Глава 6. Молекулярная генетика
Поиски причин – занятие, не имеющее конца.
У каждой причины есть своя причина, и нам приходится изучать все более и более отдаленные моменты в прошлом.
Н. Тинберген (1907–1988), выдающийся голландский биолог, один из основателей этологии, лауреат Нобелевской премии 1973 г.
Молекулярная генетика изучает молекулярные основы наследственности и изменчивости. Основное положение молекулярной генетики связано с признанием ведущей роли нуклеиновых кислот как хранителей и переносчиков генетической информации (Стент Г., Кэлинджер Р., 1981).
6.1. Структурно‑функциональная организация гена
Проблема гена – центральная проблема молекулярной генетики. Она берет свое начало с работы Т. Моргана «Теория гена» (1926), в которой ген был представлен как неделимая единица мутации (ген изменяется как целое ), рекомбинации (кроссинговер происходит в пределах гена ) и функции (все мутации одного гена связаны с одной функцией ).
С тех пор представления о гене радикально изменились. Важным этапом в развитии теории гена были работы С. Бензера в конце 1950‑х гг. (Benzer S., 1961). Они показали, что ген, представляющий собой нуклеотидную последовательность, не является неделимой единицей рекомбинации и мутации. Генетические исследования бактерий и фагов, благодаря гигантской численности их потомства, позволили уловить ничтожные доли (≈0,0001 %) внутригенных рекомбинаций, что подтверждало принцип дробимости гена. Единицу рекомбинации С. Бензер назвал реконом , а единицу мутации – мутоном . В дальнейшем было показано, что мутон и рекон соответствуют одной паре нуклеотидов. Единица генетической функции, которую С. Бензер назвал цистроном, совпадала с понятием ген, поэтому этот термин практически исчез из употребления (иногда термин цистрон еще употребляется в генетике как синоним понятия гена, когда хотят подчеркнуть его функциональное значение). С. Бензеру принадлежит крылатое выражение: «Гены – это атомы наследственности».
|
|
|
Современная теория гена сформировалась в русле нового направления, которое Дж. Уотсон назвал молекулярная биология гена (Уотсон Дж., 1978). Исследования тонкой структуры гена были проведены у вирусов, бактерий, грибов, высших эукариот. Что же показали эти исследования?
Основополагающий принцип классической генетики «один ген – один белок» подвергся серьезному пересмотру. В упрощенном виде под геном подразумевалась последовательность нуклеотидов, кодирующая одну полипептидную цепь, расположенная между старт‑сигналом и стоп‑сигналом. Затем были идентифицированы гены, кодирующие различные виды РНК, что потребовало уточнения в определении. Но новые открытия ставили новые проблемы. Чем дальше развивалась молекулярная генетика, тем труднее было дать четкое определение понятию «ген».
|
|
|
Неожиданный результат принесло изучение вирусных геномов. В 1977 г. Ф. Сэнджер у бактериофага φХ174 обнаружил «перекрывающиеся» гены, имеющие общие нуклеотидные участки. Бактериофаг φХ174 имеет кольцевую одноцепочечную ДНК и поражает клетки E. coli (Sanger F. [et al.], 1977). Затем «перекрывающиеся» гены выявили в геномах других организмов, включая человека. Изредка встречаются варианты, когда внутри одного гена целиком содержится другой, меньший – «ген в гене».
Необходимо отметить, что в «перекрывающихся» генах каждый нуклеотид принадлежит одному кодону, т. е. имеются различные рамки считывания с одной и той же нуклеотидной последовательности. Так, у фага φХ174 имеется участок молекулы ДНК, который входит в состав сразу трех генов. Но соответствующие этим генам последовательности нуклеотидов прочитываются каждый в своей системе отсчета. Поэтому нельзя говорить о «перекрывании» кода.
