ФИЗИОЛОГИЯ ЛИМБИЧЕСКОИ СИСТЕМЫ
Лимбическая система представляет собой функциональное объединение структур мозга,
участвующих в организации эмоционально-мотивационного поведения, таких как пищевой, половой, оборонительный инстинкты. Все основные формирования лимбической коры кольцеобразно охватывают основание переднего мозга и являются своеобразной границей между новой корой и стволовой частью мозга.
Морфофункциональная организация. Структуры лимбической системы включают в себя 3 комплекса. Первый комплекс — древняя кора: препериформная, периамигдалярная, диагональная кора, обонятельный мозг (обонятельные луковицы, обонятельный бугорок, прозрачная перегородка).

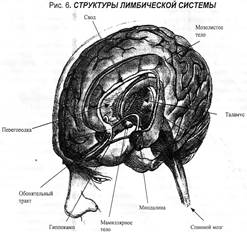
Вторым комплексом структур лимбической системы является старая кора, куда входят гиппокамп, зубчатая фасция, поясная извилина.
Третий комплекс лимбической системы: структуры островковой коры, парагиппокамповая извилина.
И, наконец, в лимбическую систему включают подкорковые структуры: миндалевидные тела, ядра прозрачной перегородки, передние ядра таламуса, сосцевидные тела, гипоталамус.
Связи в лимбической системе. Особенностью лимбической системы является наличие множественных связей как между отдельными структурами этой системы, так между лимбической системой и другими структурами мозга, по которым информация может длительно циркулировать. Благодаря таким особенностям создаются условия для эффективного управления структурами мозга со стороны лимбической системы («навязывание» лимбического влияния).
|
|
|
Лимбическая система имеет:
Ø восходящие пути, связывающие ее с новой корой, например, миндалины связаны с височными отделами коры;
Ø нисходящие пути, связывающие ее с гипоталамусом, ретикулярной формацией среднего мозга, со стволовыми центрами;
Ø проходящие пути в составе медиального пучка переднего мозга, конечной полоски и свода.
В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную специфику. К ним относится круг Пейпеса (гиппокамп → сосцевидные тела → передние ядра таламуса → поясная извилина →парагиппокамповая извилина → гиппокамп). Этот круг имеет отношение к памяти и процессам обучения.
Другой круг (миндалевидное тело → гипоталамус → структуры среднего мозга → миндалевидное тело) регулирует агрессивно-оборонительное поведение, а также пищевые и сексуальные формы поведения.
Круги разного функционального назначения связывают лимбическую систему со многими структурами центральной нервной системы, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой. Например, образная (иконическая) память формируется кортико-лимбико-таламо-кортикальным кругом, а круг, соединяющий новую кору и лимбическую систему через хвостатое ядро, имеет прямое отношение к организации тормозных процессов в коре больших полушарий.
|
|
|
Функции лимбической системы. Большое количество связей в лимбической системе, своеобразное круговое взаимодействие ее структур создают благоприятные условия для реверберации возбуждения по коротким и длинным кругам. Это, с одной стороны, обеспечивает функциональное взаимодействие частей лимбической системы, с другой — создает условия для запоминания.
За счет обилия связей внутри лимбической системы, а также ее обширных связей с другими структурами мозга, эта система выполняет достаточно широкий спектр функций. Среди них можно выделить такие как:
1)регуляция функций диэнцефальных и неокортикальных образований: формирование отношений прошлого, настоящего и будущего; обеспечение адекватного поведения в обстановке.
2)создание эмоционального фона. Согласно теории Пейпеса сенсорные пути на уровне таламуса расходятся, причём один путь идёт в проекционные зоны коры, где обеспечивается восприятие, а второй - в лимбическую систему. Вероятно, в этой системе происходит оценка поступающей информации, её сопоставление с субъективным опытом и запуск соответствующих эмоциональных реакций. В развитии эмоции участвуют все элементы лимбической системы. Подкрепляющим эффектом (положительные эмоции) обладают катехолергические и дофаминергические
|
|
|
структуры центра «удовольствия, вознаграждения», расположенного в медиальном пучке переднего мозга. Эффектом избегания (отрицательные эмоции) обладают перивентрикулярные отделы промежуточного и среднего мозга - центры «неудовольствия, избегания». Эта область значительно меньше предыдущей. Обе области частично перекрываются.
