Электропроводность электролитов
Электролитическая диссоциация количественно описывается степенью диссоциации – отношением концентрации распавшихся молекул C к их исходной концентрации: a = С / С0. Основную роль в прохождении электрического тока играют низкомолекулярные ионы Na, K, Cl, H, OH, Ca.
Если к раствору электролита с помощью электродов приложить разность потенциалов от внешнего источника напряжения, ионы будут испытывать воздействие кулоновской силы FК = qE, где q величина заряда иона, Е – напряженность электрического поля Е = U/ l (U - приложенное напряжение, l - расстояние между электродами). Под действием этой силы наблюдается упорядоченное движение электрических зарядов: положительные ионы перемещаются в направлении катода (отрицательного электрода – см. рис. 9-1), отрицательно заряженные – к аноду. Поэтому соответствующие заряды называют катионами и анионами. В процессе движения ионы испытывают силу сопротивления движению, которая количественно может быть описана формулой Стокса:
FС = 6 p hr W.
Здесь обозначено h - вязкость среды, r – радиус движущейся частицы, W - скорость ее движения.
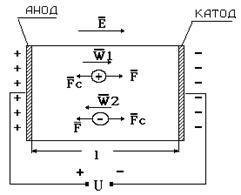
Рис. 9-1
Из этой формулы следует, что по мере увеличения скорости под действием кулоновской силы увеличивается и сопротивление среды. Когда эти силы станут равными FК = FC, ион будет перемещаться равномерно со скоростью W = qE/6phr. Упорядоченное движение ионов создает электрический ток, сила которого: i = U/R. Величину сопротивления можно выразить R = r l / S, где r – удельное электрическое сопротивление, l – длина и S – площадь поперечного сечения проводника. Используя введенные обозначения, силу тока можно представить i = U·S / r l. Отношение силы тока к площади сечения проводника называется плотностью тока j = i / S; величина, обратная удельному электрическому сопротивлению, называется удельной электропроводностью s = 1/r, U/ l = E. Здесь отметим, что величина обратная электрическому сопротивлению l= 1/R является электропроводностью, а омическое сопротивление связано с удельной электропроводностью соотношением R = l / sS. C учетом введенных обозначений получаем закон Ома, справедливый для любого участка проводника j = s ´ E – плотность электрического тока пропорциональна напряженности электрического поля Е (коэффициентом пропорциональности служит удельная электропроводность s проводника). Выясним, от каких факторов зависит величина s для электролитов.
Сила электрического тока определяется количеством заряда прошедшего через поперечное сечение проводника в единицу времени: i = Q / t, и, следовательно, плотность тока j = Q/St. Величину заряда Q можно выразить через заряд одного иона q и количество зарядов N, прошедших через поперечное сечение: Q = q ´ N. Тогда плотность тока будет равна: j = q ´N / S´t. Домножим и разделим последнее равенство на длину проводника l: j=(qN l) / (tS l). Произведение площади сечения S на длину l представляет объем проводника V = l S. Отношение числа ионов N к объему является концентрацией ионов С = N / V = N / S l, которая зависит от степени диссоциации a = C / C0. Поэтому (N/S)· l = N / V = C = a C0. Соотношение l / t = W является скоростью движения иона. Подставляя все использованные обозначения в формулу для плотности тока, получаем:
j = q a C0 W.
