Подведение итогов занятия преподавателем и проверка правильности выполнения работы каждым студентом.
1.6. Место и время занятия : учебная комната, 3 академических часа.
1.7. Оснащение занятия: таблицы, схемы.
1.8. Литература: основная (I) и дополнительная (II):
(I) 1. Учебник Биология в 2-х книгах /В.Н. Ярыгин, В.В. Глинкина,
И.Н. Волков, В.В.Синельщикова, Г.В. Черных. / М.,ГЭОТАР-Медиа, 2013. - Т.1. - 736 с.: ил. - Т.2. - 560 с.: ил.
2. Учебник Биология / А.А. Слюсарев, Н.В. Жукова/ Киев: Вища школа, 1987,415 с.
(II) 3. Медицинская паразитология/ Под ред. Н. В. Чебышева/ М.:ОАО-Издательство "Медицина", 2012. - 304 с.: ил.
4. Медицинская паразитология и паразитарные болезни/ Под ред. А.Б. Ходжаян, С.С. Козлова, М.В. Голубевой/ М.: ГЭОТАР-Медиа, 2014. - 448 с. : ил.
5. Биология с основами медицинской генетики/ Демиденко Л.А./ 3-е изд.- Симферополь, 2013. – 140 с.
6. Сборник тестов по медицинской биологии/ Авторский коллектив:
Кутя С.А. Демиденко Л.А. Ромашова М.Ф. Казакова В.В. Лященко О.И./
Симферополь, 2014. –– 96 с.
7. Методические разработки по цитологии к практическим занятиям по медицинской биологии/ С.А. Кутя ,М.Ф. Ромашова / Симферополь, 2014, - 88 с.
8. Методические разработки по генетике к практическим занятиям по медицинской биологии/ С.А Кутя., В.В. Казакова, М.Ф. Ромашова / Симферополь, 2014, - 88 с.
9. Методические разработки по паразитологии и эволюции к практическим занятиям по медицинской биологии/ С.А. Кутя ,Л.А. Демиденко ,М.Ф. Ромашова / Симферополь, 2014,- 212 с.
ЗАНЯТИЕ 4
Характеристика нуклеиновых кислот.
СТРОЕНИЕ ГЕНА ПРО – И ЭУКАРИОТ. ГЕНЫ СТРУКТУРНЫЕ, РЕГУЛЯТОРНЫЕ, т-РНК, р-РНК. ОРГАНИЗАЦИЯ ПОТОКА ИНФОРМАЦИИ В КЛЕТКЕ. РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНОВ.
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ИЗМЕНЧИВОСТИ У ЧЕЛОВЕКА.
1.1. Значение темы: Молекулярно—биологические представления о строении и функционировании генов позволяют будущим врачам глубже понять процессы нормальной жизнедеятельности человека, механизмы наследственной патологии (на молекулярном уровне), перспективы генной инженерии и биотехнологии.
1.2. Цели занятия. Общая — знать принципы организации наследственного материала эукариот, этапы механизма реализации наследственной информации и уметь связывать основные процессы молекулярно-генетической регуляции жизнедеятельности организма с патологией человека.
Знать принципы строения гена про- и эукариот, функциональные характеристики гена, экзонно-интронную организацию генома эукариот, а также генетический код и его свойства.
Знать этапы механизма реализации наследственной информации и уметь связывать основные процессы молекулярно-генетической регуляции жизнедеятельности организма с патологией человека.
1.3. Конкретные цели. Уметь:
1.3.1. Оперировать понятиями молекулярной биологии о структуре и функциях генов, реализации наследственной информации в нормальные и патологические признаки.
1.3.2. Объяснять процесс молекулярно-генетической регуляции активности генов.
1.3.3. Оперировать понятиями молекулярной биологии о структуре и функциях генов.
1.3.4. Объяснять основные свойства генетического кода и решать соответствующие задачи.
1.3.5. Оперировать понятиями молекулярной биологии о реализации наследственной информации в нормальные и патологические признаки.
1.3.6. Объяснять процесс молекулярно-генетической регуляции активности генов.
Основные теоретические сведения
Генетическая информация в клетке связана с нуклеиновыми кислотами. Их два типа: дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК). Мономерными структурными единицами нуклеиновых кислот являются нуклеотиды. Нуклеотид состоит из молекулы фосфорной кислоты, моносахарида (дезоксирибозы — ДНК или рибозы — РНК) и одного из 4-х азотистых оснований: аденина (А), гуанина (Г), цитозина (Ц) и тимина (Т) — ДНК или урацила (У) — РНК (рис.13).