Если у вирусов такая организация генетического материала позволяет осуществлять экономное использование небольших информационных возможностей своего генома, то значение «перекрывания» в огромных геномах эукариот до конца не понятно. Возможно, эта роль связана с регуляцией активности генов путем образования двух почти комплементарных РНК. Такие молекулы РНК способны образовывать двунитиевые структуры, что блокирует процесс трансляции. «Экономия места» имеет свои побочные эффекты, поскольку одна мутация может «выключить» сразу два или более генов.
|
|
|
Сенсационным открытием явилась показанная в том же 1977 г. будущими нобелевскими лауреатами Р. Робертсом и Ф. Шарпом прерывистая, «мозаичная», структура большинства эукариотических генов (Brown D., 1981). В структуре гена стали выделять экзоны – участки гена, кодирующие структуру полипептида, и интроны – участки гена, не кодирующие структуру полипептида. Термины «экзон» и «интрон» были предложены У. Гилбертом (Gilbert W., 1981). Количество интрон‑экзонных переходов в пределах гена широко варьирует. В геноме человека одни гены имеют 3–10 таких переходов, другие – более сотни. Так, ген коллагена имеет 118 экзонов. Колебание размеров более характерно для интронов (например, у человека – от 14 до 150 000 п. н.). Для некоторых эукариотических генов экзоны составляют лишь незначительную часть их длины. Только единичные гены человека лишены интронов, в том числе все гены гистонов и мт‑ДНК. Роль интронов до конца не ясна. Вероятно, они участвуют в процессах генетической рекомбинации, а также в процессах регуляции экспрессии.
|
|
|
Дальнейшие исследования в области молекулярной биологии еще больше осложнили четкость определения понятия «ген». В геноме эукариот были обнаружены обширные регуляторные области. Относить ли к гену окружающие его регуляторные области или оставить в понятии «ген» только участок транскрипции – здесь мнения генетиков разделились. Проблема осложняется тем, что регуляторные области могут лежать за пределами единиц транскрипции на расстоянии в десятки тысяч п. н. Более того, одни и те же регуляторные участки могут «обслуживать» разные гены.
В регуляторной части генома выделяют различные участки.
Промотор – небольшой участок (у человека – 75 п. н.) связывания с ДНК факторов транскрипции и образование комплекса ДНК – РНК‑полимеразы для запуска синтеза РНК.
Энхансеры – усилители транскрипции.
Сайленсеры – ослабители транскрипции.
Между энхансерами и сайленсерами нет четкого «разделения труда», поскольку обычно они взаимодействуют со многими генами. Одна и та же последовательность ДНК может выступать и в роли энхансера, и в роли сайленсера, в зависимости от типа клеток. Данные последовательности представляют собой короткие участки ДНК (100–300 п. н.), являющиеся местом прикрепления регуляторных белков. Каждый энхансер или сайленсер может взаимодействовать с целым рядом регуляторных белков. Это изменяет активность генов путем изменения конформации определенного участка ДНК. В роли энхансеров и сайленсеров выступают некоторые транспозоны, что позволяет понять их генетическую роль.
Инсуляторы – короткие последовательности (300–1000 п. н.), обеспечивающие относительную независимость функций гена, блокируя взаимодействие между энхансером и промотором.
В последних моделях структурно‑функциональной организации генома предполагается, что ДНК‑нуклеосомная нить образует функциональные специфические участки – домены, которые представляют собой петли (обычно размером 30 000–200 000 п. н.), прикрепленную к структурам ядерного матрикса. В этих моделях инсуляторам отводится важная роль, во многом определяющая функционирование домена, который, вероятно, представляет собой единую функциональную единицу (Корочкин Л. И., 2002).
Между генами существуют особые межгенные последовательности – спейсеры.
Упрощенную структуру эукариотического гена, включающую транскрибируемые и регуляторные области, можно представить следующим образом (рис. 6.1).
Нетранслируемые области выполняют регуляторную роль в процессе трансляции.
Большинство генов бактерий представлены непрерывными участками ДНК, вся информация которой используется при синтезе полипептида. Участки ДНК между генами у прокариот весьма незначительны, а внутри оперона их нет совсем.