Как было показано выше, скорость движения ионов W = qE / 6phr и поэтому:
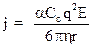
Необходимо учесть, что электрический ток создается в результате упорядоченного движения как положительных, так и отрицательных зарядов. Поэтому результирующая плотность тока:
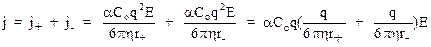 Выражение
Выражение 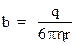 называется подвижностью ионов, имеющих радиусы для положительных r+ и r- для отрицательных ионов. С учетом введенного обозначения плотность тока будет равна:
называется подвижностью ионов, имеющих радиусы для положительных r+ и r- для отрицательных ионов. С учетом введенного обозначения плотность тока будет равна:
j = a C0 q (b+ + b-) E
Сопоставление полученного выражения с формулой закона Ома j = sE позволяет сделать вывод, что величина электропроводности электролита s выражается соотношением: s = a C0 ´q ´ (b+ + b-),
а электрическое сопротивление растворов, содержащих ионы:
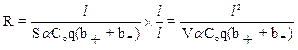
Из последнего равенства следует вывод, что сопротивление растворов электролитов в организме (плазмы крови, мочи, тканевой и спинномозговой жидкости, пота и др.) зависит от концентрации (a C0), заряда (q), подвижности (b) ионов и геометрических размеров проводника. Здесь отметим, что электрическое сопротивление обратно пропорционально объему электролита V. Среда оказывает слабое влияние на движение электрических зарядов, поскольку вязкость воды относительно невелика и низкомолекулярные ионы, за счет которых в электролитах создается электрический ток, испытывают небольшое сопротивление. Иначе дело обстоит в тканях, состоящих из клеток. Для того чтобы описать электрические свойства тканей необходимо рассмотреть закономерности протекания электрического тока и те факторы, которые влияют на его величину. Пусть на гипотетическую клетку действует электрическое поле Е в растворе электролита (рис. 9-2).
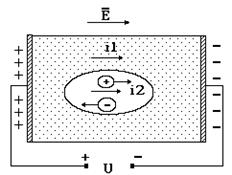
Рис. 9-2
Это поле создано разностью потенциалов U от внешнего источника напряжения. Внутри клетки (в цитоплазме) и в окружающей среде (которая, в данном случае, является аналогом тканевой жидкости) находятся свободные электрические заряды (ионы). Под действием кулоновской силы F = q Е заряды перемещаются в направлении, соответствующему знаку заряда иона.
Упорядоченное движение ионов создает электрические токи в среде i1 и в цитоплазме клетки i2. Результирующий электрический ток в соответствии с законом Ома может быть выражен: i=i1+i2=U/Z, где U – приложенное напряжение, Z – электрический импеданс (суммарное электрическое сопротивление). Импеданс характеризует электрические свойства той среды, в которой протекает электрический ток. Рассмотрим, от каких факторов зависит его величина.
Все без исключения клетки окружены мембранами, состоящими из жироподобных веществ. Поэтому мембраны плохо проводят электрический ток, оказывая сильное сопротивление движению зарядов. Электрическое сопротивление мембраны по сравнению с цитоплазмой и окружающей средой очень велико. Из сказанного становится понятно, что омическое сопротивление мембраны, определяющее движение ионов, существенно больше омического сопротивления цитоплазмы и среды. А если это так, то ионы, движущиеся в цитоплазме под действием внешнего поля Ео и создающие цитоплазматический ток i2, будут тормозиться и накапливаться около мембраны (рис. 9-3).
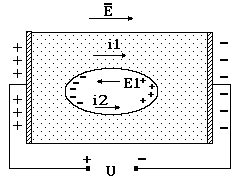
Рис. 9-3
В результате разделения ионов в клетке возникает внутреннее поле Е, направленное противоположно внешнему Е0. Исходя из принципа суперпозиции электрических полей, результирующее поле в цитоплазме Ер = Е0 – Е. Поскольку перераспределение зарядов в клетке осуществляется постепенно, то с течением времени уменьшается суммарное поле Ер и цитоплазматический ток i2. Если измерить результирующий ток i, он изменяется (i ® i1) во времени по зависимости, приведенной на рисунке 9-4.
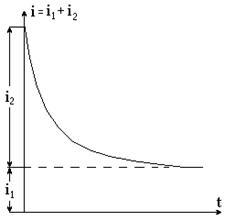
Рис. 9-4
Рассмотренная закономерность движения ионов в клетке и зависимость результирующего тока от времени позволяют сделать вывод, что мембрана клетки ведет себя как конденсатор, в котором происходит разделение зарядов. Перераспределение ионов в клетке, накопление их у мембран, уменьшение внутриклеточного поля, цитоплазматического тока – следствие явления ионной поляризации. Если бы мембраны вообще не пропускали ионы, то с течением времени цитоплазматический ток должен стать i2 = 0. Однако, хоть и плохо мембраны проницаемы для электрических зарядов и поэтому i2 ¹ 0. Из всего вышесказанного можно сделать вывод, что электрические свойства клетки должны быть представлены с помощью омического сопротивления цитоплазмы, мембраны и среды, а также электрической емкости мембраны (рис. 9-5а).
