Нейрофизиологические особенности клеток коры мозжечка
Корзинчатые и звездчатые клетки, которые заканчиваются синапсами на клетках Пуркинье, вызывают в них тормозные постсинаптические потенциалы (ТПСП) и подавление импульсной активности. Клетки Гольджи тормозят клетки-зерна по принципу обратной связи. Таким образом, большинство связей, опосредованных интернейронами коры мозжечка, являются тормозными. Исключение составляют только клетки-зерна, которые возбуждаются от моховидных волокон и сами через Т-образно ветвящиеся аксоны активируют все остальные интернейроны коры мозжечка. Однако конечный эффект этой активации опять-таки сводится к торможению.
Клетки Пуркинье, которые представляют собой выход функциональной системы, могут возбуждаться прямо через лазащие волокна и опосредованно через моховидные волокна и клетки- зерна. Возникающие под действием этого возбуждения разряды клеток Пуркинье, согласно электрофизиологическим данным, вызывают в конечном счете торможение нейронов ядер мозжечка. Эти факты свидетельствуют о том, что деятельность всей нейрональной системы коры мозжечка сводится к торможению ядер, над которыми кора надстроена. Сами же ядра мозжечка, обладающие постоянной тонической активностью, через нисходящие пути регулируют уровень возбудимости центров спинного мозга и мышечный тонус.
В белом веществе мозжечка сконцентрированы три пары ядер:
1. В белом веществе червя близко к срединной плоскости находится ядро шатра. Нейроны этого ядра посылают свои отростки к латеральному преддверному ядру (ЛПЯ) и к ретикулярной формации продолговатого мозга и моста, где берет свое начало ретикулярно-спинномозговой путь.
|
|
|
2. Латеральнее ядра шатра находятся вставочные, или промежуточные, ядра, которые разделяется на латеральное (пробковидное) и медиальное (шаровидное) вставочные ядра мозжечка. От вставочного ядра аксоны идут в средний мозг к красному ядру. Менее развитый афферентный путь от вставочного ядра идет в проме-жуточный мозг к вентролатеральному ядру таламуса — и оттуда к двигательной коре.
3. Латеральное всех ядер лежит наиболее крупное латеральное (зубчатое) ядро мозжечка, от которого мощные пучки волокон направляются к вентролатеральному ядру таламуса, и далее аксоны нейронов второго порядка проецируются в моторные зоны коры.
Функции мозжечка
Мозжечок как надсегментарный орган, входящий в систему регуляции движений, выполняет следующие важные функции:
1) регуляция позы и мышечного тонуса;
2) сенсомоторная координация позных и целенаправленных движений;
3) координация быстрых целенаправленных движений, осуществляемых по команде из
больших полушарий.
|
|
|
Эти функции мозжечка наиболее удобно разбирать в соответствии с топической
классификацей его отделов, основанной на характере эфферентных связей.
Медиальная червячная зона мозжечка в наибольшей степени связана с управлением
тонусом, позой и равновесием тела. В эту зону коры мозжечка поступает афферентная
информация, сигнализирующая о позе и состоянии локомоторного аппарата. После обработки этой информации из коры мозжечка через ядро шатра корригирующие команды направляются к ЛПЯ, к ретикулярной формации ствола и оттуда к спинальным центрам по ретикулярно- и преддверно-спинномозговым путям.
Промежуточная зона коры мозжечка, связанная со вставочным ядром (или шаровидным и пробковидным), так же как и предыдущая зона, имеет афферентные входы от спинномозговых путей. Однако кроме того, через ядра моста промежуточная зона коры мозжечка получает информацию от двигательной области коры больших полушарий. Эта информация поступает через коллатерали корково-спинномозгового пути и сигнализирует о готовящемся целенаправленном движении. Сопоставление приходящей по этим двум путям информации позволяет промежуточной зоне мозжечка участвовать в координации целенаправленных движений с рефлексами поддержания позы, в выборе позы оптимальной для выполнения движения. Нисходящие команды от промежуточной зоны коры мозжечка через вставочное ядро идут к красному ядру и далее по красноядерно-слинномозговому пути к моторным центрам спинного мозга.
|
|
|
Латеральная зона через корково-мосто-мозжечкодый путь получает афферентный вход от различных ассоциативных зон коры больших полушарий. По этим афферентным путям в кору полушарий мозжечка поступает информация о замысле движения. В полушариях и зубчатом ядре мозжечка эта информация преобразуется в программу движения, которая по мозжечково-таламо-корковому пути поступает в двигательные зоны коры больших полушарий. В дальнейшем двигательный акт реализуется за счет нисходящих команд, идущих от двигательной коры в спинной мозг по корково-спинномозговому пути. Кроме того, от зубчатого ядра мозжечка через
красное ядро могут распространяться прямые нисходящие воздействия к спинальным центрам.
Эта сложная система регуляции движений с наличием обратных связей между мозжечком и корой больших полушарий позволяет полушариям мозжечка участвовать в организации быстрых целенаправленных движений, протекающих без учета информации, идущей от восходящих сенсорных путей спинного мозга. Такие движения могут встретиться в спортивной практике, при игре на музыкальных инструментах и при некоторых других видах деятельности.
|
|
|
Экспериментальные исследования функций мозжечка показали, что этот отдел головного мозга участвует не только в регуляции движений, но и включен в систему контроля висцеральных функций. Л. А. Орбели было установлено, что раздражение мозжечка вызывает целый ряд вегетативных рефлексов, таких, например, как расширение зрачков, повышение артериального давления и т. д. Удалению мозжечка сопутствуют нарушения сердечно-сосудистой деятельности, дыхания, моторной и секреторной функций желудочно-кишечного тракта. Электрофизиологическими методами в мозжечке обнаружены висцеральные проекции внутренних органов. Так, например, при раздражении интероцепторов в коре мозжечка регистрируются
вызванные потенциалы.
