Методы выделения чистой культуры анаэробов
1. Метод Цейсселера. Исследуемый материал засевают штрихами последовательно на три чашки Петри по поверхности плотной питательной среды, помещают в анаэробные условия и культивируют в термостате при 37°С в течение 24-72 ч. Изолированные колонии анаэробов пересевают в среду для контроля стерильности (СКС) или среду Китта-Тароцци.
2. Метод Вейнберга. Несколько капель исследуемого материала вносят в пробирку с 4-5 мл раствора хлористого натрия, перемешивают и переносят в пробирку с охлажденным до 45-50°С полужидким сахарным агаром, разлитым высоким столбиком. После перемешивания последовательно засевают еще в четыре пробирки с сахарным агаром и быстро охлаждают под струей холодной воды. Выросшие через 24-72 ч в глубине агара изолированные колонии анаэробов засевают в среду Китта-Тароцци.
Многократный перенос петли с материалом по вышеуказанным методам повышает степень разобщения микробов и вероятность роста их колоний в последней пробирке или чашке Петри.
3. Метод Перетца. Готовят разведения исследуемого материала как указано выше. Содержимое пробирки с соответствующим разведением выливают в стерильную чашку Петри, на дне которой на двух стеклянных или деревянных палочках лежит стеклянная пластинка размером 6х6 см. Среду заливают таким образом, чтобы она заполнила пространство между пластинкой и дном чашки Петри. При появлении микробного роста стеклянную пластинку удаляют, а изолированные колонии засевают в пробирку со средой Китта-Тароцци для получения чистой культуры.
|
|
|
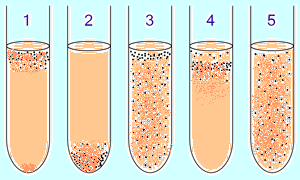
| Рис. 8.2. Рост микробов в жидкой питательной среде в зависимости от отношения к кислороду |
Аэробные и анаэробные бактерии предварительно идентифицируются в жидкой питательной среде по градиенту концентрации O2 (рис. 1):
1. Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать максимальное количество кислорода (исключение: микобактерии – рост пленкой на поверхности из-за восколипидной мембраны).
2. Облигатные анаэробные бактерии собираются в нижней части, чтобы избежать кислорода (либо не дают роста).
3. Факультативные бактерии собираются в основном в верхнем (окислительное фосфорилирование является наиболее выгодным, чем гликолиз), однако они могут быть найдены на всем протяжении среды, так как от не зависят O2.
4. Микроаэрофилы собираются в верхней части пробирки, но их оптимум — малая концентрация кислорода.
5. Аэротолерантные анаэробы не реагируют на концентрации кислорода и равномерно распределяются по пробирке.
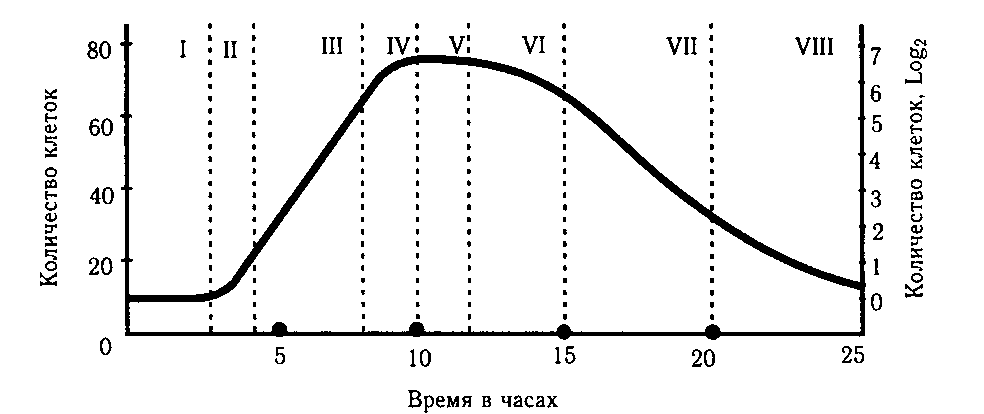
Рис.8.3. Стадии роста периодической микробной культуры.
|
|
|
I —исходная стационарная фаза; II — лаг-фаза (фаза задержки размножения); III — фаза логарифмического роста; IV — фаза отрицательного ускорения; V — стационарная фаза; VI — фаза ускоренной гибели; VII — фаза логарифмической гибели; VIII — фаза уменьшения скорости гибели. На оси ординат показаны скорость размножения бактерий в условных единицах (слева) и величина популяции бактерий, выраженная логарифмом от числа живых клеток на 1 мл среды.
Фазы размножения бактерий во времени зависят от вида бактерий, качества питательной среды, ее концентрации, температуры и аэрации. Поэтому продолжительность каждой фазы указана ориентировочно. При оптимальных условиях деление клетки у ряда особей происходит в разное время, например, кишечные палочки делятся через 15 - 20 минут, брюшнотифозные бактерии - 20 - 25 минут, стрептококки - 25 - 30 минут, микобактерии туберкулеза - 18 - 20 часов.