3)регуляция вегетативных и соматических процессов при эмоционально-мотивационной деятельности. Некоторые авторы называют лимбическую систему висцеральным мозгом, т.е. структурой ЦНС, участвующей в регуляции деятельности внутренних органов: при возбуждении миндалевидных тел, прозрачной перегородки, обонятельного мозга изменяется активность вегетативной системы в соответствии с условиями окружающей среды. Лимбическая система является высшим регулятором вегетативной нервной системы, осуществляя контроль над гипоталамусом, который является ее эфферентным звеном. Изменяя в ту или иную сторону возбудимость гипота-
ламических центров, лимбическая система определяет знак соответствующей вегетативной реакции. Так формируется иерархическая система управления вегетативной сферой, интегрирующая вегетативные и соматические реакции.
|
|
|
При раздражении лимбической системы наблюдается саливация, изменение потоотделения, функций сердечно-сосудистой и дыхательной систем. Отличительной особенность этих вегетативных реакций является то, что они сопровождаются координированными моторными реакциями (например, жевание, глотание). Кроме того, лимбическая система обеспечивает позно-тонические реакции при эмоциональных состояниях (устрашающие позы при гневе и т.д.).
Как известно, эмоциональная окраска поведенческих реакций определяется также соответствующими эндокринными сдвигами. Например, нисходящие влияния ядер миндалевидного комплекса и гиппокампа на гипоталамус, а через него на гипофиз изменяют продукцию кортикотропина, который регулирует секрецию кортикостероидов. Таким образом, изменяя гормональный фон, лимбическая система в естественных условиях может участвовать в формировании мотиваций (побуждений к действию) и регулировать реализацию действий, направленных на устранение
побуждения.
4)регуляция уровня внимания, восприятия, памяти и воспроизведения эмоционально значимой информации. Механизмы долговременной памяти связаны с гиппокампом, т.к. в нем наблюдаются круги реверберации. Ему и миндалине отводится большая роль в механизме обучения. Двустороннее удаление миндалины вызывает потерю памяти (амнезию).
В гиппокампе находятся нейроны, которые реагируют на появление каждого нового стимула, т.е. участвуют в ориентировочном рефлексе, реакциях настороженности, повышения внимания, в динамике обучения. Активация нейронов гиппокампа наблюдается при высоком уровне эмоционального напряжения: страхе, агрессии, голоде, жажде. Также в гиппокампе происходит отбор существенной информации от несущественной.
5)выбор и реализация адаптивных форм поведения, включая такие виды поведения как поисковое, пищевое, половое, оборонительное, а также регуляция динамики врожденных форм поведения;
6)участие в организации цикла «сон — бодрствование». Лимбическая система как филогенетически древнее образование оказывает регулирующее влияние на кору большого мозга и подкорковые структуры, устанавливая необходимое соответствие уровней их активности. Холинергические структуры лимбической системы образуют гипногенный лимбико-среднемозговой путь. Возбуждение структур этого круга вызывает блокаду всех активирующих влияний ретикулярной формации среднего мозга на кору больших полушарий. Таким образом между ретикулярной формацией и лимбической системой существуют реципрокные взаимоотношения.
Несомненно, что важную роль в реализации всех перечисленных функций лимбической системы играет поступление в эту систему мозга информации от обонятельных рецепторов и ее обработка. Обонятельный анализатор, как самый древний из анализаторов, является неспецифическим активатором всех видов деятельности коры большого мозга.
МЕДИАТОРЫ АВТОНОМНОЙ (ВЕГЕТАТИВНОЙ) НЕРВНОЙ СИСТЕМЫ
Ацетилхолин высвобождается в окончаниях всех преганглионарных (и симпатических, и парасимпатических) нейронов и большинства постганглионарных парасимпатических нейронов. Кроме того, часть постганглионарных симпатических нейронов, идущих к потовым железам, и симпатические нейроны, вызывающие расширение сосудов скелетных мышц, также осуществляют передачу с помощью ацетилхолина.