В организации митохондриального генома эукариот много общего с геномом прокариот, что служит веским доводом в пользу симбиотической теории происхождения митохондрий. Генымт‑ДНК расположены компактно, в них практически отсутствуют интроны и спейсеры. В ряде случаев гены даже перекрываются. У человека 93 % мт‑ДНК являются кодирующими. Показана значительная гомология мт‑ДНК человека и мыши.

Рис. 6.1. Структура эукариотического гена:
1 – энхансеры; 2 – сайленсеры; 3 – промотор; 4 – экзоны; 5 – интроны; 6 – участки экзонов, кодирующие нетранслируемые области
Другой удивительной особенностью генома эукариот явились повторяющиеся последовательности, т. е. последовательности ДНК, присутствующие в количестве нескольких копий. По частоте в геноме эукариот можно выделить три типа последовательностей ДНК (Айала Ф., Кайгер Дж., 1988). Это уникальные последовательности , представленные одной или несколькими копиями; умеренные повторы , представленные от десятка до нескольких тысяч копий на геном; высокоповторяющаяся ДНК , представленная от нескольких тысяч до миллиона копий на геном. Большинство функционирующих генов являются уникальными последовательностями, некоторые представлены умеренными повторами. Уникальные последовательности преобладают и в межгенных участках, но именно многократно повторяющиеся последовательности этих областей и вызывают особый интерес, во многом оставаясь загадкой.
Структурно различают тандемные повторы, которые расположены вплотную друг к другу, образуя блоки (кластеры), и диспергированные повторы, которые разбросаны по геному.
Тандемные повторы образуют особую сателлитную ДНК. Число разных копий в сателлитной ДНК варьирует от сотен до миллионов. Размер единицы повторов редко превышает 200 нуклеотидов, но может состоять и из одной «буквы». Недавно были обнаружены «мегасателлиты» размером до 5000 п. н., которые повторяются 50–400 раз (Тарантул В. З., 2003). Локализована сателлитная ДНК преимущественно в гетерохроматиновых районах, особенно в области центромеры и теломеры. Только «микросателлиты», представляющие повторы единиц из 1–4 нуклеотидов, рассеяны по всему геному.
Диспергированные повторы более разнообразны и многочисленны. Их размер обычно колеблется от 100 до 10 000 п. н. Они присутствуют во всех хромосомах человека и других млекопитающих. Сложная классификация диспергированных повторов включает различные группы и подгруппы, однако границы между ними размыты. Рекордную частоту в геноме человека (более миллиона копий на геном) имеют Alu‑повторы, размером около 300 п. н. Большинство диспергированных повторов относится к группе транспозонов, уникальной характеристикой которых, как уже говорилось выше, является способность перемещаться по геному.
Характеристика конкретных повторов в определенных местах генома играет важную роль в генетической идентификации личности.
Эволюционное значение повторов мы рассмотрим ниже, а сейчас отметим, что среди повторяющихся участков генома неожиданным открытием явились так называемые псевдогены – нефункционирующие последовательности ДНК, сходные с функционирующими генами (Proudfoot N., 1980). В геноме человека, например, около 20 000 псевдогенов. В частности, в семействе генов‑рецепторов обоняния их почти 60 %. Псевдогены еще больше осложнили определение понятия «ген». Можем ли мы псевдогены считать генами? И что же все‑таки такое ген?
Таким образом, используя термин «ген» для обозначения определенной последовательности ДНК, мы теперь вкладываем в него возможность прерывистой структурной организации, возможность участия части этой последовательности в составе другого гена, неоднозначность экспрессии этого участка, наличие генов как для белков, так и для РНК.
Подводя итог исследований молекулярной биологии гена и понимая всю сложность этой проблемы, остановимся на лаконичном определении гена, которое приводит в своей книге В. З. Тарантул: «Ген – это физическая (определенный участок ДНК) и функциональная (кодирует белок или РНК) единица наследственности» (Тарантул В. З., 2003). Размеры гена варьируют в чрезвычайно широких пределах. Так, самый маленький ген человека (МСС‑7) имеет всего 21 п. н., а самый большой (ген дистрофина ) – 2,2 млн п. н. (Гринев В. В., 2006).
