Характеристика генетических и негенетических факторов регуляции экспрессии генов
| Название регуляторных белков и эффекторов | Точка приложения | Функция |
| Белки-репрессоры | Присоединяются к оператору — регуляторной последовательности ДНК, расположенной между промотором (частично перекрывают промотор) и генами оперона | Препятствуют соединению РНК- полимеразы с промотором и снижают уровень транскрипции генов оперона. Регуляция экспрессии генов на основе белков-репрессоров называется негативной, так как в ее основе лежит репрессия оперона регуляторным белком |
| Белки-активаторы (апоиндукторы) | Присоединяются к актива- тору — регуляторной последовательности ДНК, предшествующей промотору и частично перекрывающей его | Облегчают связывание РНК- полимеразы с промотором, стимулируя тем самым транскрипцию. Регуляция экспрессии генов на основе белков-активаторов называется позитивной |
| Индукторы | Связываются с активными белками-репрессорами и инактивируют их. Связываются с неактивными белками-активаторами и активируют их | «Включают» транскрипцию генов. Регуляция экспрессии генов на основе белков-репрессоров и индукторов называется негативной индукцией. «Включают» транскрипцию генов. Регуляция экспрессии генов на основе белков-активаторов и индукторов называется позитивной индукцией |
| Корепрессоры | Связываются с неактивными белками-репрессорами и активируют их. Связываются с активными белками-активаторами и инактивируют их | «Выключают» транскрипцию генов. Регуляция экспрессии генов на основе белков-репрессоров и корепрессоров называется негативной репрессией. «Выключают» транскрипцию генов. Регуляция экспрессии генов на основе белков-активаторов и корепрессоров называется позитивной репрессией |
В 1961 г. французские ученые Ф. Жакоб и Ж. Моно предложили оперонную модель регуляции экспресии прокариотических генов, объясняющую механизм контроля синтеза белков у прокариот путем «включения» и «выключения» тех или иных генов в зависимости от потребности бактериальной клетки в метаболитах. Согласно этой модели, структурные гены оперона, кодирующие белки, участвующие в одной цепи биохимических преобразований, располагаются рядом друг с другом и образуют единый функциональный блок. Все гены оперона имеют общие регуляторные элементы (промотор, оператор, терминатор и др.) и транскрибируются, образуя одну полицистронную мРНК, с которой транслируются несколько разных полипептидов. Важное преимущество, которое дает объединение разных генов в один оперон, состоит в том, что это позволяет осуществлять координированный контроль экспрессии генов путем одновременного их «включения» или «выключения». Например, при локализации в одном опероне генов, кодирующих ферменты, катализирующие реакции одного катаболического пути, синтез каждого из них в ответ на появление соответствующего субстрата в окружающей среде осуществляется одновременно.
|
|
|
|
|
|
Классическим примером регуляции экспрессии генов у прокариот служит функционирование лактозного (lac) оперона кишечной палочки, в состав которого входят три структурных гена, кодирующих белки-ферменты, участвующие в усвоении лактозы: β-галактозидазу (бета-галактозидазу), пермеазу и трансацетилазу, а также регуляторные последовательности для связывания белка- репрессора и белка-активатора — CRP- белка (от англ. catabolite repressor protein) (рис. 4.31).
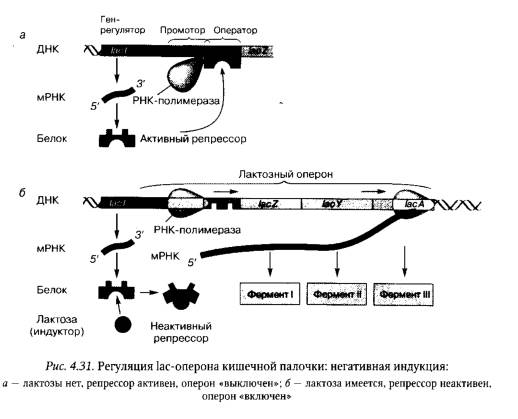
При выращивании Е.coli на среде, содержащей только глюкозу, ген-регулятор lас-оперона синтезирует активный белок-репрессор, который, взаимодействуя с оператором, «выключает» транскрипцию структурных генов, кодирующих ферменты, необходимые для усвоения лактозы (см. рис. 4.31). Если же клетки Е.coli перенести на среду, содержащую только лактозу, то, проникая внутрь клеток, небольшая часть лактозы превращается в аллолактозу. Аллолактоза, связываясь с белком-репрессором, инактивирует его. В результате РНК-полимераза осуществляет транскрипцию полицистронной мРНК для синтеза всех ферментов, необходимых для транспорта и метаболизма лактозы. В данном случае регуляция экспрессии генов оперона осуществляется на основе негативной индукции. При этом индуктором генов Lас-оперона служит лактоза. Опероны, регулируемые подобным образом, получили название индуцибельных, или индуцируемых, оперонов.
|
|
|
При культивировании кишечной палочки на среде, содержащей как лактозу, так и глюкозу, бактерии утилизируют только глюкозу. Подобная избирательность в выборе субстрата обусловливается наличием у Е. coli механизма положительной регуляции активности генов1ас-оперона (рис. 4.32).