Модель строения молекулы ДНК предложили в 1953 году Ж. Уотсон и Ф. Крик. Основные свойства этой модели следующие:
1. ДНК образована 2 полинуклеотидными цепями, при этом соединение нуклеотидов в цепь обеспечивается ферментом ДНК-полимеразой.
2. Цепи правозакручены спирально вокруг общей оси, один виток включает 10 пар нуклеотидов.
3.Фосфатные группировки находятся снаружи спирали, а азотистые основания внутри.
4. Цепи антипараллельны, то есть последовательность атомов одной цепи противоположна таковой в другой.
5. Цепи удерживаются водородными связями между азотистыми основаниями по принципу комплементарности: аденин соединяется двумя связями с
тимином, гуанин тремя связями с цитозином (следует заметить, что водородные связи слабые и нарушаются при изменении рН клетки или нагревании до 1000 С.
Нуклеотиды различаются только по азотистым основаниям. Название нуклеотидов также связано с названием этих оснований: нуклеотид, содержащий аденин, называется адениловым, гуанин - гуаниловым, тимин - тимидиловым, цитозин - цитидиловым.
Две цепочки, состоящие из десятков и сотен нуклеотидов, скручиваются между собой и образуют двойную спираль. Диаметр ее равен 2x10-9 м (2нм), а расстояние между соседними нуклеотидами – шаг молекулы – 3,4x10-10 м (0,34нм). Десять нуклеотидных пар образуют полный виток двойной спирали (шаг спирали) длиной 3,4x10-9 м (3,4нм). 1нм = 10-9 м.
Азотистые основания одной спирали ДНК располагаются точно против азотистых оснований второй спирали. При этом обнаруживается важная закономерность: против тимина одной спирали всегда располагается аденин другой спирали, против цитозина – гуанин и наоборот. Соединения других типов никогда в норме не возникают. Это обьясняется тем, что между такими парами нуклеотидов, как аденин - тимин, гуанин - цитозин, существует особый тип химической связи, получивший название комплементарности (лат комплемент - дополнение). При этом между А и Т образуются две водородные связи, а между Г и Ц - три.
Репликация (редупликация) ДНК (репликатио - повторение, редупликатио -удвоение) – процесс самовоспроизведения макромолекул нуклеиновых кислот, обеспечивающий точное копирование генетической информации и передачу ее от поколения к поколению. Самоудвоение молекулы ДНК происходит в период интерфазы перед делением клеток. При этом молекула ДНК, состоящая из цепей нуклеотидов, начинает раскручиваться и раздваиваться с одного конца. Около каждой из двух цепей из свободных нуклеотидов, находящихся в ядре клетки, по принципу комплементарности синтезируется вторая цепь ДНК. В результате вместо одной молекулы ДНК образуются две, причем обе являются точными копиями исходной (материнской) молекулы. Таким образом, в каждой новой молекуле ДНК одна цепь происходит из первоначальной молекулы ДНК, а вторая синтезируется вновь. Как процесс расхождения молекулы на две цепи, так и процесс синтеза осуществляется за счет деятельности ряда ферментов.
По современным представлениям ДНК имеет две функции.
- Аутосинтетическая — способность к самоудвоению в синтетическом периоде интерфазы.
- Гетеросинтетическая — передача информации о структуре белка на молекулу и-РНК, которая играет главную роль в процессе биосинтеза белка в клетке.
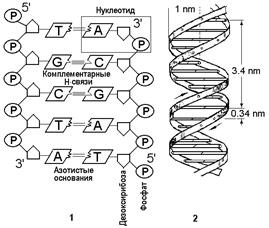
Рис.13 Схема участка молекулы ДНК(1) и ее спиральной структуры(2)
В ходе репликации после удвоения ДНК в ней обнаруживаются ошибки, возникающие под действием различных факторов физической и химической природы - при этом возникает репарация - самовосстановление первичной структуры ДНК, которая может быть экцизионной, фотерепарацией и рекомбинантной репарацией.
Молекулы РНК также представляют собой полинуклеотиды, но состоят из одной цепи, которая включает рибозу, фосфорную кислоту и 4 типа азотистых оснований: аденин, гуанин, цитозин и урацил (близкий по строению к тимину). Соответственно имеется 4 типа нуклеотидов: адениловый, гуаниловый, цитидиловый и урациловый. Молекулы РНК гораздо короче, чем молекулы ДНК, и имеют значительно меньшую молекулярную массу, которая редко превышает 100 тыс.у.е.
Существует несколько видов РНК, отличающихся по структуре и функциям.