Генетический код
Генетическая информация записывается последовательностями нуклеотидов в нуклеиновых кислотах с помощью 4 символов, как информация текста книги записывается с помощью букв, количество которых зависит от конкретного алфавита. В структуру белка эта информация «переписывается» с помощью 20‑буквенного «алфавита» природных аминокислот. Для перезаписи нуклеотидной последовательности нуклеиновых кислот в последовательность аминокислот должна существовать система кодировки, которая и получила название генетического кода. Генетический код – это соответствие определенной последовательности нуклеотидов определенной аминокислоте.
Выяснение природы генетического кода и экспериментальное определение смысла каждого кодона можно отнести к самым выдающимся достижениям современной науки. Первые результаты были представлены Ф. Криком и соавторами на V Международном биохимическом конгрессе в Москве в 1961 г. (Crick F. [et al.], 1961). Генетический код был полностью расшифрован к 1966 г. В его расшифровке принимали участие ведущие ученые всего мира. Он имеет следующие основные характеристики:
Триплетность – каждая аминокислота зашифрована последовательностью из 3 нуклеотидов (триплетом или кодоном ).
Вырожденность – большинство аминокислот шифруются более чем одним кодоном (от 2 до 6).
Неперекрываемость – один и тот же нуклеотид не может входить одновременно в состав 2 соседних кодонов.
Универсальность – характерен для всех организмов живой природы.
Универсальность генетического кода свидетельствует о единстве происхождения всех живых организмов.
Первое исключение из правила универсальности продемонстрировали митохондрии, причем в митохондриях разных организмов значение некоторых кодонов варьировало. Тогда стали говорить о квазиуниверсальности генетического кода. В последнее время найдены единичные отклонения генетического кода других «экзотических» объектов. К ним относятся микоплазмы , некоторые грибы, инфузории , хлоропласты растений. Российский генетик В. А. Ратнер пишет: «Это своеобразные „диалекты“ генетического кода, отражающие специфику их эволюции» (Ратнер В. А., 2002). Обнаруженные вариации генетического кода показывают, что код эволюционировал. В определении направления эволюции генетического кода между генетиками нет единого мнения.
В универсальном генетическом коде 61 кодон кодируют 20 аминокислот. Три кодона не соответствуют никакой аминокислоте и определяют момент окончания синтеза полипептида. Это так называемые терминирующие кодоны (стоп‑кодоны) – УАА, УАГ, УГА. Они играют роль знаков препинания между генами. Соответствие структуры гена (в нуклеотидах) и структуры кодируемого им белка (в аминокислотах) получило название коллинеарности. Интересно отметить, что лишь прямое подтверждение коллинеарности в 1964 г. послужило окончанием более чем 10‑летних дискуссий вокруг гипотез реализации наследственной информации.
Значения кодонов генетического кода приведены в табл. 6.1. Указаны аминокислоты, встречающиеся в белках, и соответствующие им кодоны информационной РНК.
Таблица 6.1. Генетический код
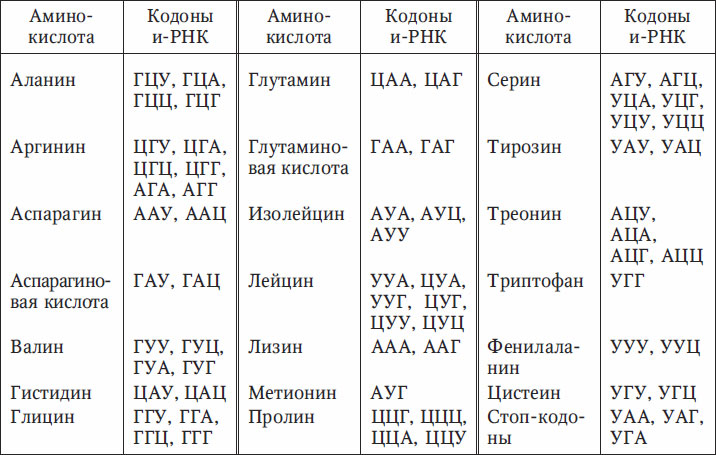
Еще в 1963 г. Г. Понтекорво, открывая симпозиум «От менделевских факторов к генетическому коду», подчеркнул, что все биохимические процессы клетки можно разделить на ступенчатые и матричные. Ступенчатые процессы – это обмен низкомолекулярных соединений, матричные – это синтез макромолекул (белков и нуклеиновых кислот). Активность генов непосредственно связана с матричными процессами. В процессе репликации происходит воспроизведение генетического материала. Реализация генетической информации – экспрессия генов, выражается в процессах транскрипции и трансляции.
