Транскортикальная организация корковых нейронов.
Известно, что многие вставочные пирамидные нейроны имеют горизонтально направленные аксоны, а многие ассоциационные нейроны — длинные горизонтальные коллатерали. Это говорит за образование нейронных связей, а значит и нервных кругов в транскортикальном направлении. Немало физиологических фактов, говорящих за это, можно было бы привести, но мы приведем только некоторые новейшие факты.
Морелл (23) на кроликах и кошках подвергал двигательную зону одной лапы подпороговой анодной поляризации. Это приводило к повышению возбудимости в проекционных пирамидных нейронах этой лапы, без вызова
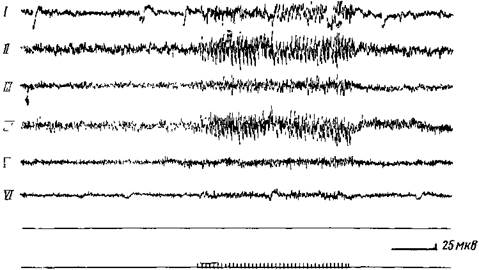
Рис. 60. Электрические ответы, зарегистрированные в различных слоях зрительной коры собаки на световое мелькание 10 в сек.
Биполярное отведение с межполюсным расстоянием 0,5 мм. Глубина погружения электродов от поверхности коры: слой 1—71,1 м к , II —213,0 м к , 117—426,0—368,2 мк; IV—651 мк; V —851,0—823,6 лиг; VI —1192,8—1107,6 мк (Рабинович)
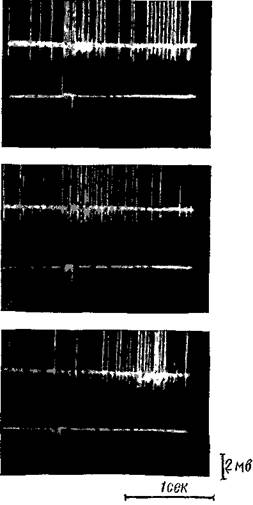
движения конечности. Из поляризованного участка микроэлектродами отводилась активность отдельных нейронов. Если во время поляризации раздражали кору электрическим током на расстоянии 1—2 см, то эти нейроны отвечали двойными вспышками пиковых потенциалов (рис. 62, а). Первая вспышка наступала со скрытым периодом в несколько мсек., а вторая — в несколько десятых секунды и, притом, тем позднее, чем дальше были раздражающие электроды. После перерезки коры вокруг участка, где находились поляризующий и отводящий электроды, без повреждения подкоркового белого вещества исчезала вторая вспышка (рис. 62, б), а после отделения этого участка коры от подкоркового белого вещества без кругового обреза исчезала первая вспышка (рис. 62, в). Очевидно, из раздражаемого участка возбуждение передавалось к поляризованному участку как по ассоциационным пирамидным нейронам посредством одной или двух синаптических передач, производя первую вспышку, так и по вставочным пирамидным нейронам посредством длинной серии синаптических передач, производя вторую вспышку.
Очевидные факты транскортикального распространения возбуждения даны в опытах Хананашвили (24, 25). Он наблюдал у собак, что после перерыва внутрикорковых связей, между отдельными участками затылочной области, происходит определенное нарушение зрительной функции. Именно, в первые дни после операции полностью отсутствует предметное зрение, со временем оно восстанавливается, но и через месяц оно осуществляется все еще труднее,
106

