Оценка процесса β-окисления с энергетич. точки зрения.
-Если окисляется 1 М миристиновой(С14) к-ты (14:0),то из неё получается 7 М(14/2) ацетил-Ко-А,кот-ые распадаясь в цитратном ц. Кребса, дают ~70 М АТФ (7*10)
- При д-ии ацетил-Ко-А-дегидрогеназы и β-гидроксиацил-Ко-А-дегидрогеназы образ по 6 М ФАДН2(14/2-1) и НАДН+,кот-ые дают соответ-но 9 (6*1,5) М АТФ и 15 (6* 2,5) М АТФ.
 Всего: 70+9+15=94 М АТФ(надо вычесть затрач-ые на активацию ЖК 2 М АТФ(р-ия АТФ АМФ),в итоге получ. энергетич. рез-т 92 М АТФ.
Всего: 70+9+15=94 М АТФ(надо вычесть затрач-ые на активацию ЖК 2 М АТФ(р-ия АТФ АМФ),в итоге получ. энергетич. рез-т 92 М АТФ.
· Дать определение понятию липиды, объяснить, указать их свойства, функциональное значение для организма. Привести классификацию липидов, дать общие формулы представителей классификации.
Липиды(от греч. lípos — жир), жироподобные вещества, входящие в состав всех живых клеток и играющие важную роль в жизненных процессах. Будучи одним из основных компонентов биологических мембран, Л. влияют на проницаемость клеток и активность многих ферментов, участвуют в передаче нервного импульса, в мышечном сокращении, создании межклеточных контактов, в иммунохимических процессах. Др. функции Л. — образование энергетического резерва и создание защитных водоотталкивающих и термоизоляционных покровов у животных и растений, а также защита различных органов от механических воздействий.
· КЛАССИФИКАЦИЯ ЛИПИДОВ
Существует несколько классификаций липидов. Наибольшее распространение получила классификация, основанная на структурных особенностях липидов. По этой классификации различают следующие основные классы липидов.A. Простые липиды: сложные эфиры жирных кислот с различными спиртами.1. Глицериды (ацилглицерины, или ацилглицеролы – по международной номенклатуре) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот.2. Воска: сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов.Б. Сложные липиды: сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы.1. Фосфолипиды: липиды, содержащие, помимо жирных кислот и спирта, остаток фосфорной кислоты. В их состав часто входят азотистые основания и другие компоненты:а) глицерофосфолипиды (в роли спирта выступает глицерол);б) сфинголипиды (в роли спирта – сфингозин).2. Гликолипиды (гликосфинголипиды).3. Стероиды.4. Другие сложные липиды: сульфолипиды, аминолипиды. К этому классу можно отнести и липопротеины.B. Предшественники и производные липидов: жирные кислоты, глице-рол, стеролы и прочие спирты (помимо глицерола и стеролов), альдегиды жирных кислот, углеводороды, жирорастворимые витамины и гормоны.
· Синтез высших жирных кислот. Привести последовательность реакций с формулами,названиями ферментов,участием АТФ, НАДФ, указать локализацию процесса и его значение.
Образование малонил-КоА. Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца. Катализирует эту реакцию фермент ацетил-КоА-кар-боксилаза. Фермент содержит в качестве простетической группы биотин. Авидин – ингибитор биотина угнетает эту реакцию, как и синтез жирных кислот в целом.
Установлено, что ацетил-КоА-карбоксилаза состоит из переменного числа одинаковых субъединиц, каждая из которых содержит биотин, биотинкарбоксилазу, карбоксибиотинпереносящий белок, транскарбоксилазу, а также регуляторный ал-лостерический центр, т.е. представляет собой полиферментный комплекс.
Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА: 
Малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментной системы малонил-КоА быстро превращается в жирные кислоты.
Энзиматические системы, осуществляющие синтез жирных кислот, называются жирно-кислотными синтетазами. Они широко встречаются в природе и могут быть изолированы из различных одноклеточных организмов, растений и животных тканей.
Жирно-кислотные синтетазы делятся на 2 группы. К первой группе относятся полиэнзимные, не поддающиеся фракционированию комплексы с мол. м. порядка 500000, в которых все индивидуальные энзимы собраны в компактную структуру. В частности, в эту группу входят жирно-кислотные синтетазы животных тканей и дрожжей.
Вторая группа включает жирно-кислотные синтетазы, из которых отдельные энзимы могут быть выделены методами белкового фракционирования. Такие синтетазы встречаются у ряда микроорганизмов (в частности, у E.coli) и растений. Иными словами, в этих случаях все индивидуальные ферменты синтетазной системы находятся в виде автономных полипептидов.
Мультиферментный комплекс, называемый синтетазой (синтазой) жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Этот белок относительно термостабилен, имеет две свободные HS-группы (цистеина и фосфопантетеинового остатка, присоединенного к ОН-группе серина) и вовлекается в процесс синтеза высших жирных кислот практически на всех его этапах. Мол. масса АПБ составляет около 10000. Данный белок в синтетазной системе выполняет роль КоА. Заметим, что в животных тканях не удалось обнаружить свободного АПБ, подобного микробному. Из печени выделен полиэнзимный комплекс, содержащий все энзимы, необходимые для синтеза жирных кислот. Энзимы комплекса настолько прочно связаны друг с другом, что все попытки изолировать их в индивидуальном виде не увенчались успехом. Приводим последовательность реакций, происходящих при синтезе жирных кислот:

Далее цикл реакций повторяется. Допустим, что идет синтез пальмитиновой кислоты (С16). В этом случае образованием бутирил-АПБ завершается лишь первый из 7 циклов, в каждом из которых началом является присоединение молекулы малонил-АПБ к карбоксильному концу растущей цепи жирной кислоты. При этом отщепляется дистальная карбоксильная группа малонил-АПБ в виде СО2. Например, образовавшийся в первом цикле бутирил-АПБ взаимодействует с малонил-АПБ:

Завершается синтез жирной кислоты отщеплением HS-АПБ от ацил-АПБ под влиянием фермента деацилазы. Например:

Суммарное уравнение синтеза пальмитиновой кислоты можно записать так:

Или, учитывая, что на образование одной молекулы малонил-КоА из ацетил-КоА расходуются одна молекула АТФ и одна молекула СО2, которая затем отщепляется, суммарное уравнение можно представить в следующем виде:

·  Основные этапы биосинтеза жирных кислот можно представить в виде схемы:
Основные этапы биосинтеза жирных кислот можно представить в виде схемы:
· Привести классификацию липопротеинов плазмы крови и расказать об их строении, какое они имеют значение для организма? Какие заболевания связаны с ними?
Плазменные липопротеины (ЛП) – это сложные комплексные соединения, имеющие характерное строение: внутри липопротеиновой частицы находится жировая капля (ядро), содержащая неполярные липиды (три-глицериды, эстерифицированный холестерин); жировая капля окружена оболочкой, в состав которой входят фосфолипиды, белок и свободный холестерин. Толщина наружной оболочки липопротеиновой частицы (ЛП-частица) составляет 2,1–2,2 нм, что соответствует половине толщины ли-пидного бислоя клеточных мембран. Это позволило сделать заключение, что в плазменных липопротеинах наружная оболочка в отличие от клеточных мембран содержит липидный монослой. Фосфолипиды, а также неэсте-рифицированный холестерин (НЭХС) расположены в наружной оболочке таким образом, что полярные группы фиксированы наружу, а гидрофобные жирно-кислотные «хвосты» – внутрь частицы, причем какая-то часть этих «хвостов» даже погружена в липидное ядро. По всей вероятности, наружная оболочка липопротеинов представляет собой не гомогенный слой, а мозаичную поверхность с выступающими участками белка. Существует много различных схем строения ЛП-частицы. Предполагают, что входящие в ее состав белки занимают только часть наружной оболочки. Допускается, что часть белковой молекулы погружена в ЛП-частицу глубже, чем толщина ее наружной оболочки (рис. 17.4). Итак, плазменные ЛП представляют собой сложные надмолекулярные комплексы, в которых химические связи между компонентами комплекса носят нековалентный характер. Поэтому применительно к ним вместо слова «молекула» употребляют выражение «частица». Аполипопротеины (апобелки, апо) входят в состав липопротеинов. Это один белок либо несколько белков, или полипептидов, которые называют апобелками (сокращенно апо). Апобелки выполняют не только структурную функцию, но и обеспечивают активное участие комплексов ЛП в транспорте липидов в токе крови от мест их синтеза к клеткам периферических тканей, а также обратный транспорт холестерина в печень для дальнейших метаболических превращений. Апобелки выполняют функцию лигандов во взаимодействии ЛП со специфическими рецепторами на клеточных мембранах, регулируя тем самым гомеостаз холестерина в клетках и в организме в целом. Не меньшее значение имеет также регуляция апобелками активности ряда основных ферментов липидного обмена: лецитин-холестеролацилтрансферазы, липопротеинлипазы, печеночной триглицеридлипазы. Структура и концентрация в плазме крови каждого апобелка находится под генетическим контролем, в то время как содержание липидов в большей степени подвержено влиянию диетических и других факторов. Дислипопротеинемией (ДЛП) называют изменения в содержании липо-протеинов в плазме (сыворотке) крови: повышение, снижение или практически полное отсутствие. Сюда же относят случаи появления в крови необычных или патологических ЛП.
Окисление насыщенных и ненасыщенных жирных кислот. Привести последовательность реакций с формулами,названиями ферментов, участием АТФ,НАД, указать локализацию процесса и его значение.Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg2+. Реакция катализируется ферментом ацил-КоА-синтетазой 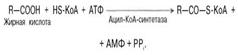 В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Считают, что активация жирной кислоты протекает в 2 этапа. Сначала жирная кислота реагирует с АТФ с образованием ациладенилата, представляющим собой эфир жирной кислоты и АМФ. Далее сульфгидрильная группа КоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-КоА и АМФ.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление. Переносчиком активированных жирных кислот
с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин. Ацильная группа переносится с атома серы КоА на гидро-ксильную группу карнитина с образованием ацилкарнитина, который диффундирует через внутреннюю митохондриальную мембрану:

Реакция протекает при участии специфического цитоплазматического фермента карнитин-ацилтрансферазы. Уже на той стороне мембраны, которая обращена к матриксу, ацильная группа переносится обратно на КоА, что термодинамически выгодно, поскольку О-ацильная связь в кар-нитине обладает высоким потенциалом переноса группы. Иными словами, после прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацилтрансферазы:
 Внутримитохондриальное окисление жирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Внутримитохондриальное окисление жирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты. Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-де-гидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:

Существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):

Заметим, что гидратация еноил-КоА стереоспецифична, подобно гидратации фумарата и аконитата (см. с. 348). В результате гидратации транс-Δ2-двойной связи образуется только L-изомер 3-гидроксиацил-КоА.
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакцию катализируют НАД+-зависимые дегидрогеназы:
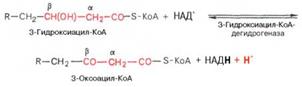
Тиолазная реакция. В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-ке-тотиолазой):

Баланс энергии. При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5 х 7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле трикарбоновых кислот, дает 12 молекул АТФ, а 8 молекул ацетил-КоА дадут 12 х 8 = 96 молекул АТФ.
Таким образом, всего при полном β-окислении пальмитиновой кислоты образуется 35 + 96 = 131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131 – 1 = 130 молекул АТФ. Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь АТФ характеризуется величиной 7,6 ккал/моль. Нетрудно подсчитать, что примерно 990 ккал (7,6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТФ, а оставшаяся часть, очевидно, теряется в виде тепла.
· Что из себя представляют фосфолипиды, привести формулы, указать их местонахождение и значение. Какие азотистые основания встречаются в составе фосфолипидов/формулы/? Как и где осуществляется их переваривание?
Фосфолипиды являются производными фофотидной кислоты в их состав входит глицерин, жк, фосфорная кислота и обычно азотосодержащие соединения, они являются важнейшим компонентом клеточных мембран. Для фосфолипидов характерно что одна часть их молекулы гидрофобная, а другая гирофильна благодаря отрицательному заряду остатка фосфорной кислоты и положительного заряда радикала, они обладают выраженными полярными свойствами.
CH2—O—CR1=O CH2—O—CR1=O ….O─CH2─CH2─NH3
CH —O—CR2=O CH —O—CR2=O Фосфотидилэтаноламин
CH2—O─POH=O CH2—O─PO-=O
OH O─CH2─CH2─N(CH3)3
Фосфотидная кислота Фосфотидилхолин
В фосфоглициридах одна из трех гидроксильных групп глицерина св-на с фосфорной кислотой а ФК соединена эфирной св-ю с азотистым основанием(холин,этаноламин,серин)
· Переваривание фосфоглицеридов
В кишечнике подвергаются воздействию специфических гидролитических ферментов, (фосфолипаз) разрушающих эфирные связи между компонентами, в фосфолипидах. Фосфолипиды выделяются с панкреатическим соком, фосфоглицериды расщепляются в кишечнике с образованием глцерина, высших ЖК азотистого основания и фосфорной кислоты. Фосфолипаза, поджелудочная железа поступает в полость тонкого кишечника неактивной форме и только после воздействия трипсина приобретает активность.
· Синтез триглицеридов. Привести последовательность реакций с формулами, названиями ферментов, участием АТФ, НАДФ, указать локализацию процесса и его значение.
Дата добавления: 2015-12-19; просмотров: 1; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!