Транскрипция
Транскрипцией называется процесс переноса генетической информации с ДНК на РНК. Матрицей для синтеза РНК служит только одна из двух цепей ДНК, так называемая смысловая цепь (3'→5' ). Однако в этом правиле встречаются исключения. Так, в мт‑ДНК обе цепи являются смысловыми с локализацией в них разных генов, причем процесс транскрипции идет на них в противоположных направлениях. Некоторые ядерные гены также расположены на «антисмысловой» цепи, с которой и происходит их транскрипция.
У прокариот гены одного оперона транскрибируются вместе. У эукариот транскрипция обычно происходит на участке только одного гена. Транскрипция мт‑ДНК эукариот осуществляется на единый мультигенный транскрипт, который затем разрезается. Во всех случаях транскрипция идет по единому принципу в 3 стадии.
1. Инициация – начало синтеза. Происходит присоединение к промотору комплекса ферментов, основным из которых является ДНК‑зависимая РНК‑полимераза . Это сложный белок, состоящий из нескольких субъединиц и играющий ключевую роль в процессе транскрипции. Промоторы прокариот и эукариот содержат короткие универсальные последовательности нуклеотидов, которые распознаются РНК‑полимеразами и служат местом их присоединения. У прокариот имеется один тип РНК‑полимеразы , у эукариот – три:
– РНК‑полимераза‑1 – для синтеза 3 видов (из 4) р‑РНК;
– РНК‑полимераза‑2 – для синтеза гя‑РНК (предшественника и‑РНК);
– РНК‑полимераза‑3 – для синтеза т‑РНК, мя‑РНК и 1 вида р‑РНК (5S‑РНК).
Присоединение РНК‑полимеразы к промотору инициирует раскручивание двойной цепочки ДНК и освобождение нуклеотидных связей.
2. Элонгация – последовательное присоединение свободных нуклеотидов к «смысловой цепи» ДНК по принципу комплементарности (А‑У, Г‑Ц) и соединение их при помощи РНК‑полимеразы в единую полирибонуклеотидную цепочку.
3. Терминация – завершение синтеза РНК в участке‑терминаторе, который узнается РНК‑полимеразой при участии особых белковых факторов терминации .
В результате процесса транскрипции синтезируются разные молекулы РНК. Эти молекулыдля своей функциональной активности чаще всего должны пройти этап тех или иных модификационных изменений. На этом этапе мы можем наблюдать принципиальное различие в экспрессии генов прокариот и эукариот. Если у прокариот процессы транскрипции и трансляции идут практически одновременно, то у эукариот эти этапы разделены во времени. Экспрессия генов у эукариот в ходе эволюции становится сложным и многоступенчатым процессом. Она включает в себя дополнительный этап формирования функционально активных молекул РНК, который получил название процессинга. Протекание процессинга в клетке имеет несколько вариаций.
При экспрессии генов, кодирующих структуру белка, в результате процесса транскрипции, который заканчивается в зоне терминации, образуется гетерогенная ядерная РНК (гя‑РНК). Она копирует всю нуклеотидную последовательность ДНК от промотора до терминатора, включая нетранслируемые области (рис. 6.2).

