Механизмы электрогенеза
Все живые организмы в процессе своей жизнедеятельности генерируют электрические поля в очень широком диапазоне напряжений (от единиц микровольт до сотен вольт) и частот (от долей герца до сотен и даже тысяч герц). Электрическая активность является показателем состояния отдельных биологических структур (клеток, тканей и органов), и поэтому регистрация изменения электрического поля во времени (электрограмм) широко используется в электродиагностике заболеваний. В зависимости от биологического объекта принято различать следующие виды электрограмм: электрокардиограмму (ЭКГ) – запись электрической активности сердца, электроэнцефалограмму (ЭЭГ) – головного мозга, электромиграмму (ЭМГ) – мышцы, электрогастрограмму (ЭГГ) – желудка и др. Особенно широкое распространение в современной клинической медицине получила электрокардиограмма, анализ которой является обязательной процедурой для постановки диагноза при заболеваниях сердца. Электрические поля используются для информационных процессов, протекающих в живом организме. Раздражители любой природы (звуковые, тепловые, механические, химические) преобразуются в электрические поля в специальных биологических структурах - рецепторах.
В процессе восприятия параметры раздражителей (интенсивность, время действия) трансформируются в соответствующие показатели электрической активности – амплитуду разности потенциалов (биопотенциалов), частоту электрических импульсов и др. Более того, в основе работы нервной системы, где протекают жизненно важные информационные процессы, лежат механизмы генерации, проведения и переработки электрических (нервных) импульсов.
В связи с тем, что электрическая активность определяет состояние биологических структур, оказалось возможным с помощью внешних электрических воздействий (электростимуляции) управлять реакциями тканей и органов. Особенно существенное значение эта процедура имеет при заболеваниях, сопровождающихся нарушением нервной регуляции, например, при параличах.
Определяющая роль электрических полей для процессов жизнедеятельности организма, широкое использование электродиагностики в медицине, применение электрических воздействий для регуляции состояния тканей и органов требуют теоретического рассмотрения механизмов генерации биопотенциалов (электрогенеза), закономерностей распространения электрических импульсов по нервным волокнам, способов анализа электрограмм и выявления диагностических показателей. В зависимости от биологического субстрата, в котором возникает электрическая активность, принято различать клеточные и тканевые биопотенциалы. Клеточные регистрируются на уровне отдельных клеток, тканевые – в тканях и органах, представляя собой результат сложения (суперпозицию) клеточных электрических полей. В зависимости от состояния биологических структур различают биопотенциалы покоя и действия. Первые возникают в клетках, находящихся в состоянии относительного покоя (например, в расслабленной мышечном волокне). Потенциалы действия генерируются в том случае, когда биологический субстрат переходит из состояния покоя в возбужденное состояние (например, при сокращении сердца). Здесь мы отметим, что потенциалы покоя регистрируются только в отдельных клетках, потенциалы действия возникают на клеточном и органно-тканевом уровнях.
8.1. КЛЕТОЧНЫЙ ПОТЕНЦИАЛ ПОКОЯ
Все клетки без исключения окружены мембранами, отделяющими цитоплазму от окружающей среды. Поскольку мембрана является своеобразной границей раздела, в клетке можно зарегистрировать два вида потенциалов (точнее говоря, разности потенциалов). Между наружной поверхностью мембраны и окружающей средой U (рис. 8-1), а также между внутренней поверхностью и цитоплазмой существуют разность потенциалов U, которые называются поверхностным потенциалом. Эта электрическая активность возникает в результате электролитической диссоциации молекул, расположенных на поверхностях мембраны.
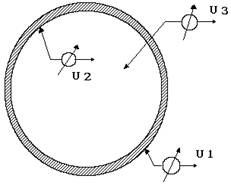
Рис. 8-1
Многочисленными исследованиями доказано, что на внешней поверхности всех без исключения клеток в результате диссоциации возникает отрицательный электрический заряд. За последние годы установлено, что этот заряд играет достаточно важную роль в межклеточных взаимодействиях. Так, он препятствует сближению клеток крови на расстояния, при которых образуются межмолекулярные связи, и наступает слипание (агрегация) клеток. Установлено, что при поражении ткани злокачественным ростом величина поверхностного заряда увеличивается, и, следовательно, возрастают силы межклеточного отталкивания. Полагают, что этот механизм лежит в основе отрыва клеток, их миграции по жидким средам организма и возникновения новых очагов злокачественного поражения (метастазирование злокачественных опухолей). До настоящего времени не ясна роль внутреннего поверхностного потенциала мембраны. Кроме рассмотренных видов электрических полей, можно зарегистрировать разность потенциалов U4 между цитоплазмой клетки и окружающей средой (см. рис. 8-1), т.е. электрическое поле в мембране. Эта электрическая активность получила название трансмембранной (мембранной) разности потенциалов - мембранных биопотенциалов.