Рибосомальные РНК (р-РНК) содержатся в рибосомах.
Транспортные РНК (т-РНК) самые небольшие по размеру, участвуют в транспортировке аминокислот к месту синтеза белка.
Информационные РНК (и-РНК) синтезируются на участке одной из цепей молекулы ДНК и передают информацию о структуре белка из ядра клеток к рибосомам.
Транспортные РНК (т-РНК) имеет форму трилистника и содержит 3 петли. Антикодоновая петля имеет антикодон, то есть триплет нуклеотидов, соответствующий своей аминокислоте. Псевдоуридиновая петля необходима при взаимодействии с рибосомой. Дигидроуридиновая петля служит для соединения с ферментом аминоацилсинтетазой.
По современным представлениям ген – участок молекулы ДНК, содержащий информацию о синтезе одного полипептида. Различают гены – структурные, которые кодируют определенные белки и регуляторные – направляют деятельность структурных генов.
Последовательность аминокислот в белке определяется последовательностью нуклеотидов в молекуле ДНК, его генетическим кодом. Т. о., наследственная информация на ДНК записана с помощью генетического кода.
Код имеет следующие основные свойства:
1. Триплетность — одну аминокислоту кодируют три нуклеотида.
2. Вырожденность (избыточность) — одну аминокислоту кодируют от двух до четырех триплетов. Всего имеется 64 триплета: 61 триплет несет информацию об аминокислотах, а 3 стоп-кодона обозначают окончание синтеза полипептидной цепи.
3. Неперекрываемость — нуклеотид одного триплета не может входить в состав соседнего триплета.
4. Универсальность — код един для всех живых организмов (синтез белка).
Генетический код и-рнк
| первый нуклеотид триплета (5 – конец) | второй нуклеотид триплета | третий нуклеотид триплета (3-конец) | |||
| а | г | у | ц | ||
| а | лиз лиз асн асн | арг арг сер сер | иле мет иле иле | тре тре тре тре | а г у ц |
| г | глу глу асп асп | гли гли гли гли | вал вал вал вал | ала ала ала ала | а г у ц |
| у | стоп стоп тир тир | стоп три цис цис | лей лей фен фен | сер сер сер сер | а г у ц |
| ц | глн глн гис гис | арг арг арг арг | лей лей лей лей | про про про про | а г у ц |
СИНТЕЗ БЕЛКА включает несколько уровней: 1. Претранскрипционный
2. Транскрипционный 3. Транспортный 4. Трансляционный 5. Посттрансляционный. (рис. 13).
1. Претранскрипционный уровень: стартовый этап синтеза. Во время него происходит активация молекулы ДНК с помощью специальных белков.
2. Транскрипционный уровень: Транскрипция - синтез и-РНК на молекуле ДНК с последовательностью нуклеотидов комплементарной молекуле ДНК. Матрицей для и-РНК служит только одна из двух имеющихся цепей ДНК, то есть синтез носит полуконсервативный характер. Процесс контролируется ферментами РНК-полимеразами.
В молекуле ДНК выделяется определенный фрагмент- промотор как начальный участок синтеза. За ним следуют информационные нуклеотидные кодоны. Существует 2 вида наборов нуклеотидов. Триплеты, несущие информацию об аминокислотах – экзоны. Они образуют короткие сегменты ДНК и, как правило, разделены более длинными неинформативными участками нуклеотидов, которые называют интронами. В каждом гене количество экзонов на один превышает число интронов. С началом транскрипции в первичную и-РНК включаются как экзоны, так и интроны. Первичный транскрипт полностью копирует молекулу ДНК.
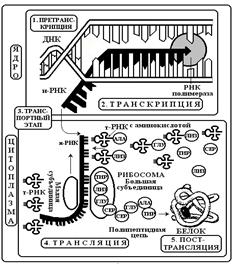 Рис. 13. Схема этапов синтеза белка
Рис. 13. Схема этапов синтеза белка
3. Транспортный уровень: охватывает период между транскрипцией и трансляцией. На данном этапе происходит процессинг, то есть созревание и-РНК. Суть его – удаление интронов. Экзоны сохраняются и соединяются в единую цепь с помощью ферментов лигаз. Указанное явление называется сплайсинг. В ходе его возможны ошибки транскрипции, когда удаляются не все интроны. Прошедшая сплайсинг и-РНК значительно укорочена, компактна и может служить матрицей для синтеза белка. Для этого она переносится из ядра в цитоплазму к рибосомам. В клетках животных и растений на всем пути от ядерного хроматина к рибосомам и-РНК находится не в свободном состоянии, а связана со специальными белками-переносчиками, часть из которых различна в ядре и цитоплазме. Белки «везут и-РНК в рибосому на перекладных».