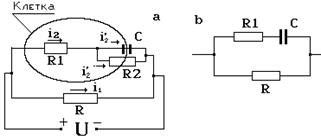
Рис. 9-5
Комбинация из этих элементов, описывающая закономерности протекания электрического тока, называется эквивалентной электрической схемой (см. рис. 9-5а). На этом рисунке приведены электрические элементы, описывающие протекание тока в среде i1 и в цитоплазме i2: R1 – сопротивление цитоплазмы, R2 – мембраны, R – окружающего электролита, С – поляризационная емкость мембраны. Поскольку мембрана плохо пропускает ионы, то ее сопротивление R2 очень велико по сравнению с R1 и R3, и поэтому можно использовать упрощенную эквивалентную схему (рис. 9-5b). В данном случае, когда рассматриваются электрические свойства отдельной клетки, объем раствора электролита V достаточно велик и поэтому мало сопротивление R, которое обратно пропорционально V. Поскольку емкость С и цитоплазматическое сопротивление R1 включены параллельно R, можно считать, что омическое сопротивление раствора, в котором находится одна клетка, омическому сопротивлению электролита. В тканях, состоящих из большого количества клеток, дело обстоит иначе (рис. 9-6). Внешняя среда представляет собой электропроводящую жидкость, объем которой зависит от вида ткани. Например, в крови, на жидкую часть (плазму) приходится до 50% объема всей крови. Поэтому омическое сопротивление крови существенно меньше, чем у других тканей. Наконец, следует отметить, что это сопротивление зависит и от концентрации ионов в тканевой жидкости. К примеру, костная ткань обладает достаточно высоким сопротивлением именно из-за малого количества свободных зарядов.
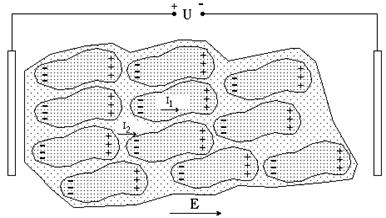
Рис. 9-6
С учетом всего сказанного, величину результирующего электрического сопротивления тканей (импеданс) можно оценить с помощью эквивалентной электрической схемы, приведенной на рисунке 9-7.
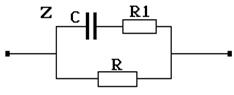
Рис. 9-7
Здесь обозначено: С – поляризационная емкость, R1 – омическое сопротивление цитоплазмы, R – результирующее омическое сопротивление тканевой (внеклеточной) электропроводящей жидкости. Представление электрических свойств ткани с помощью эквивалентной схемы позволяет заключить, что электрический импеданс будет различен на постоянном и переменном электрическом токе. Как известно, электрическое сопротивление емкости Xс = 1 / wC зависит от частоты w переменного электрического тока. Поэтому и импеданс Z должен изменяться в зависимости от частоты приложенного электрического напряжения U. В данном случае с увеличением частоты величина импеданса уменьшается (рис. 9-8), стремясь в пределе к некоторому минимальному значению Zmin.
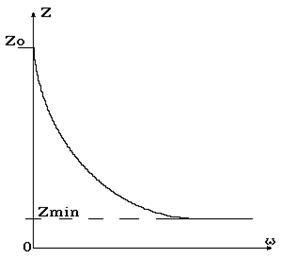
Рис. 9-8
Зависимость суммарного электрического сопротивления тканей от частоты получила название дисперсии электрического импеданса.