8.2. МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
В основе механизма возникновения мембранных потенциалов лежит неравновесное распределение электрогенных ионов калия К+, натрия Na+ и хлора Cl- в цитоплазме и окружающей среде. Главную роль в возникновении трансмембранного электрического поля играют ионы К+ и Na+. Неравновесная концентрация этих ионов возникает в результате работы специальных молекулярных механизмов – ионных насосов. В процессе расщепления молекул АТФ освобождается энергия, которая используется для переноса ионов из области с меньшей в область с большей концентрацией: насос переносит ионы калия внутрь клетки, а ионы натрия в окружающую среду (рис. 8-2). Доказано, что на два иона калия транспортируется три иона натрия в окружающую среду. Именно поэтому в ходе работы насоса из клетки удаляется большее количество положительных зарядов, чем поступает из среды. Поэтому говорят, что ионный К-Na - насос является электрогенным, способствующим возникновению отрицательного потенциала внутри цитоплазмы. В процессе работы насосов в клетке создается избыточная концентрация ионов калия. В окружающей среде ионов натрия и хлора содержится больше, чем в цитоплазме клетки. Если существует неравновесная концентрация ионов, то возникает их диффузия из области с большей в область с меньшей концентрацией.
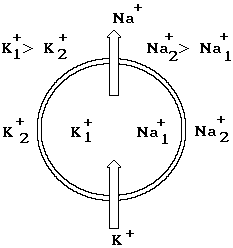
Рис. 8-2
Поэтому ионы калия будут диффундировать из клетки, а ионы натрия и хлора в цитоплазму (рис. 8-3). В процессе движения ионы переносят через мембрану электрический заряд. При этом соотношение положительных и отрицательных зарядов по обе стороны мембраны изменяется и на ней возникает электрическое поле.
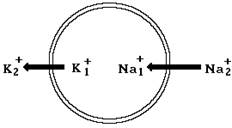
Рис. 8-3
Определим, от каких факторов будет зависеть величина разности потенциалов на мембране при диффузии ионов. В процессе переноса электрического заряда Q через мембрану совершается работа А, которые, как известно, связаны с возникающей разность потенциалов соотношением: А = Q (j1 - j2). Величина заряда Q = N Z e, где N – число перенесенных ионов, Z – количество элементарных зарядов е. Выразим N через постоянную Авогадро NA: N = nNA (n – количество перенесенных молей ионов). С учетом введенных обозначений затраченная работа на создание трансмембранного электрического поля будет равна:
A = n NA e Z (j1 - j2).
Обратим внимание на одно важное обстоятельство: в процессе диффузии зарядов возникает электрическое поле, которое тормозит движение соответствующего иона. Так, если диффундирует ион калия из клетки в среду, то в цитоплазме возникает недостаток, а в среде избыток положительных зарядов (цитоплазма заряжается отрицательно по отношению к среде). При этом возникает электрическое поле Е, которое направлено в цитоплазму и препятствует диффузии ионов калия. Поскольку работа по переносу зарядов совершается за счет энергии неравновесной концентрации,
E = n R T ln(C1/C2),
где С1 и С2 концентрации ионов калия по обе стороны мембраны, то процесс переноса будет уравновешиваться, когда энергия электрического поля (работа, затраченная на его создание) станет равной энергии неравновесной концентрации:
А = Е или n NA e Z (j1 – j2) = n R T ln(C1/C2).
Учитывая, что произведение числа Авогадро NA на элементарный заряд е представляет константу – число Фарадея F = NA e и, сокращая обе части равенства на число молей n, получаем уравнение Нернста, описывающее величину разности потенциалов (напряжения), возникающую на мембране:
j1 – j2 = (RT / ZF) ln(C1/C2).