Рис. 6.2. Структура гя‑РНК:
1–5'‑нетранслируемая область; 2–3'‑нетранслируемая область; 3 – копии экзонов; 4 – копии интронов
После этого гя‑РНК претерпевает процессинг или процесс образования функционально активных м‑РНК. Он включает в себя процесс вырезания интронов и соединение экзонов – сплайсинг, процесс присоединения 7‑метил‑ГТФ к 5' – концу гя‑РНК с образованием «кэпа» («шапочки») – кэпирование и процесс присоединения полиаденилового участка (поли‑А) размером в 100–250 нуклеотидов к 3 – концу – полиаденилирование. В результате процессинга образуется матричная РНК (рис. 6.3).

Рис. 6.3. Структура м‑РНК:
1 – «кэп»; 2 – поли‑А‑участок; 3 – копии экзонов
Обычно гя‑РНК в несколько раз (иногда в десятки раз) больше м‑РНК.
Предполагается, что функция «кэпа» связана с инициацией процесса трансляции в результате прикрепления лидирующего участка м‑РНК к определенному участку рибосомы, а полиадениловый «хвост» защищает м‑РНК от ферментативного разрушения во время транспортировки к рибосомам. Точность сплайсинга регулируют мя‑РНК, которые имеют участки, комплементарные концам интронов.
У прокариот и‑РНК образуется в результате транскрипции сразу, и процессинга этот вид РНК не претерпевает.
Процессинг ядерных р‑РНК и т‑РНК проходит по другим схемам, без процессов кэпирования и полиаденилирования, хотя сплайсинг часто происходит. Так, 3 вида р‑РНК (из 4) у эукариот образуются в сложном процессе из одного транскрипта (45S‑РНК) путем его расщепления. Также происходит процессинг некоторых видов т‑РНК эукариот, хотя другие т‑РНК образуются без расщепления. Процессинг т‑РНК часто представляет собой длинную цепь модификаций нуклеотидов (метилирование, дезаминирование и др.)
Процессы модификации т‑РНК и р‑РНК происходят и у прокариот, что позволяет характеризовать их как прокариотический процессинг.
Молекула и‑РНК (после процессинга у эукариот и без процессинга у прокариот) участвует в другом матричном процессе – трансляции (синтезе белка).
Трансляция
Трансляцией называется процесс синтеза полипептидной цепочки на нити матричной РНК. Данный процесс протекает на рибосомах, поэтому вначале необходимо рассмотреть этот важнейший органоид клетки.
Рибосомы представляют собой миниатюрные клеточные «фабрики белка». Они являются клеточными органоидами, состоящими из белков (примерно 40 %) и р‑РНК (60 %). Это самый многочисленный органоид – их число в клетке может достигать 10 млн.
Все рибосомы состоят из двух субъединиц – малой и большой (рис. 6.4). Размер рибосом и их субъединиц выражается скоростью седиментации частиц в растворе (S – константа Сведберга ). Рибосомы прокариот характеризуются значениями 70S (30S + 50S), эукариот – 80S (40S + 60S). Рибосомы хлоропластов и митохондрий похожи на рибосомы прокариот, но отличаются значительной вариабельностью по размерам. Рибосомы содержат два участка – А (аминоацильный) и Р (пептидильный), являющимися основными каталитическими центрами. Помимо них, имеются и другие центры связывания ферментов. Специфичность участков определяется сочетанием соответствующих областей обеих субъединиц. При диссоциации субъединиц их специфичность теряется.