Никотин вызывает подобный ацетилхолину эффект при действии на постсинаптическую мембрану постганглионарного нейрона, сложные эфиры холина, пилокарпин и токсин мухомора мускарин — на мембрану эффекторного органа. Такое различие в вызываемых реакциях послужило основанием к выделению двух типов холинорецепторов: никотинового (Н- холинорецепторы - находятся на постсинаптической мембране постганглионарного нейрона,) и мускаринового (М-холинорецепторы - на постсинаптической мембране органов-эффекторов/
Норадреналин является медиатором в постганглионарных симпатических окончаниях сосудов сердца, печени, селезенки.
Попадая на постсинаптическую мембрану, норадреналин взаимодействует с ее
специфическими химическими компонентами — адренорецепторами. В соответствии с
чувствительностью к различным катехоламинам (обычно к адреналину, норадреналину,
изопротеренолу) адренорецепторы делят на а-адренорецепторы (более чувствительны к норадреналину) и ft -адренорецепторы (более чувствительны к адреналину).
В обычных жизненных условиях реакция органа на адреналин и норадреналин,
выделяющиеся нервными окончаниями или приносимые кровью, зависит от преобладания а - или ft -адренергического действия.
Серотонин: в мозге серотонин содержится главным образом в структурах, имеющих отношение к регуляции висцеральных органов. Особенно много его в лимбической системе, ядрах шва. Аксоны нейронов, богатых серотонином проходят в бульбоспинальных путях и оканчиваются в сегментах спинного мозга. Здесь они контактируют с клетками преганглионарных симпатических нейронов и со вставочными нейронами студенистого вещества. Фармакологически установлено существование трех типов серотонинергических рецепторов: D, М, Т.
D -серотонинергические рецепторы локализуются в основном в гладкой мышце.
Взаимодействие серотонина с этими рецепторами сопровождается сокращением гладких мышц. М-серотонинергические рецепторы расположены в основном в ганглиях автономной нервной системы. Влияя на эти рецепторы, серотонин вызывает ганглиостимулирующий эффект. Т- серотонинергические рецепторы обнаружены в сердечной и легочной рефлексогенных зонах. Действуя на них, серотонин вызывает коронарный и легочный хеморефлексы.
Аденозинтрифосфат (АТФ) является синаптическим передатчиком, широко
представленным в различных органах и особенно в эффекторных нейронах метасимпатической нервной системы, где локализуется в пресинаптических терминалях. В связи с тем, что при стимуляции этих терминалей выделяются пуриновые продукты распада — аденозин и инозин, эта передача получила название пуртергической.
Пуринергические нейроны являются главной антагонистической тормозной системой по отношению к холинергической возбуждающей системе в механизме кишечной пропульсии. Они участвуют в нисходящем» торможении. Сокращения кишки, следующие за возникающим в результате активации пуринергических структур расслаблением, обеспечивают соответствующий механизм для прохождения пищевого комка (болюса) по кишке. Пуринергические нейроны участвуют также в механизме рецептивной релаксации желудка, расслаблении пищевого и анального сфинктеров.
Реакции гладкомышечных органов на АТФ опосредованы его взаимодействием с
пуринорецепторами клеточных мембран. На основании фармакологических критериев выделено два их типа. Пуринорецепторы первого типа более чувствительны к продукту распада АТФ — аденозину, второго — к самому АТФ.
Трансдукторы. Для выполнения своих функций и поддержания гомеостаза автономная нервная система наряду с обычными нейронами содержит особые клетки, прием информации в которых осуществляется обычным путем, а ответы — эндокринным способом. Такие клетки называются трансдукторами.
К трансдукторам относят:
1) хромаффинные клетки мозгового слоя надпочечников, которые отвечают на холинергическии передатчик преганглионарного симпатического волокна выделением адреналина и норадреналина;
2) юкстагломерулярные клетки почки, которые отвечают на адренергический передатчик постганглионарного симпатического волокна выделением в кровяное русло ренина;
3) нейроны супраоптического и паравентрикулярного ядер гипоталамуса, реагирующие на адренергический, холинергический и другие передатчики выделением вазопрессина и окситоцина;
4) нейроны ядер гипоталамуса, выделяющие в сосудистую систему факторы регуляции.