Из этого соотношения видно, что напряжение, которое возникает при диффузии электрогенных ионов, зависит от соотношения их концентраций по обе стороны мембраны, и температуры Т. Обратим внимание также и на то, что в зависимости от направления диффузии и знака иона изменяется знак разности потенциалов. Например, ион натрия, имея положительный заряд, перемещается из окружающей среды в цитоплазму (из области с большей в область с меньшей концентрацией). Поэтому в процессе его движения цитоплазма приобретает положительный потенциал по отношению к среде (разность потенциалов противоположна по знаку напряжению, возникающему при диффузии ионов калия). Диффузия ионов хлора создает такую же разность потенциалов по знаку, что и ионы калия.
По-видимому, результирующее напряжение на клеточной мембране следует определять сложением (суперпозицией) электрических полей (с учетом их направления), созданных всеми электрогенными ионами. Другими словами, всю трансмембранную разность потенциалов можно рассчитать на основании использования формулы Нернста:
Um = j1 - j2 = UК + UNa + UCl =
= 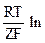 K2/K1 + Na2/Na1 + Cl2/Cl1
K2/K1 + Na2/Na1 + Cl2/Cl1
В этом соотношении обозначено: UК, UNa, UCl – калиевый, натриевый и хлорный потенциал, созданные в результате диффузии соответствующих ионов, К1, Na1, Cl1 - концентрации соответствующих ионов в клетке, К2, Na2, Cl2 - в среде. Поскольку K2<K1 , а Z = + 1, калиевый потенциал UК = K2/K1 <0 – диффузия ионов калия из цитоплазмы в среду способствует возникновению отрицательного потенциала в клетке по отношению в окружающей среде. Для натрия Na2 > Na1, отношение концентраций в среде и клетке больше нуля, логарифм этого отношения больше нуля и поэтому UNa = Na2/Na1 >0. Для хлора Cl2 > Cl1, но поскольку Z = –1, UCl = Cl2/Cl1< 0. Поэтому U = -UК + UNa - UCl
Как показывает более детальный анализ, определение результирующего напряжения на мембране с помощью формулы Нернста не совсем точно, потому что мембрана обладает неодинаковой проницаемостью к различным электрогенным ионам.
По современным представлениям, движение ионов калия и натрия из области с большей в область с меньшей концентрацией осуществляется через специальные ионноселективные мембранные каналы. На 1 кв. мкм площади мембраны располагается несколько сотен каналов. В состоянии покоя клеток функционирует больше калиевых каналов и поэтому проницаемость мембраны для этого иона значительно больше, чем для иона натрия. Если принять проницаемость для иона калия РК за 1, то соотношение проницаемостей имеет следующий вид РК: РNa: РСl= 1: 0,04: 0,45. Из приведенных соотношений понятно, что на результирующий мембранный потенциал решающее влияние оказывает диффузия ионов калия. Количественно мембранное напряжение описывается формулой Гольдмана-Ходжкина, учитывающей соотношение концентраций электрогенных ионов и соответствующие проницаемости мембраны:
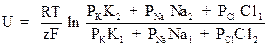
В этой формуле обозначено: PK, PNa, PCl – проницаемости мембраны для соответствующих ионов, а K1, Na1, Cl1 – концентрации ионов внутри клетки, а K2, Na2, Cl2 – в окружающей среде. Подсчет мембранного потенциала по этой формуле дает величину, которая измеряется в эксперименте.
Если в первом приближении пренебречь в формуле Гольдмана-Ходжкина величинами, содержащими проницаемости для натрия и хлора, то:
U 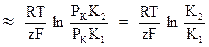
Последнее соотношение представляет собой уравнение Нернста, описывающее калиевый потенциал. В состоянии покоя проницаемость мембраны наиболее высока для ионов калия, и поэтому можно считать, что величина мембранного потенциала приблизительно равна Нернстовскому калиевому потенциалу. Если измерить мембранный потенциал в нервной клетке и сравнить его с рассчитанным значением по формуле Нернста, то получаются соответственно следующие величины –75 мВ и –80 мВ. Это значит, что в состоянии покоя калиевый потенциал практически равен мембранному, а игнорирование других ионов по формуле Нернста дает ошибку всего в 5 мВ. Отметим, что знак «минус» говорит о том, что цитоплазма клетки заряжена отрицательно по отношению к окружающей среде, поскольку ионы калия выносят положительный заряд из клетки. Совершенное точное значение мембранного потенциала получается по формуле Гольдмана-Ходжкина.