Воспользовавшись эквивалентной электрической схемой, можно выяснить физический смысл максимального импеданса Z0 (при w = 0) и Zmin (при w ® ¥). Очевидно, на постоянном токе, когда w = 0 и XС = 1 / wC ® ¥, электрический импеданс определяется сопротивлением тканевой жидкости R, т.е. Z = R. C увеличением частоты, когда ZС = 1 / wC ® 0, Z ® Zmin. Поскольку величина Zmin определяется совокупностью параллельных омических сопротивлений (цитоплазмы и среды), то 1/Zmin = 1/R1 + 1/R2. Обычно для многих тканей, в которых не очень большой объем тканевой жидкости, R >> R1 и поэтому Zmin ~ R1. Если же этот объем велик, как в крови, то Zmin ~ R1R / (R1 + R). На высоких частотах электрический импеданс тканей определяется омическими сопротивлениями клеток и внеклеточной среды.
Из выше рассмотренных теоретических представлений следует вывод: электрические свойства тканей зависят от концентрации ионов в цитоплазме, в тканевой жидкости, от структуры органа или ткани (соотношения суммарного объема клеток и тканевой жидкости) и, наконец, от свойств клеточных мембран. Именно эти зависимости используются для разработки биофизических принципов, способов и устройств, позволяющих диагностировать патологические состояния и процессы в живом организме. Остановимся на некоторых таких методических подходах.
Как уже отмечалось, поляризационная емкость С определяет способность тканей к ионной поляризации – накоплению ионов на клеточных мембранах, и обеспечивает дисперсию электрического импеданса. При повышении проницаемости мембран, при их разрушении вследствие патологических процессов эта способность утрачивается, и в таких тканях ионная поляризация должна проявляться в меньшей степени. Для выявления подобных состояний был предложен коэффициент поляризации тканей, как отношение электрического импеданса на частоте 1 кГц (Z1) к импедансу на частоте 10 кГц (Z 10):
k = Z1/Z10
Поскольку с увеличением частоты электрического тока импеданс уменьшается, Z1 >> Z10 и k > 1.Таким образом, величина k характеризует способность клеточных мембран к ионной поляризации. При разрушении клеток, при гибели тканей, когда уменьшается накопление ионов на мембранах k ® 1 и Z1 ® Z10. К примеру, если для нормальной матки коэффициент поляризации в среднем составляет 2,6, то при развитии опухоли в области изъязвления он составляет всего 1,6. Поэтому величина коэффициента поляризации определяет структурные особенности тканей и функциональные свойства клеточных мембран.
Другим важным примером практического использования электрического импеданса для исследования биологических процессов в организме может служить импедансная реография.
9.2. ИМПЕДАНСНАЯ РЕОГРАФИЯ
Импедансная реография – метод диагностики состояния кровообращения органов и тканей по результатам регистрации электрического импеданса. Как уже отмечалось, объем внеклеточной электропроводящей плазмы в крови достаточно велик, и поэтому результирующее сопротивление Zmin = R1R / (R1 + R) мало по сравнению с сопротивлением любых других тканей. Именно поэтому величина электрического импеданса тканей и органов должна зависеть от объема кровенаполнения – чем больше объем крови, тем меньше должен быть импеданс. Поскольку сердце работает в периодическом режиме, во время систолы ткани и органы наполняются кровью, а в ходе диастолы она оттекает в венозную систему. Следовательно, электрический импеданс должен периодически изменяться, отражая периодическую работу сердца и характер кровенаполнения органов и тканей. Для графической регистрации электрического импеданса тканей используется реограф, принципиальная схема которого приведена на рисунке 9-9.
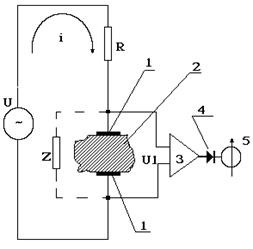
Рис. 9-9
От источника переменного электрического напряжения U электрический ток i через сопротивление R с помощью электродов 1 подводится к изучаемой ткани. Электрические свойства ткани или органа можно представить с помощью электрического импеданса Z.
