Тканевые потенциалы
Как показывают наблюдения, при возбуждении тканей (мышечной, нервной) возникает электрическая активность, которая и называется тканевыми биопотенциалами. Им принадлежит исключительно важная роль в диагностике функционального состояния тканей и органов. Рассмотрим механизм возникновения тканевых биопотенциалов. Нет никаких сомнений в том, что эта активность является следствием генерации клеточных потенциалов действия, поскольку все ткани состоят из клеток. Для упрощения анализа будем рассматривать возникновение тканевых потенциалов на физической модели, которая представляет собой мнослой возбудимых клеток, ориентированных вдоль оси 00' (рис. 8-10а). Как уже отмечалось, в состоянии покоя наружная поверхность клетки имеет более высокий потенциал, чем цитоплазма. Если все клетки монослоя находятся в одном и том же функциональном состоянии, то на внешней поверхности всех клеток будет один и тот же потенциал.

Рис. 8-10
Измерительное устройство, электроды которого установлены на первой и последней клетке монослоя, не зарегистрирует электрической активности. Другими словами, в состоянии покоя разность тканевых биопотенциалов равна нулю j2 - j1 = U = 0. Если теперь на одну из клеток монослоя воздействовать раздражителем R пороговой силы, наблюдается ее возбуждение и перезарядка клеточной мембраны – на поверхности появляется отрицательный потенциал по отношению к цитоплазме (рис. 8-10b). На внешней поверхности монослоя возникает разность потенциалов между рядом расположенными клетками, которая создает электрический ток. В данном случае электроды измерительного устройства оказываются в точках с различными потенциалами, и поэтому напряжение U = j2 - j1 > 0. За счет протекания локальных токов i снижается трансмембранная разность потенциалов у рядом расположенной клетки, и, когда она достигается критического значения, генерируется потенциал действия. При этом увеличивается отрицательный потенциал на внешней поверхности монослоя, и поэтому измерительный прибор покажет увеличение разности потенциалов. При последующем распространении возбуждения по клеткам разность потенциалов возрастает (рис. 8-11а) и достигает наибольшего значения в том случае, когда волна возбуждения достигнет середины расстояния между электродами. При возбуждении всех клеток монослоя (рис. 8-11b) электроды оказываются в точках с одним и тем же потенциалом. В этом случае разность потенциалов становится равной нулю. За процессом возбуждения следует восстановление исходного состояния. Поскольку длительность фазы возбуждения постоянна, первой клеткой, которая возвратится в исходное состояние, будет та, которая первая возбуждалась. При перезарядке мембраны на ее внешней поверхности возникает положительный потенциал, и поэтому электроды измерительного устройства опять оказываются в точках с разными потенциалами (см рис. 8-11с).
|
|
|
|
|
|

Рис. 8-11
Следует обратить внимание на тот факт, что в ходе восстановления исходного состояния ткани наблюдается изменение знака разности потенциалов j2 - j1 < 0. Другими словами, при восстановлении исходного состояния генерируется электрическая активность противоположного знака по сравнению с распространением процесса возбуждения. Как и в предыдущем случае, разность потенциалов будет возрастать до тех пор, пока волна восстановления исходного состояния не достигнет середины расстояния между электродами. Затем регистрируемое напряжение уменьшается и становится равным нулю после возвращения всех клеток в исходное состояние.
На рисунке 8-12 приведен график изменения электрического напряжения во времени в ходе распространения возбуждения и восстановления исходного состояния. Анализ этого графика (электрограммы) позволяет предложить количественные показатели для диагностической оценки изменения состояния возбудимой ткани. В качестве таких показателей в электродиагностике широко используются временные параметры, амплитуда электрической активности и форма тканевых биопотенциалов. Исходя из вышесказанного, время t1 (от начала действия раздражителя R до достижения максимального значения) потенциалов) описывает распространение возбуждения от первой клетки монослоя до середины расстояния между электродами, t2 – до возбуждения всего слоя.
|
|
|

Рис. 8-12
Поэтому период Т1 = t1 + t 2 характеризует длительность распространения возбуждения всех клеток в монослое. Интервал t3 описывает длительность, в течение которой все клетки остаются в возбужденном состоянии. Временной интервал T4 = t4 + t5 характеризует процесс восстановления исходного невозбужденного состояния ткани.
Следует отметить, что интервал t3 достаточно изменчив, а в некоторых случаях может равняться нулю (сразу за процессом возбуждения развивается восстановление исходного состояния). Очень часто не равны и временные интервалы Т1 и Т2 (обычно длительность распространения возбуждения меньше, чем Т2). Таким образом, временные интервалы электрограмм характеризуют скорость распространения возбуждения и восстановления исходного состояния тканей.
|
|
|
Амплитудные значение электрограмм Um1 и Um2 зависят от биопотенциалов отдельных клеток (потенциала покоя, амплитуды потенциала действия) и количества клеток, принимающих участие в генерации тканевых биопотенциалов. Чем больше потенциал покоя и амплитуда потенциала действия, тем выше амплитуда тканевых биопотенциалов. Поскольку потенциалы клеток в первую очередь зависят от соотношения концентраций электрогенных ионов в клетках и в среде, снижение амплитуды, выявляемое в клинической диагностике, наиболее часто обусловлено нарушением энергетических процессов в клетках (нарушением обмена веществ).
Форма электрических импульсов является комплексной характеристикой, зависящей от соотношения амплитудных и временных параметров электрограммы. Однако в клинической практике часто выявляется типичное нарушение формы тканевых биопотенциалов, характерное для определенных видов патологии. Поэтому форма тканевых биопотенциалов тоже используется в качестве диагностического критерия.
8.6. ОСНОВЫ ТЕОРИИ ДИПОЛЬНОГО ЭЛЕКТРИЧЕСКОГО ГЕНЕРАТОРА
Для описания закономерностей возникновения электрической активности в тканях произвольной формы разработана теория дипольного электрического генератора. Основные положения этой теории заключаются в следующем. В процессе распространения волны возбуждения и восстановления исходного состояния на внешней поверхности клеточных мембран возникает разность потенциалов. Другими словами, локальные электрические токи в среде создаются ЭДС - Е (рис. 8-13), где R – электрическое сопротивление среды, окружающей клетки. Электрическое поле, которое создает эта ЭДС можно описать с помощью представлений о дипольном электрическом генераторе.