Все эти экспериментальные факты позволили Л. А. Орбели считать мозжечок
млекопитающих не только регулятором моторной деятельности, но и адаптационно-
трофическим органом, который определяет возбудимость вегетативных и соматических центров.
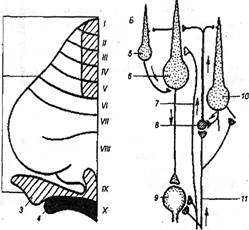
Рис. 4. Морфофункционалъная организация мозжечка
1-Х — доли мозжечка по номенклатуре Ларсела;
1— ростральная доля,
2 — каудальная
3 — околоклочок,
4 — клочково-узелковая доля,
5 — корзинчатая клетка,
6 — клетка Пуркинье,
7 — лазающее волокно,
8— клетка-зерно,
9 — клетка внутримозжечкового ядра,
10 — клетка Гольджи,
11 — мшистое волокно
БАЗАЛЬНЫЕ ГАНГЛИИ
Все отделы, лежащие выше промежуточного мозга, образуют передний мозг. Он состоит из двух частей: наружного слоя, или коры и глубинных образований - так называемых базальных ганглиев.
К базальным ганглиям относят: полосатое тело (стриатум), состоящее из хвостатого ядра и скорлупы; бледный шар (паллидум), подразделяющийся на внутренний и внешний отделы; черная субстанция; субталамическое ядро. В состав базальных ганглиев часто включают также ограду и реже - миндалину.
Базальные ганглии головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности. Причем, каждая из функций может быть усилена или заторможена активацией отдельных образований базальных ядер.
Базальные ганглии, как и мозжечок, используются в качестве системы, в которой уточняется программа выполнения сложных движений. Информация от ассоциативных участков коры, т. е. мест, где зарождается замысел движения, поступает одновременно к мозжечку и параллельно к базальным ганглиям, а от них через таламус поступает в двигательную кору, где уточненная программа используется для управления: информация идет через пирамидный и экстрапирамидный путь к а-мотонейронам спинного мозга. При нарушении этой сложной связи происходят изменения в двигательной сфере человека.
Связи между отдельными структурами базальных ганглиев осуществляются
следующим образом: информация от ассоциативной коры попадает к полосатому телу (хвостатое ядро + скорлупа), а от него информация идет по двум каналам:
1) к черной субстанции, от которой информация возвращается к полосатому телу (дофаминергический путь) и одновременно от черной субстанции направляется к таламусу;
2) информация от полосатого тела поступает к бледному шару, от него - к таламусу. Вся информация, пришедшая к таламусу от бледного шара и от черной субстанции, в
дальнейшем поступает к двигательной коре.
Большая часть афферентных сигналов, приходящих к базальным ганглиям, поступает в полосатое тело. Эти сигналы исходят почти исключительно из трех источников:
- от всех областей коры больших полушарий
- от внутрипластинчатых ядер таламуса
- от черной субстанции (по дофаминергическому пути).
Эфферентные волокна от полосатого тела идут к бледному шару и черной субстанции. От черной субстанции начинается не только дофаминергический путь к полосатому телу, но также и пути, идущие к таламусу. От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных трактов базальных ганглиев, заканчивающийся преимущественно в таламусе и в меньшей степени в крыше среднего мозга. Таким образом, базальные ганглии играют главным образом роль промежуточного звена в цепи, связывающей двигательные области коры со всеми
остальными ее областями.
От черной субстанции идут нисходящие пути к у-мотонейронам спинного мозга и к клеткам. Реншоу, благодаря чему она контролирует активность а-мотонейронов спинного мозга.
Бледный шар имеет выход к структурам ствола мозга, в частности - к ретикулярной формации, а от нее - к ретикулоспинальному пути. А также бледный шар координирует ориентировочную реакцию, движения конечностей, пищевое поведение (жевание, глотание и т.д.).
Итак, путь "ассоциативная кора - полосатое тело - бледный шар (и параллельно – черная субстанция) - таламус - двигательная кора" играет важную роль в деятельности мозга.
При нарушении функции бледного шара и черной субстанции наблюдается гипокинезия, мышечная гипертония (ригидность), тремор покоя (исчезает при движении). Больным трудно начать движение и трудно завершить начатое движение. Симптомокомплекс этот известен как болезнь Паркинсона. Развитие его связывают с тем, что нарушается функция дофаминергических нейронов черной субстанции.
Дофамин синтезируется в клетках черной субстанции. Он подавляет фоновую активность большинства нейронов хвостатого ядра, а это позволяет снять тормозящее действие этого ядра на активность бледного шара. При недостатке дофамина в неостриатуме, что наблюдается при дисфункции черного вещества, нейроны бледного шара растормаживаются, активизируют спинно-стволовые системы, это приводит к двигательным нарушениям в виде ригидности мышц.
Дата добавления: 2019-02-12; просмотров: 385; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!