Рис. 61 Схема нейронных с в язей в з рительной вос п рин и мающей о б ла с ти.
Наверху дано схематичсское изображение 17 и 19 полей коры. Римскими цифрами обозначены слои коры (слева — дтя поля 19, справа — 17). Границы между полями обозначены пунктиром. Поле 18 на схеме не изображено
В средней части схемы даны подкорковые образования. Внизу — сетчатка глаза, С — звездчатые клетки Кахала с длинными аксонами, st — короткоаксонные звездчатые клетки, А ff 1 — специфические афференты из наружного коленчатого тела, А ff 2 — вертикальные афференты неизвестного происхождения, А ff 3 — ассоциационный афферент, М — пирамидная клетка Мейнерта, arc — пирамидная клетка с дугообразным аксоном, a — аксоны пирамидных клеток, P ul v — подушка зрительного бугра, cg l — наружное коленчатое тело, Pg n — прегеникулярное ядро, h — гипоталамус, Р r — протектуальное ядро; c q a — переднее двухолмие, п III — глазодвигательный нерв, c h — хиазма, R — сетчатка, g — ганглиозные клетки сетчатки, а1 — амакринные клетки, tr о — зрительный тракт Арабскими цифрами обозначены слои наружного коленчатого тела. Штриховые линии обозначают центрифугальную систему волокон, которые начинаются в коре и других отделах головного мозга и оканчиваются в сетчатке на амакринных клетках Схема Щкольник-Яррос составлена по ее собственным и литературным данным (Минковского, Ле Гро Кларка, Полякова, Кахала, Новохатского и др. )
|
|
чем до операции. Отсюда следует,, что транскортикальные связи играют определенную роль в предметном зрении. Но даже множественные насечки в зрительной или сенсомоторной коре, как это делали Сперри (26, 27) и Хананашвили (25), не приводят к существенному нарушению кожного, мышечного или зрительного восприятия, благодаря развитому вертикальному строению коры и свободной передаче возбуждения по ассоциационным пирамидным нейронам.
Транскаллозальные связи.
В процессе эволюционного развития большого мозга и неокортекса все большую роль приобретали межполушарные связи. У высших позвоночных эти комиссуральные волокна проходят через переднюю комиссуру и мозолистое тело (corpus callosum). В передней комиссуре имеются волокна, относящиеся к ольфакторным и лимбическим структурам. Межполушарные связи неокортекса определенным образом организованы, например, в сплениуме (splenium) проходят волокна зрительного различения, в задней части — тактильные, в колене (genu) и клюве (rostrum) проходят связи префронтальной коры.
| Рис. 62. Транскортикальное проведение возбуждения Двигательная зона коры кошки подвергается анодной поляризации. Отводятся пирамидные нейроны поляризованного участка. Раздражается кора электрическим токсм на расстоянии 1,5 см от поляризованного участка а — в ответ на раздражение возникают двойные вспышки пиковых потенциалов; б — после обреза коры вокруг поляризованного участка (перерезка транскортикальных путей) отводится лишь первая вспышка; в — после ^отделения коры от [подкоркового белого вещества без кругового [обреза (перерезка ассоциационных путей) в ответ на то же раздражение наступает только вторая вспышка (Моррел) |
Подробное изучение функций мозолистого тела было предпринято Сперри и сотр. (28). При целостности головного мозга у млекопитающих при обучении определенным движениям на показ фигуры через один глаз (другой глаз закрывался) этж же движения вызывались при предъявлении той же фигуры через другой глаз. Сперри и сотр. рассекали зрительную хиазму и мозолистое тело у кошек и обезьян и затем обучали определенным движениям на предъявление одной фигуры через один глаз, а другой фигуры через другой глаз. В этом случае, когда первую фигуру предъявляли через другой глаз, а вторую фигуру через первый, животные не реагировали правильно. Отсюда следует, что в нормальных условиях зрительное возбуждение одного полушария передается через мозолистое тело в другое полушарие.
То же самое наблюдали после перерезки мозолистого тела на голубях,, у которых зрительный нерв полностью перекрещивается (см. подробно в главе IX).
Помимо выяснения роли каллозальных связей в сопряженной работе левого и правого полушария, в последнее время прямо показана значительная роль этих волокон в передаче определенных афферентных сигналов к про-
108
|
|

тивоположному полушарию. Какова же «емкость» и каковы возможности в передаче информации по этим путям? Так, в опытах на кошках было показано, что после рассечения зрительной хиазмы и последующего раздельного обучения обоих полушарий только через один глаз, выполнение задачи на зрительное различение, проводящееся через «необученный» глаз (т. е. проверка, насколько информация о зрительном различении передалась по каллозальным путям), всегда менее эффективно, чем через афферентный путь
В случае решения противоположных задач, раздельно предъявляемых по прямой афферентной системе (когда глаз связан только с полушарием одноименной стороны) и по каллозальным путям (когда глаз связан с противоположным полушарием только через каллозальные пути), преобладание берет первое, т. е. выполняется задача, предъявленная по прямому афферентному пути. Это касается не только зрительного, но и тактильного различения (Мийерс, 29).
| Рис. 63, Распределение комиссуральных волокон по поверхности левого полушария у кошки I— медиальная поверхность, II — вид сверху, III — вид сбоку (Эбнер и Мийерс) |
|
|

Электрический анализ показал, что при непосредственном раздражении волокон мозолистого тела и отведении вызванных потенциалов от поверхности коры (например, в супрасильвиевой извилине), транскаллозальные вызванные потенциалы имеют специфические особенности. Они возникают в иных структурах, нежели вызванные потенциалы на афферентное прямое раздражение коры (Фешер и др., 30).
| Рис. 64. Распределение комиссуральных волокон по поверхности левого полушария у обезьян (Масаса mulatt o ) Определение велось по методу Наута после полной перерезки передней комиссуры и мозолистого тела (Мийерс) |
Морфологические транскаллозальные связи изучены в ряде лабораторий. В последнее время исследовано распределение дегенерирующих волокон в коре кошки, енота и обезьяны после перерезок мозолистого тела и передней комиссуры (Эбнер и Мийерс, 31). Так, у кошки, как представлено на рис. 63, /—///, отчетливо видны зоны, свободные от комиссуральных связей. Однако, как отмечает и сам Мийерс, еще нет полной ясности в отношении связи между функциональной деятельностью и наличием или отсутствием комиссуральных волокон в данных областях левого и правого полушарий. У обезь-
109
яны после полной перерезки комиссур дегенерация волокон наблюдается шире, чем у кошки. У обезьяны после полной перерезки комиссур в затылочных долях наблюдается такая же картина дегенерации, как и после удаления только затылочных долей противоположного полушария (см. рис. 64). Отчетливо показано отсутствие комиссуральных связей в передней трети височной доли. На приводимых рисунках видно, что отсутствуют связи в пределах 1,2 и 3 поля (представительство верхних конечностай) и в передней части теменной доли. Особенно богаты комиссуральными связями задняя теменная и прецентральная области (Мийерс, 29).
Дата добавления: 2019-07-17; просмотров: 388; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!