Для исключения влияния поляризационной емкости на результаты измерения используется относительно высокая частота переменного напряжения f = 100 - 500 кГц. При этом можно считать, что на таких частотах XС=1/wC @ 0, а электрический импеданс Z=Zmin. Падение напряжения на ткани U1 после усиления усилителем 3 и выпрямления детектором 4 регистрируется пишущим прибором 5. Величину напряжения U1 можно определить с помощью закона Ома: U1 = UZ/(Z + R). В реографе сопротивление R выбираются таким образом, чтобы R >> Z и поэтому:
U1 = UZ/(Z + R) ~ UZ/R. Отсюда Z = U1 R/U.
Реограф регистрирует величину обратную электрическому импедансу – электропроводность:
l = 1/Z = U / (UR).
Графическая запись колебаний электрического импеданса во времени называется реограммой (см. рис. 9-10). Из последнего соотношения следует, что во время систолы t1, когда кровь поступает в ткань, импеданс уменьшается и увеличивается электропроводность. В фазу диастолы t2 кровь оттекает и электропроводность уменьшается. Таким образом, t1 + t2 = T представляет собой период работы сердца, а величина ему обратная f=1/T – частоту сердечных сокращений.
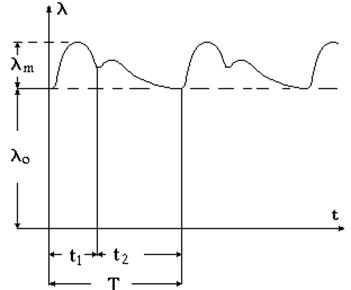
Рис. 9-10
Как свидетельствует проведенный анализ, временные интервалы реограммы характеризуют фазы сердечных сокращений. Однако диагностическая ценность реограмм не ограничивается выявленными показателями.
Теоретическое рассмотрение электропроводящих свойств тканей позволяет заключить, что величина электрического импеданса зависит от объема электролита – чем больше крови поступает в ткань, тем сильнее должна возрастать электропроводность. Поэтому величина lm характеризует объем крови, поступившей в ткань за одно сокращение сердца (пульсовое кровенаполнение).
Анализ реограмм позволяет оценить механические свойства тех сосудов, кровенаполнение которых вызывает колебание электрического импеданса. Если в результате физиологических или патологических процессов уменьшается эластичность (возрастает упругость) сосудов, то поступление крови в ткань затруднено – требуется большее давление для деформации (растяжения) сосудистой стенки. На таких реограммах (рис. 9-11) по сравнению с нормой отмечается уменьшение амплитуды колебаний lm2, падение скорости увеличения электропроводности: наблюдается сглаживание пульсаций. Наоборот, скорость снижения проводимости в период диастолы возрастает, т.к. растянутый сосуд за счет высокой упругости оказывает выраженное давление на оттекающую кровь. При полной закупорке сосудов колебание электропроводности не наблюдается. При повышенной эластичности (уменьшении упругости) сосудов увеличивается амплитуда колебаний, скорость нарастания и уменьшается скорость падения электропроводности.
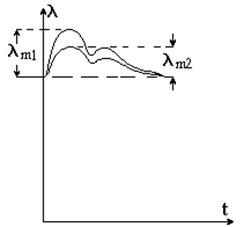
Рис. 9-11
Таким образом, регистрация электрического импеданса (электропроводности) во времени, и последующий биофизический анализ позволяет получить ценную диагностическую информацию о деятельности сердца (определить длительность систолы, диастолы, частоту сердечных сокращений, ударный и минутный объем), а также оценить кровообращение в тканях (их пульсовое кровенаполнение, механические свойства сосудов).
9.3. ТКАНИ ОРГАНИЗМА КАК ДИЭЛЕКТРИКИ
Из физики известно, что к диэлектрикам относятся такие материалы, в которых отсутствуют свободные электрические заряды. Среди биологических тканей, приближающихся по своим электрическим свойствам к диэлектрикам, следует упомянуть жировую ткань, кости, сухожилия, эпидермис. Следует, однако, отметить, что их электропроводящие свойства очень сильно зависят от содержания водных растворов - в сухом состоянии упомянутые биологические объекты - хорошие диэлектрики. На рис. 9-12 приведена зависимость электрического сопротивления кожи R от интенсивности потоотделения Q (Q = M/(St), где М – масса пота в граммах, продуцируемого потовыми железами на 1 кв. м. поверхности кожи в 1 час). Из этого графика видно, что с уменьшением потовыделения сопротивление кожи очень быстро возрастает, достигая при Q < 40 г/кв.м. ´ час десятков МОм.