4. Трансляционный уровень: Трансляция – синтез полипептидной цепи из аминокислот согласно кодирующей и-РНК. В ходе трансляции происходит перевод генетической информации в аминокислотную последовательность: ДНК, и-РНК, белок. Трансляция является очень важной частью общего метаболизма клетки. В ней задействованы не менее 20 ферментов (аминоацилсинтетаз), до 60 различных т-РНК, 3-5 молекул р-РНК и макомолекулы и-РНК. Здесь выделяют следующие этапы: инициация, элонгация, терминация.
Инициация – начало трансляции. Фермент аминоацилсинтетаза активирует определенную аминокислоту и вовлекает ее в белковый синтез. активированная аминокислота соединяется с т-РНК специализированной на данной аминокислоте. Доставленная к рибосоме и-РНК связывается с рибосомальной РНК. При этом рибосома диссоциирует, ее малая и большая субъединицы несколько раздвигаются и рибосома насаживается на и-РНК. В начальной части и-РНК имеются стартовые кодоны АУГ и ГУГ. К ним присоединяется т-РНК, несущая соответствующую аминокислоту формилметионин. Следовательно, эта аминокислота всегда является первой в синтезируемом на рибосоме белке.
Элонгация – продолжение трансляции. Рибосома двигается вдоль молекулы и-РНК. Транспортные РНК связываются с рибосомой и их антикодоны избирательно по правилу комплементарности контактируют с кодонами и-РНК. При совмещении кодона и антикодона аминокислота отрывается от т-РНК и включается в полипептидную цепь из аминокислот в большой субъединице рибосомы. При формировании первичной белковой структуры каждая новая аминокислота занимает место за аминокислотой, включенной непосредственно перед ней.
Терминация –окончание синтеза. В и-РНК есть трейлерный участок, содержащий стоп-кодоны УАА, УАГ,УГА. Они указывают на завершение синтеза данного белка. Поэтому при контакте с ними сборка аминокислотной цепи заканчивается
5. Посттрансляционный.уровень: синтезированаябелковая молекула начинает усложнять свою первичную структуру под действием ферментов. Происходит ее конформация, изменяется пространственная организация белка, он приобретает вторичную, третичную и четвертичную структуры.
Таким образом, мы познакомились с основными этапами синтеза белковой молекулы и узнали центральную догму молекулярной биологии. Она гласит: информация в живых организмах передается по цепи ДНК-РНК-белок. В настоящее время доказано явление обратной транскрипции, когда передача информации происходит от РНК к ДНК. В то же время совершенно невозможен перенос информации от белков обратно к нуклеиновым кислотам. Это означает, что изменения (модификации) белков, как конечных генных продуктов, не наследуются.
В соответствии с существующими представлениями синтез белка всегда начинается с работы генов. Под ней подразумевается их способность транскрибировать, то есть направлять синтез и-РНК. Но не всегда работа гена заканчивается сборкой белковой молекулы. Как заметил один американский генетику, для того, чтобы выдать белок гену нужно пробиться через «клеточные джунгли». Выход конечного белкового продукта – это экспрессия гена. Она совершается в результате деятельности всей клетки с ее многокомпонентными механизмами белкового синтеза.
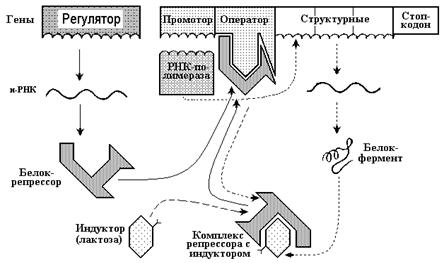
ОПЕРОННАЯ СИСТЕМА:
Гены функционально неоднородны. Это обнаружили в 1961 году французские ученые Ф. Жакоб и Ж. Моно. Они доказали, что существует две группы генов: структурные, управляющие синтезом специфических белков и регуляторные, контролирующие деятельность структурных генов. Синтез белка в клетке регулируется с помощью оперонной системы, состоящей из регуляторных (промотор и оператор) и структурных генов (рис. 14).
Следовательно, регуляция белкового синтеза в организме эукариот — процесс очень сложный. Он связан с наличием в клетке и с существованием, помимо генной, нервно-эндокринной системы регуляции. Открытия, сделанные в области молекулярной генетики, являются величайшими достижениями биологии.
Дата добавления: 2019-09-08; просмотров: 198; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!