8.3. ПОТЕНЦИАЛ ДЕЙСТВИЯ НЕЙРОНА
При раздражении нервных клеток (нейронов) электрическим импульсом или некоторыми химическими веществами наблюдается процесс возбуждения и генерация нервных импульсов. По своей природе нервный импульс является электрическим. Поэтому процесс возбуждения нейронов следует рассматривать как изменение их состояния в результате генерации электрической активности.
По современным представлениям возбуждение нейронов происходит в том случае, когда интенсивность действующего раздражителя достигает определенной величины, называемой порогом возбуждения. В этом случае наблюдается изменение проницаемости клеточной мембраны за счет образования дополнительных каналов для ионов натрия и калия. На рисунке 8-4 представлены изменения проницаемости для соответствующих ионов во времени.
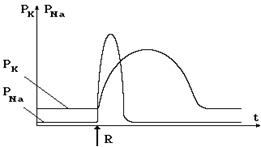
Рис. 8-4
Из графика видно, что проницаемость для ионов натрия очень быстро возрастает во времени, а затем также быстро возвращается к исходному значению. Время, в течение которого проницаемость для натрия превышает соответствующую проницаемость для калия, составляет порядка 1 мс. При повышении проницаемости для натрия эти ионы в большем количестве поступают внутрь клетки. Поскольку они переносят положительный заряд, наблюдается нейтрализация отрицательного потенциала цитоплазмы, и когда проницаемость для калия сравняется с проницаемостью для натрия (первая точка пересечения графиков), в первом приближении можно считать, что разность потенциалов становится равной нулю (при таком приближении не учитывается вклад ионных насосов в создание разности потенциалов на мембране).
Затем проницаемость для натрия превышает проницаемость для калия, внутрь клетки в единицу времени входит больше положительных зарядов, чем выходит в окружающую среду. Именно поэтому цитоплазма клетки перезаряжается и становится положительной. Величина положительного потенциала возрастает по мере нарастания проницаемости для ионов натрия, а потом, когда последняя уменьшается, происходит снижение мембранной разности потенциалов. При равенстве проницаемостей (вторая точка пересечения графиков) разность потенциалов становится равным нулю. За 1 импульс через 1 кв. мкм мембраны проходит около миллиона ионов натрия.
Как следует из графика на рис. 8-4, проницаемость для ионов калия также возрастает, но с меньшей скоростью. Когда проницаемость для натрия возвращается к исходной величине, проницаемость для калия остается большей, чем в состоянии покоя. Это значит, что в это время больше калия выходит из клетки, чем в состоянии покоя. Поэтому на мембране регистрируется разность потенциалов по величине большая, чем в покое. Сопоставление динамики проницаемости мембраны для калия и натрия с изменением мембранной разности потенциалов приведено на графике на рис. 8-5. Зависимость этого напряжения от времени называется потенциалом действия или нервным импульсом.
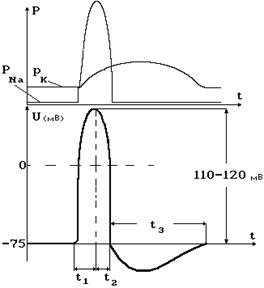
Рис. 8-5
Принято различать следующие временные интервалы (фазы) потенциала действия: фазу деполяризации t1 (время от начала изменения мембранного потенциала до достижения максимума), фазу реполяризации t2 (востановления потенциала покоя) и фазу гиперполяризации t3 (время, в течение которого потенциал на мембране превышает потенциал покоя). Длительность нервного импульса T = t1 + t2 – это практически то время, в течение которого проницаемость мембраны для ионов натрия превышает проницаемость для калия. Как уже отмечалось, эта величина составляет порядка 1 мс. Фаза гиперполяризации продолжается в течение большего времени: 10-100 мс. Амплитуда потенциала действия достигает 110-120 мВ.
Дата добавления: 2016-01-05; просмотров: 66; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!