Рис. 8-13
Напомним некоторые сведения из электродинамики. Электрическим диполем называется система из двух связанных электрических зарядов, равных по величине и противоположных по знаку, у которых не совпадают центры (см. рис. 8-14а).
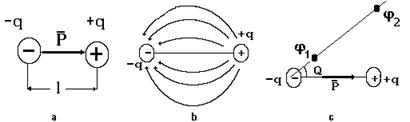
Рис. 8-14
Расстояние между центрами зарядов l называется плечом диполя. Важной характеристикой диполя служит электрический момент Р, величина которой определяется как произведение электрического заряда на плечо: P= q ´ l. Электрический момент – вектор, направленный вдоль плеча от отрицательного заряда к положительному. Вокруг диполя в окружающем пространстве возникает электрическое поле, силовые линии которого изображены на рисунке 8-14b. Электрическое поле описывается разностью потенциалов. Как показывает теория, разность потенциалов между любыми двумя точками пропорциональна электрическому моменту диполя и косинусу угла между направлением вектора момента и прямой проведенной через точки, в которых измеряется электрическое напряжение:
U ~ P·cos Q.
Используя понятие диполя, можно описать тканевые биопотенциалы. На рисунке 8-15 изображена монослойная модель, в которой развивается последовательное возбуждение клеток.

Рис. 8-15
На внешних поверхностях двух рядом расположенных клеток возникают различные электрические заряды (на возбужденной клетке отрицательный, на невозбужденной – положительный). Поэтому можно считать, что при возбуждении клеток появляется электрический диполь (отрицательный заряд на возбужденной, положительный на невозбужденной клетке). Плечо диполя в данном случае – это расстояние между центрами зарядов. Электрический момент ориентирован вдоль оси монослоя от отрицательного заряда к положительному, т.е. он всегда направлен от возбужденных клеток к невозбужденным. При распространении возбуждения на новые клетки (рис. 8-15b) увеличивается заряд (отрицательный заряд), длина плеча и, следовательно, электрический момент диполя. В данном случае, прямая проходящая через точки установки электродов измерительного прибора параллельна оси монослоя (т.е. cos Q = 1), и поэтому регистрируется наибольшая по амплитуде разность потенциалов. В ходе распространения возбуждения происходит возрастание электрического момента и амплитуды напряжения. Это происходит до тех пор, пока возбуждение не достигнет середины расстояния между точками установки электродов. В дальнейшем электрический момент и амплитуда тканевых биопотенциалов уменьшаются. Когда все клетки находятся в состоянии покоя или возбуждения, плечо диполя равно нулю, равен нулю электрический момент и поэтому в ткани не обнаруживается электрическая активность. В связи с тем, что в процессе распространения возбуждения и восстановления исходного состояния электрический момент диполя изменяется по величине и направлению, в окружающем пространстве возникает переменное электрическое поле. Именно поэтому говорят о дипольном электрическом генераторе.
Представление о таком генераторе позволяет описать электрические поля, возникающие вокруг возбужденных тканей практически любой формы. Рассмотрим, к примеру, некоторый орган, имеющий пространственную структуру. Пусть возбуждение возникает в некоторой точке "а" и распространяется по всему органу (заш- трихованный участок на рисунке 8-16).

Рис.8-16
Для каждого элементарного участка поверхности, разделяющей возбужденные и невозбужденные клетки, можно определить элементарный электрический момент (Р1, Р2...). Векторная сумма этих моментов позволяет рассчитать результирующий дипольный электрический момент, который в данном случае называется вектором возбуждения. Как уже отмечалось, этот вектор всегда направлен в сторону невозбужденных участков ткани. Зная направление вектора возбуждения, можно обосновать выбор точек для установки электродов измерительного прибора. Из теории следует, что максимальная разность потенциалов будет в том случае, если прямая, проходящая через точки установки электродов, параллельна вектору возбуждения (дипольного электрического момента). Поэтому в процессе возбуждения органа максимальное значение тканевых биопотенциалов обнаруживает прибор U1. Измерительный прибор U2 не будет регистрировать разность потенциалов, поскольку угол Q между вектором возбуждения и прямой, проходящей через электроды, равен 90о (cos Q = 0).
Дата добавления: 2016-01-05; просмотров: 44; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!