Рис. 7. ПРИНЦИПИАЛЬНАЯ СХЕМА ДУГИ АВТОНОМНОГО (СПРАВА) И
СОМАТИЧЕСКОГО (СЛЕВА) РЕФЛЕКСОВ (ПО А. Д. НОЗДРАЧЕВУ)

1 — спинной мозг;
2 — тело чувствительного нейрона;
3 — спинальный ганглий;
4 — задний корешок;
5 — вставочный (преганглионарный) нейрон дуги
автономного рефлекса;
6 - передний корешок;
7 — преганглионарное волокно;
8 — околопозвоночный узел;
9 — белая соединительная ветвь;
10 — двигательный (постганглионарный) нейрон дуги
автономного рефлекса;
11 - соматический нерв;
12 — постганглионарное волокно;
13 — серая соединительная ветвь;
14 — висцеральная ветвь;
15 — предпозвоночный узел (брыжеечного, чревного или солнечного сплетения);
16 — кишка;
17 — эффекторный нейрон функционального модуля
метасимпатической нервной системы;
18 — функциональный модуль
метасимпатической нервной системы;
19 — интернейрон функционального
модуля;
20 — чувствительный нейрон
функционального модуля;
21 —двигательное соматическое волокно
22— двигательный нейрон соматической
рефлекторной дуги;
23 — вставочный нейрон соматической рефлекторной дуги.
СТЕРЕОТАКСИЧЕСКАЯ ТЕХНИКА
Стереотаксическая техника широко применяется для введения в различные подкорковые структуры стимулирующих и отводящих подкорковых электродов и микропипеток. Для точного попадания в исследуемые подкорковые ядра в экспериментах используют животных, имеющих средние размеры черепа.
Стереотаксический прибор состоит из специальных держателей, с помощью которых голова фиксируется всегда в строго определенном положении относительно рамы стереотаксиса. Ушные держатели вводят в наружные слуховые проходы. Снизу голову животного фиксируют подведением пластин под верхнюю челюсть. Сверху закрепляют глазные держатели, давящие на нижние края орбит.
Таким образом, голова животного, а следовательно, и мозг оказываются ориентированными в постоянной координатной системе, где воображаемая горизонтальная плоскость проходит по нижним краям орбит и наружным слуховым проходам. Перпендикулярно этой плоскости располагается сагиттальная плоскость, проходящая по межполушарной борозде и средней точке линии ушных держателей. Третья плоскость — фронтальная — перпендикулярна выше описанным плоскостям и проходит по линии ушных держателей. Центральной (нулевой) точкой является точка соприкосновения ушных держателей. Для человека разработаны значительно более
сложные стереотаксические приборы. Для точного определения координат подкорковых структур созданы стереотаксические атласы мозга различных животных. Широко применяется во многих лабораториях мира стереотаксический атлас кошки, составленный X. Джаспером и К. Аймон-Марсаном (рис. 8), а также атлас. К. Р. Хименес-Кастелланоса, который позволяет представить взаиморасположение подкорковых таламических структур (Рис.9).

Рис. 8. Стереотаксическая установка для проведения нейрохирургических операций на мозге человека

Рис.9. Фронтальный разрез мозга кошки с осями координат и буквенными обозна-чениями подкорковых структур мозга (из атласа Джаспера и Айрон-Марсана) (фронтальный план 13,0 мм).
Особое значение приобретают стереотаксическая техника и метод электрофизиологического контроля для нейрохирургической клиники. Известно, что целый ряд тяжелых хронических заболеваний ЦНС поддается только хирургическому лечению, которое сводится к локальному разрушению определенных подкорковых ядер. В связи с этим появилась новая дисциплина — «стереотаксическая нейрохирургия». Применение стереотаксической техники невозможно без функционального электрофизиологического контроля, поскольку ни рентгеновский контроль, ни
специальные таблицы, ни сложные математические расчеты не обеспечивают абсолютной точности при введении подкорковых электродов в мозг человека, отсюда вытекает необходимость выработки четких электрофизиологических критериев для определения функциональных особенностей тех или иных центров, расположенных в глубине мозга.
ЭЛЕКТРОЭНЦЕФАЛОГРАФИЯ (ЭЭГ)
Спонтанная электрическая активность мозга характеризуется специфическими ритмами
определенной частоты и амплитуды и может быть записана одновременно от многих участков черепа. Это позволяет изучать пространственные специфические паттерны ЭЭГ. ЭЭГ представляет собой низкоамплитудные колебания электрической активности, частотные и амплитудные характеристики которых зависят от состояния ЦНС. .