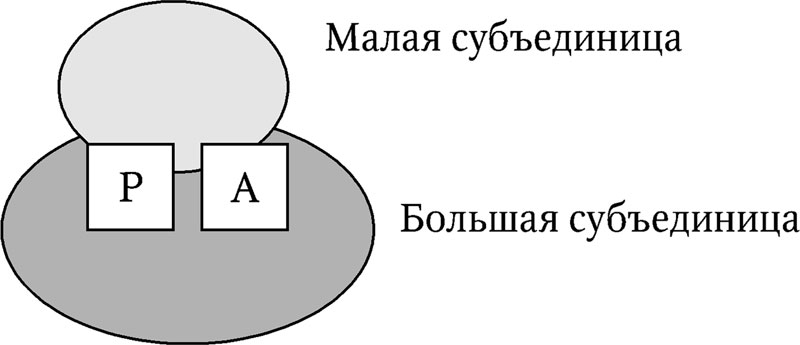
Рис. 6.4. Структура рибосомы
Как уже говорилось выше, рибосомы эукариот имеют в своем составе 4 разновидности р‑РНК. Три из них образуются из единого предшественника (45S‑РНК), синтез которого происходит в специализированной ядерной структуре – ядрышке – при помощи РНК‑полимеразы‑1. В ядрышках эукариот концентрируются петли хромосом, содержащие гены р‑РНК. Эти гены обычно имеют много копий. Так, у человека 200 копий генов р‑РНК располагаются на концах 5 пар хромосом (т. е. они имеются на 10 хромосомах из 46), поэтому сразу после митоза можно видеть 10 маленьких ядрышек, которые быстро сливаются в одно большое. Синтез 4‑й р‑РНК (5S‑РНК) происходит вне ядрышка (у человека на 1‑й хромосоме).
После поступления м‑РНК на рибосомы и начинается процесс трансляции, в котором выделяют 4 стадии:
1. Стадия активации аминокислот. Активация свободных аминокислот осуществляется при помощи особых ферментов (аминоацил‑т‑РНК‑синтетаз) в присутствии АТФ. Для каждой аминокислоты существуют свои ферменты и свои т‑РНК. Активированная аминокислота присоединяется к своей т‑РНК с образованием комплекса аминоацил‑т‑РНК (аа‑т‑РНК). Только активированные аминокислоты способны образовывать пептидные связи и формировать полипептидные цепочки.
2. Инициация. Начинается с присоединения лидирующего 5'‑конца и‑РНК с малой субъединицей диссоциированной рибосомы. Соединение происходит так, что стартовый кодон (обычно АУГ) оказывается в «недостроенном» Р‑участке. Комплекс аа‑т‑РНК с помощью антикодона т‑РНК присоединяется к стартовому кодону и‑РНК. Имеются многочисленные (особенно у эукариот) белки – факторы инициации.
У прокариот стартовый кодон кодирует N‑формилметионин, а у эукариот – N‑метионин. В дальнейшем эти аминокислоты вырезаются ферментами и не входят в состав белка. После образования инициирующего комплекса происходит объединение субъединиц и «достраивание» Р– и А‑участков рибосом.
3. Элонгация. Начинается с присоединения в А‑участке и‑РНК второго комплекса аа‑т‑РНК с антикодоном, комплементарным следующему кодону и‑РНК. На рибосоме оказываются две аминокислоты, между которыми возникает пептидная связь. После соединения аминокислот первая т‑РНК освобождается от аминокислоты и покидает рибосому. Рибосома перемещается вдоль нити и‑РНК на один триплет (в направлении 5' → 3' ). Вторая аа‑т‑РНК перемещается в Р‑участок, освобождая А‑участок, который занимает следующая аа‑т‑РНК. Таким же образом присоединяются 4‑я, 5‑я и т. д. аминокислоты, принесенные своими т‑РНК.
4. Терминация. Завершение синтеза полипептидной цепочки. Наступает тогда, когда рибосома дойдет до одного из терминирующих кодонов. Имеются особые белки (факторы терминации), которые узнают эти участки.
На одной молекуле и‑РНК может располагаться несколько рибосом (такое образование называется полисома ), что позволяет осуществлять синтез нескольких полипептидных цепей одновременно.
Процесс биосинтеза белка проходит с участием большего количества специфических биохимических взаимодействий. Он представляет собой фундаментальный процесс природы. Несмотря на чрезвычайную сложность (особенно в клетках эукариот), синтез одной молекулы белка длится всего 3–4 секунды.
По аналогии с геномом, в последнее время получает распространение термин «протеом» как совокупность функциональных белков клетки. Необходимо заметить, что если гены во всех клетках организма одинаковы, то наборы белков весьма разнообразны, меняясь в разных типах клеток по ходу онтогенеза.
Дата добавления: 2019-02-12; просмотров: 371; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!