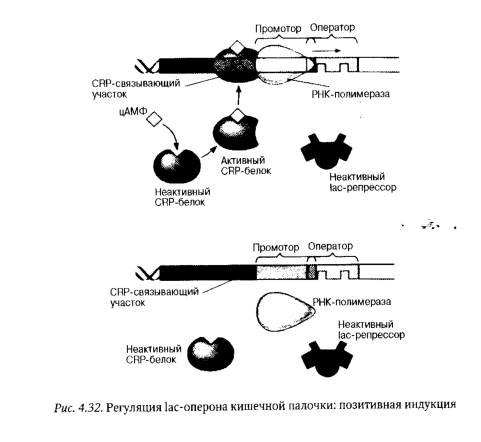
Роль активатора 1ас-оперона играет CRP-белок, который сам по себе неактивен и не может связываться с регуляторными последовательностями ДНК промотора. Эту способность CRP-белок приобретает лишь в комплексе с низко молекулярным соединением — циклическим аденозинмонофосфатом (цАМФ), который накапливается в клетках кишечной палочки при отсутствии в культуральной среде, где они выращиваются, глюкозы1. CRP-белок в составе комплекса с цАМФ связывается с регуляторной последовательностью, частично перекрывающей промотор, и облегчает присоединение к нему РНК-полимеразы, стимулируя тем самым транскрипцию генов 1ас-оперона. При наличии в среде глюкозы и лактозы CRP-белок остается неактивным, и гены 1ас-оперона практически не транскрибируются. Это объясняется низким аффинитетом фермента РНК-полимеразы к промотору 1ас-оперона и неспособностью указанного фермента связываться с промотором в отсутствие активного активатора.
|
|
|
Двойной контроль экспрессии генов 1ас-оперона с участием белка-репрессора и белка-активатора позволяет кишечной палочке не только быстро перестраивать свой метаболизм в изменяющихся условиях среды, поскольку содержимое кишечника человека, где эта бактерия обитает, характеризуется большим непостоянством, но и осуществлять биологически оправданный выбор субстрата для реакции брожения. Действительно, при наличии в среде глюкозы и лактозы бактерии кишечной палочки усваивают только глюкозу, достигая этим наиболее экономного расходования имеющихся у них энергетических ресурсов. Объясняется это тем, что, в отличие от глюкозы, использование в качестве энергетического субстрата лактозы требует предварительного расщепления молочного сахара на глюкозу и галактозу, что сопряжено с дополнительными затратами энергии.
У кишечной палочки регуляция экспрессии генов осуществляется не только на основе негативной индукции, но и посредством негативной репрессии. Примером такой регуляции служит функционирование триптофанового ( trp ) оперона, содержащего 5 структурных генов, кодирующих ферменты, необходимые для синтеза аминокислоты триптофана из ее предшественников (рис. 4.33).
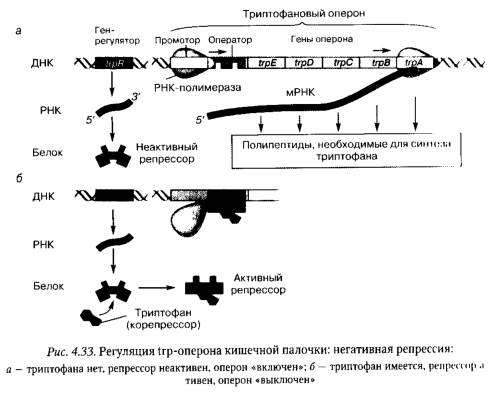
При недостатке в клетке триптофана белок-репрессор, оставаясь неактивным, не препятствует РНК-полимеразе присоединяться к промотору trp-оперона и транскрибировать полицистронную мРНК, кодирующую ферменты, необходимые для синтеза этой аминокислоты из ее предшественников. При накоплении триптофана в клетке указанная аминокислота связывается с белком-репрессором и, действуя как корепрессор, активирует его. Активированный белок-репрессор связывается с оператором и блокирует присоединение РНК-полимеразы к промотору. В результате trp-onepoн «выключается» и синтез триптофана прекращается. Опероны, регулируемые подобным образом, получили название репрессибельных оперонов.
Дата добавления: 2021-03-18; просмотров: 462; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!