Длительность фазы отмирания клеток связана с видовыми особенностями бактерий. Период отмирания пневмококка длится 2 - 3 дня, а кишечной палочки - месяцы. В стадии отмирания клетки слабо окрашиваются, а некоторые из них не воспринимают краски. Кроме того, изменяются формы бактерий, их биохимическая активность и антигенные свойства.
|
|
|
Сходство процессов брожения и дыхания.
1) Оба процесса (дыхание и брожение) — окислительно-восстановительные, и это определяет их сходство. И при брожении, и при дыхании образуется биологически доступная энергия в форме АТФ.
2) И при брожении, и при дыхании происходит гликолиз — анаэроб-ная стадия окисления химических соединений с извлечением энергии за счет идущего в цитоплазме субстратного фосфорилирования.
3) И в процессе брожения, и в процессе дыхания участвуют ферменты оксидоредуктазы: дегидрогеназы — НАД-дегидрогеназы, или никотин-амидадениндинуклеотиддегидрогеназы, отнимающие водород от субстратов при субстратном фосфорилировании.
Бродильный (ферментативный) метаболизм.
Ферментация, или брожение, — процесс получения энергии, при котором отщепленный от субстрата водород переносится на органические соединения. Кислород в процессе брожения участия не принимает. Восстановленные органические соединения выделяются в питательную среду и накапливаются в ней. Ферментироваться могут углеводы, аминокислоты (за исключением ароматических), пурины, пиримидины, многоатомные спирты. Не способны сбраживаться ароматические углеводороды, стероиды, каротиноиды, жирные кислоты. Эти вещества разлагаются и окисляются только в присутствии кислорода, в анаэробных условиях они стабильны. Продуктами брожения являются кислоты, газы, спирты.
|
|
|
При ферментации гексоз (глюкозы) пируват лишь частично окисляется в цикле трикарбоновых кислот. Последний выполняет только функции поставщика предшественников для биосинтетических процессов. Энергия в форме двух молекул АТФ образуется в результате субстратного фосфорилирования, протекающего при окислении триозофосфата в пируват. Отщепившийся от субстрата водород, находящийся в форме восстановленного НАД, переносится на пируват, превращая его в цепи реакций в этанол, кислоты, газы. Исходя из природы конечных продуктов, различают несколько типов брожения углеводов: спиртовое, молочнокислое, муравьинокислое, маслянокислое.
Спиртовое брожение встречается в основном у дрожжей. Конечными продуктами являются этанол и С02. Спиртовое брожение используется в хлебопекарной промышленности и виноделии.
Молочнокислое брожение происходит у S. pyogenes, Е. faecalis, S. Salivarius, а также у бактерий родов Lactobacillus и Bifidobacterium. Продуктами этого типа брожения являются молочная кислота, этанол и уксусная кислота. Продукты молочнокислого брожения играют большую роль в формировании колонизационной резистентности бактериями рода Lactobacillus и Bifidobacterium, составляющих облигатную флору кишечника. Молочнокислые бактерии широко используются в молочной промышленности для получения молочнокислых продуктов, а также в создании пробиотиков.
Муравьинокислое (смешанное) брожение встречается у представителей семейств Enterobacteriaceae и Vibrionaceae. Различают два типа этого брожения. При первом происходит расщепление пи- рувата с образованием через цепь реакций муравьиной, янтарной и молочной кислот. Сильное кислотообразование можно выявить реакцией с индикатором метиленовым красным, который меняет окраску в сильнокислой среде. При втором типе брожения образуется целый ряд кислот, однако главным продуктом брожения являются ацетоин и 2,3-бутандиол, образующиеся через цепь реакций из двух молекул пирувата. Эти вещества при взаимодействии с а-нафтолом в щелочной среде вызывают образование окраски бурого цвета, что выявляется реакцией Фогеса—Проскауэра, используемой при идентификации бактерий.
Маслянокислое брожение. Масляная кислота, бутанол, ацетон, изопропанол и ряд других органических кислот, в частности уксусная, капроновая, валериановая, пальмитиновая, являются продуктами сбраживания углеводов сахаролитическими строгими анаэробами. Спектр этих кислот, определяемый при помощи газожидкостной хроматографии, используется как экспресс-метод при идентификации анаэробов.
Ферментация белков. Если для бактерий с бродильным метаболизмом источником энергии служат белки, то такие бактерии называются пептолитическими. Пептолитическими являются некоторые клостридии, в частности С. histolyticum и С. botulinum. Пептолитические бактерии гидролизуют белки и сбраживают аминокислоты. Многие аминокислоты сбраживаются совместно с другими, при этом одна выполняет функцию донора, а другая — функцию акцептора водорода. Аминокислота-донор дезаминируется в кетокислоту, которая в результате окислительного декарбок- силирования превращается в жирную кислоту.
Дата добавления: 2019-02-22; просмотров: 1040; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!