Использование неорганических доноров водорода.
Многие группы почвенных и водных бактерий могут использовать в качестве доноров водорода или электронов неорганические соединения или ионы (ионы аммония, нитрита, сульфида, тиосульфата, сульфита и двухвалентного железа), а также элементарную серу, молекулярный водород и СО2 т.е. способны получать в результате их окисления восстановительные эквиваленты и энергию для синтетических процессов. Получение энергии происходит, как правило, в результате дыхания с О2, как конечным акцептором водорода. Электроны, освобождающиеся при окислении упомянутых неорганических субстратов, поступают в дыхательную цепь на уровне цитохрома с или цитохрома а. Поскольку фосфорилирование при этом может происходить лишь на одном этапе окисления, выигрыш в энергии соответственно невелик. Часть этой энергии затрачивается на то, чтобы «оттеснить» электроны, поступающие на участок цитохромов, по дыхательной цепи назад, на уровень пиридиннуклеотидов (восстановительные эквиваленты для синтезов), и восстановить последние. Лишь немногие из относящихся к этой группе бактерий способны расти за счет «анаэробного дыхания», используя в качестве акцепторов водорода нитрат, нитрит, закись азота и т. п. Такой образ жизни с использованием неорганического донора водорода называютхемолитотрофным.Большинство бактерий с таким типом метаболизма используют СО2 в качестве единственного или главного источника клеточного углерода. Они являются поэтому автотрофами (хемолитоавтотрофами). Почти все аэробные хемолитоавтотрофные бактерии, до сих пор изученные в этом отношении, ассимилируют углерод СО2 через рибулозобисфосфатный цикл.
|
|
|
Превращение аммиака (аммония) в нитрат- нитрификация - осуществляется двумя группами нитрифицирующих бактерий: одни окисляют аммиак, образуя нитрит, а другие окисляют нитрит в нитрат:
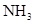 à
à  à
à 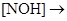

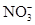
При окислении нитрита электроны переносятся на цитохром а1.
Различные соединения серы могут окисляться тиобациллами до сульфата:
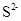 à Sà
à Sà 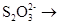
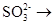

Электроны, высвобождающиеся при окислении сульфита до сульфата, поступают вдыхательную цепь. Некоторые тиобациллы способны использовать выделяющуюся при этом окислении энергию для фосфорилирования на уровне субстрата:




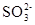 аденозинфосфосульфат
аденозинфосфосульфат
АМР 2 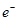 Pi ADP
Pi ADP
Железобактерии окисляют двухвалентное железо до трёхвалентного:
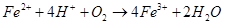
Почти во всех группах бактерий, синтезирующих АТР путем окислительного фосфорилирования в анаэробных условиях («анаэробное дыхание»), есть формы, способные использовать молекулярный водород в качестве донора электронов. Существуют также аэробные бактерии, окисляющие водород с использованием кислорода в качестве конечного акцептора электронов.
|
|
|
Ряд бактерий способны использовать СО в качестве единственного источника электронов, окисляя его до СО2.
Фиксация СО 2.
Большинство бактерий , способных расти в условиях, когда единственным источником углерода служит углекислота, фиксируют её через рибулозобифосфатный цикл. К таким бактериям относятся аэробные хемолитоавтотрофные, фототрофные и цианобактерии. Рибулозобифосфатный цикл – это восстановительный процесс, в котором СО2 восстанавливается до уровня углеводов. На первом этапе при участии рибулозобифосфат-карбоксилазы к рибулозо-1,5-бифосфату присоединяется СО2, и в результате образуются две молекулы 3-фосфоглицериновой кислоты. [ Этот же фермент в отсутствие СО2 и в присутствии О2 окисляет рибулозобифосфат до фосфогликолата и 3-фосфоглицерата. Эта реакция участвует в образовании гликолевой кислоты у автотрофных бактерий (в фотодыхании)]. Затем 3-фосфоглицерат восстанавливается до глицеральдегид-3-фосфата за счёт NAD(P)H2 и гидролиза АТР. На последнем этапе цикла глицеральдегид-3-фосфат и его изомер дигидроксиацетонфосфат участвуют в ряде реакций, приводящих к регенерации рибулозобифосфата, и цикл замыкается.
|
|
|
Часть триозофосфатов выводится из цикла и используются для синтеза гексозофосфатов и других сахаров. Ряд промежуточных продуктов также выводятся из цикла и используются для разнообразных синтезов.
Анаэробные автотрофные бактерии располагают двумя другими механизмами ассимиляции СО2. Метанобразующие, ацетогенные и сульфатредуцирующие бактерии, способные использовать в качестве доноров электронов Н2 или СО, восстанавливают СО2 по анаэробному ацетил-СоА пути до ацетил-СоА и пирувата. Зелёные серобактерии фиксируют СО2 исключительно с помощью реакций восстановительного цикла трикарбоновых кислот; СО2 фиксируется благодаря восстановительному карбоксилированию сукцинил-СоА.
Фотосинтез .
Фотосинтез – происходящее в клетках фототрофных организмов преобразование световой энергии в биохимически доступную энергию (АТР) и восстановительную силу [NAD(P)H2], а также связанный с этим синтез клеточных компонентов. Фотосинтетическое фосфорилирование и фотосинтетическое восстановление пиридиннуклеотидов – это процессы, ведущие к образованию первых стабильных продуктов фотосинтеза. АТР и NAD(P)H2 используются затем для фиксации СО2 и дальнейших процессов биосинтеза. Организмы, использующие воду в качестве донора электронов, осуществляют оксигенный фотосинтез, идущий с выделением кислорода. Организмы, использующие в качестве донора электронов другие вещества (с более высокой степенью восстановления – Н2S, H2, органические вещества), осуществляют аноксигенный фотосинтез, идущий без выделения кислорода. Фотосинтез происходит в мембранах или на их поверхности, а фиксация СО2 – в цитоплазме.
|
|
|
Мембрана содержит в себе пигментные молекулы, переносчики электронов и ферменты. Подавляющее большинство молекул (бактерио)хлорофилла, а также дополнительные пигменты(каротиноиды, фикобилипротеины) образуют систему антенны, ответственную за поглощение света и распределение энергии. Незначительная часть молекул (бактерио)хлорофилла выполняет роль фотохимического реакционного центра, в котором протекает собственно фотохимическая окислительно-восстановительная реакция. Пигменты антенн улавливают свет и передают энергию первичному донору реакционного центра [комплекс (бактерио)хлорофилла с белками]. Под воздействием энергии света донор передаёт электрон первичному акцептору реакционного центра и сам окисляется (возникает «дырка»). Дырка затем заполняется электроном от какого-либо внешнего донора. От первичного акцептора электроны проходят через ряд переносчиков, в конце восстанавливая NADP. При оксигенном фотосинтезе работают две фотосистем, связанные между собой электрон транспортной цепью, важным звеном в которой является пластохинон, который подобно убихинону в дыхательной цепи находится в большом избытке и выполняет функцию депо электронов. Фотосинтетический перенос электронов показан на следующей схеме:


 Fe-S
Fe-S
-белок


 цит.b Fd NADP
цит.b Fd NADP
 X320 пласто- циклический
X320 пласто- циклический
 хинон поток электронов
хинон поток электронов
цит.f
 пластоцианин пигменты
пластоцианин пигменты


 пигменты антенн h
пигменты антенн h 

 антенн h Хл.а.
антенн h Хл.а.
 Хл.а.
Хл.а. 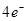
 2H2O
2H2O
O2 + 4 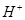
Две фотосистемы вместе со связывающей их электрон-транспортной цепью обеспечивают направленный поток электронов от воды (с внутренней стороны мембраны) к NADP(с внешней стороны). Перенос 1 электрона через обе фотосистемы сопровождается выведением 2 протонов во внешнюю среду (с участием пластохинона). В результате мембрана аккумулирует энергию в форме протонного потенциала, и эта энергия используется для синтеза АТР с помощью АТРазы или для совершения какой-либо работы.
В аноксигенном фотосинтезе участвует только одна фотосистема: она поддерживает циклический транспорт электронов. В качестве доноров электронов могут использоваться сероводород, сера, тиосульфат, органические соединения (малат, сукцинат и др.) и молекулярный водовод. Фотосинтетический перенос электронов приводит к созданию протонного градиента; для восстановления NADH2 требуется обратный транспорт электронов, протекающий с затратой энергии.
В плазматической мембране галобактерий выделяются тёмно-красные пятна, образованные так называемой пурпурной мембраной. Её цвет обусловлен наличием в ней бактериородопсина. Благодаря этому пигменту на свету создаётся протонный градиент между наружной и внутренней сторонами мембраны, т.е. энергия света превращается в одну из конвертируемых форм энергии.
Дата добавления: 2020-12-12; просмотров: 126; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!