими и дихоподиально нарастающими плагиотропными или ортотропными побегами. Корни придаточные, первично гоморизные, апицегенные, не гомологичны корням папоротникообразных и семенных растений. Регенерации у таких корней нет, в тонких корнях – актиностела, в толстых – плектостела. Корни плауновых возникли в ходе эволюции независимо от корней папоротникообразных и семенных растений. Филлоиды (листоподобные органы) – энационного происхождения. В стебле – актино- и плектостела с экзархной протоксилемой. Спорангии – на спорофиллах, реже пазушные. Гаметофиты обоеполые, подземные или полуподземные микотрофные, реже надземные (тогда автотрофные), иногда полу- и галопаразиты. Экология: Наземные ползучие или эпифитные травы, обитатели умеренных (до тундр и высокогорий) широт и тропич.дожд.лесов.
24.
Отдел ПЛАУНОВИДНЫЕ. Сборная группа лигульных плауновидных. Порядок ПРОТОЛЕПИДОДЕНДРОВЫЕ. Особенности строение, время существование. Класс СЕЛЯГИНЕЛЛОВЫЕ. Особенности строения, биологии и цикла развития разноспоровых плауновидных. Краткая характеристика их важнейших представителей.
Отдел ПЛАУНОВИДНЫЕ– см предыдущий вопрос.
Нетаксономическая группа лигульных плауновидных включает преимущественно разноспоровые плауновые, которые обладают особой анатомо-морфологической структурой – лигулой. Лигула расположена на адаксиальной стороне микрофильного листа. Она выходит из основания ямки. От проводящего пучка жилки листа к ней подходит тяж трахеид. Функции лигулы до конца не ясны. Предполагается, что она может всасывать, или, наоборот, выделять воду. У современных представителей она утратила свое значение и сохраняется как атавизм.
У многих представителей этой группы имеются особые вегетативные органы ризофоры, которые возникли, по-видимому, независимо в разных таксонах.
Порядок ПРОТОЛЕПИДОДЕНДРОВЫЕ. Полностью вымершая группа. Существовала в нижнем девоне – самом начале нижнего карбона. Изучена недостаточно, их происхождение и систематика неясны.Небольшие в стелющиеся растения с полегающими побегами. Корни у пролепидодендровых не известны. Стебли с актиностелой (экзархная и мезархная протоксилема).Листья не энационного, а теломного происхождения. Равноспоровые растения с уникальными среди Lycopodiophyta располождением спорангиев – ориентированы вдоль листа и вскрывались продольной щелью. Гаметофиты неизвестны.
Класс Селягинелловые. Ныне живущие и ископаемые формы. Род Selaginella (700 видов) известен с перми. Вымершие жили в карбоне и мезозое. Напочвенные, эпифитные и эпилитные травы с плагио-, реже ортотропными побегами 15-20 см (лианы – от 1,5 до 20 м). Побеги билатеральные, с четырехрядными листьями («спинные» и «боковые»). Стела стебля (гапло-, плекто- или полистела) – внутри воздухоносной полости на особых тяжах (трабекулярных нитях). Лигула – в пазухе листа, в небольшой ямке. Ризофоры (безлистные осевые органы) закладываются в развилках стебля экзогенно; от них отходят эндогенные апицигенные придаточные корни; анатомия корня своеобразны (монархный пучок). Разноспоровые растения. Обитатели тропиков и субтропиков, но есть и высокогорные и приполярные растения. Подавляющее большинство – гигрофиты, обитатели влажных дождевых тропических лесов; в меньшей степени – мезофиты. Иногда встречаются оригинальные пойкилогидрические ксерофиты.
ЖЦ Селагинелла плауновидная Selaginella selaginoides. Побеги ползучие, ветвистые, стебель округлый. Листочки тоненькие, отогнутые назад, удлиненно-яйцевидные, заостренные, зубчатые. На их верхней стороне у основания имеется язычок. Спороносные колоски, или стробилы, верхушечные, одиночные, овально-цилиндрические, длиной около 3 см. Споролистики длиннее вегетативных листочков (около 5 мм), с многочисленными длинными зубчиками и язычками. В верхней части колоска расположены мелкие микроспорофиллы с микроспорангием у основания. Они содержат множество микроспор, из которых внутри развиваются мужскиемикрогаметофиты, или заростки, со сперматозоидами. В нижней части колоска находятся мегаспорофиллы с макро-или мегаспорангием. Они содержат по 4 крупных мегаспоры, в которых прорастают женские мегагаметофиты, или заростки. Таким образом, заростки двудомные. На женском заростке имеется небольшая, выдвинутая наружу лопасть, на которой находятся архегонии с яйцеклеткой. После оплодотворения из зиготы развивается зародыш нового спорофита.
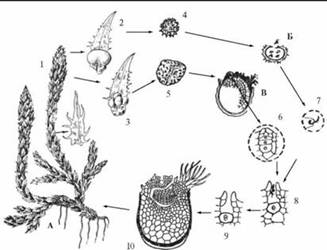
Цикл развития разноспоровых плаунов на примере селагинеллы плауновидной: А — спорофит; Б — мужской заросток — микрогаметофит; В — женский заросток — мегагаметофит; 1 — стробил; 2 — микроспорофилл с микроспорангием; 3 — мегаспо-рофилл с мегаспорангием; 4 — микроспора; 5 — мегаспора; 6 — архегоний; 7 — спермий; 8 — оплодотворение; 9 — зигота; 10 — развитие нового спорофита
25.