Регистрацию ЭЭГ проводят в специальной экранированной
камере, предохраняющей (экранирующей) объект от воздействия внешних электрических и магнитных полей. Электрическую активность мозга отводят с помощью специального электродного устройства, которое состоит из отводящих электродов и держателя электродов, выполненного обычно в виде шлема. ЭЭГ отражает колебания во времени разности потенциалов между двумя электродами. Для расположения электродов используют международную схему «10—20». Отведение маркируют буквой, указывающей
на область мозга, от которой оно производится, — F, О, Т, Р, С. Электроды должны иметь надежный контакт с кожей.
При записи ЭЭГ используют разного рода воздействиях на испытуемого: 1. При расслаблении мышц и закрытии глаз спустя несколько минут наблюдают появление на ЭЭГ альфа-ритма. При открытии глаз наблюдают депрессию альфа-ритма. 2. Проба с
фотостимуляцией проводится при частоте подачи световых сигналов 20 Гц. Включают электроэнцефалограф и записывают исходный фон, затем включают световой раздражитель и через 1— 2 мин регистрируют ЭЭГ. В этом случае колебания потенциала на ЭЭГ начинают повторять частоту следования светового раздражителя. Это явление носит название «усвоение ритма» раздражения.
Существует определенная классификация различных видов отведений, используемых в ЭЭГ .
Ритмы ЭЭГ
Альфа-ритм с частотой 8-13 Гц и амплитудой 5-100 мкВ регистрируется преимущественно в затылочной и теменной областях. Наблюдается в состоянии спокойного бодрствования, медитации при закрытых глазах (если глаза открыты, он обычно исчезает, заменяясь более быстрым бетта- ритмом). Часто на ЭЭГ видно, что его амплитуда постепенно увеличивается, а затем уменьшается. По данным некоторых авторов у слепых с врожденной или многолетней патологией, а также при сохранности только светоощущения альфа-ритм отсутствует. Исчезновение альфа-ритма наблюдается и при атрофии зрительного нерва. Предполагается у них, что альфа-ритм совпадает с наличием предметного зрения.
Бетта-ритм имеет частоту 18-30 Гц и амплитуду колебаний 2-20 мкВ. Его локализация в прецентральной и фронтальной коре. Резко усиливается при различных видах интенсивной деятельности

Рис.11. Схема стандартного расположения электродов на голове человека в соответствии с международной системой отведения ЭЭГ «10-20» (по Н. Jasper, 1958).
а — вид спереди, б — вид сверху, в — вид сбоку.
Гамма-колебания охватывают частоты от 30-120-170 Гц, а по данным нескольких авторов до 500 Гц, при их амплитуде 2 мкВ. Их можно наблюдать в прецентральной, фронтальной, височной, теменной зонах коры. Наблюдается при решении задач, требующих максимального сосредоточения.
Дельта-волны возникают в диапазоне 0,5-4,0 Гц (20-200мкВ), зона их появления варьирует. Возникает при естественном и наркотическом сне, а также при регистрации ЭЭГ от участков коры, граничащих с опухолью.
Тета-волны имеют частоту 4-7 Гц (5-100 мкВ) и чаще наблюдаются во фронтальных зонах и гиппокампе. Связан с поисковым поведением, усиливается при эмоциональном поведении.
Каппа-колебания на частоте 8-12 Гц (5-40 мкВ), сходен по частоте с альфа-ритмом, можно увидеть в височной области в процессе умственной деятельности.

В целом, ЭЭГ позволяет определить характер состояния мозга (активный, бодрствующий или спящий мозг), стадии естественного сна, позволяет выяснить так называемый парадоксальный сон, она даст возможность судить о глубине наркоза, о наличии патологического очага в мозге (эпилептический очаг, опухоль) и т. д.
Характер электроэнцефалограммы (ЭЭГ) определяется функциональным состоянием нервной ткани, уровнем протекающих в ней обменных процессов. Нарушение кровоснабжения, гипоксия или глубокий наркоз приводят к подавлению биоэлектрической активности коры больших полушарий. Зависимость ЭЭГ от общего состояния организма широко используют в клинике для контроля за ходом операции и уровнем наркоза.
Дата добавления: 2019-02-12; просмотров: 3229; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!