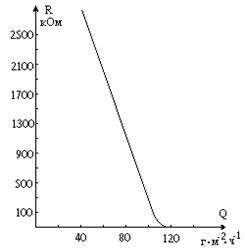
Рис. 9-12
В диэлектриках все электрические заряды находятся в связанном состоянии. От характера этих связей различают полярные и неполярные диэлектрики. В полярных диэлектриках связанные заряды образуют диполи. Не возвращаясь к рассмотренному выше понятию электрического диполя, отметим, что при воздействии внешнего постоянного поля Е заряды диполя испытывают воздействие равных по величине и противоположных по направлению кулоновских сил F=qE (рис. 9-13).
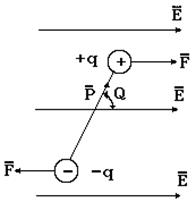
Рис. 9-13
Пара этих сил создает механический момент, который стремится ориентировать диполь таким образом, чтобы вектор электрического момента совпадал по направлению с напряженностью электрического поля Е. Как показывает теория, величина механического момента равна: M = P·E·sinQ, где Q – угол между направлениями векторов электрического момента и напряженности электрического поля. Ориентация диполей в электрическом поле называется дипольной поляризацией. Механическому влиянию электрического поля препятствует тепловое хаотическое движение диполей. В результате ориентации диполей их электрические поля суммируются (рис. 9-14) и, поскольку силовые линии диполей направлены противоположно внешнему полю, напряженность внутреннего поля Е1 направлена против вектора внешнего поля Е0.
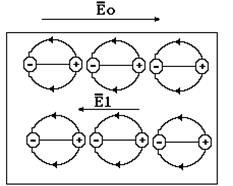
Рис. 9-14
В соответствии с принципом суперпозиции электрическое поле в полярном диэлектрике будет равно: Е = Е0 - Е1. Отношение Е0/Е= e называется относительной диэлектрической проницаемостью материала. Как следует из приведенного соотношения, эта величина характеризует степень ослабления внешнего электрического поля в диэлектрике в результате дипольной поляризации. Если поделить выражение Е = Е0 - Е1 на Е: 1 = Е0/Е - Е1/Е и обозначить Е1/Е = c, то 1 = e - c. Величина c = Е1/Е называется коэффициентом поляризации (или электрической восприимчивости). Она характеризует соотношение электрического поля, созданного ориентированными диполями и результирующего поля в диэлектрике Е.
В неполярных диэлектриках связанные заряды образуют сферически симметричную систему, как например, в атоме водорода. В этом атоме электронные оболочки образуют сферу относительно положительно заряженного ядра (рис. 9-15). При воздействии внешнего электрического поля на связанные заряды действует кулоновская сила, которая вызывает нарушение симметрии. В данном случае деформированную систему связанных зарядов, у которых не совпадают центры, следует рассматривать как электрический диполь. А если это так, то для неполярного диэлектрика справедливы теоретические положения, рассмотренные выше. Следовательно, и в неполярном диэлектрике в результате деформационной поляризации происходит уменьшение внешнего электрического поля.
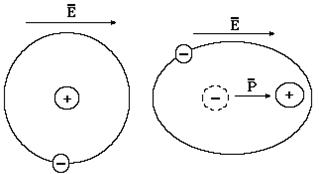
Рис. 9-15
Количественными характеристиками подобного взаимодействия служат относительная диэлектрическая проницаемость и коэффициент поляризации. Эти величины определяют электрические свойства диэлектриков и характеризуют степень дипольной или деформационной поляризации.
Дата добавления: 2016-01-05; просмотров: 54; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!