26.



27






28.




29.



30.







30
Подотдел Папоротники - Polypodidae
Общая характеристика.
Ископаемые и современные растения.
Особенности жизненного цикла:чередование поколений с доминированием бесполого поколения (спорофита).Жизненные формы: древовидные (ископаемые и современные) и многолетние травянистые растения размерами от 5 мм до 25 м. Побеги дифференцированы на стебли и вайи, имеются придаточные корни. Примитивные имеют теломную организацию.
Стебли: у травянистых форм обычно укороченные, горизонтальные, проводящая система в виде диктиостелы или сифоностелы, у вымерших и в примитивных современных семействах (Схизейных, Гименофилловых, Глейхениевых) – протостелы. Ксилема состоит из трахеид.
Листовые следы двух-, многопучковые. Над местом отхождения листового следа в стеле образуется листовая лакуна.
Листья теломной природы, крупные, обычно перисто рассеченные, они у папоротников называются вайями. В молодом состоянии нередко улиткообразно свернуты. Взрослые – перистые (от однажды- до многократно перистых), реже цельные или пальчатые. Вайи папоротников часто совмещают функции фотосинтеза и спороношения, но у многих (страусник, оноклея) они дифференцированы на стерильные (фотосинтезирующие) и фертильные (спороносные). В этом случае спорофиллы хорошо отличаются по форме и цвету от трофофиллов.

а – черешок;
б – пластинка вайи;
в – перо первого порядка;
г – перышки (или перья второго порядка);
д – рахис.
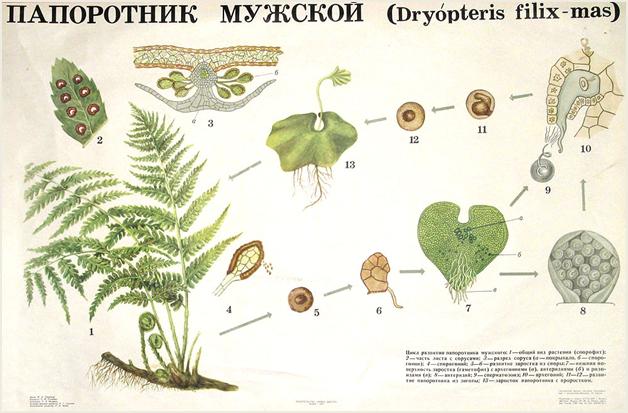
Спорангии у большинства современных папоротников мелкие, с небольшим (часто 64) числом спор. Но у ужовниковых и мараттиевых они крупные и содержат до 15000 спор. Обычно снабжены механизмом для вскрывания и рассеивания спор. У некоторых мараттиевых и ужовниковых они срастаются в синангии.
У большинства папоротников спорангии тесно сближены в группы (образуют так называемые сорусы). Сорусы могут быть открытыми (например, у голокучника), или закрыты выростами эпидермальной ткани – покрывальцами (так называемыми индузиями).
Споры морфологически одинаковые (равноспоровость), иногда дифференцированы на микро- и мегаспоры (разноспоровость) – у сальвиниевых, азолловых и марсилеевых.
Гаметофит: из равных спор вырастает гаплоидный обоеполый гаметофит. Он представляет собой тонкие зеленые недолговечные пластиночки до 5 мм шир., с архегониями и антеридиями обычно на нижней стороне.
У тропических папоротников гаметофит долгоживущий, лентовидный. Он размножается вегетативно – выводковыми почками (геммами). Бывает нитчатый гаметофит, похожий на водоросли. У некоторых видов он цилиндрический, бесхлорофильный, подземный и микоризный. У разноспоровых папоротников гаметофиты раздельнополые и сильно редуцированные.
Хозяйственное значение.
Молодые листья некоторых видов (например, орляка) и сердцевину древовидных папоротников употребляют в пищу.
Многие папоротники выращивают в комнатах и оранжереях как декоративные, например, асплениум, адиантум и др.
Экстракты из щитовника обладают антигельминтным действием и используются в медицине.
Водные папоротники рода Азолла в тропической Азии служат азотным удобрением рисовых полей.
Дата добавления: 2018-02-28; просмотров: 4974; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!