Генетика пола и сцепленное с полом наследование
Генетический механизм определения пола в природе обусловлен генами, локализованными на особых половых хромосомах, имеющихся в кариотипе. Пол, у которого в кариотипе одинаковые половые хромосомы, называется гомогаметным, а пол, у которого в кариотипе разные половые хромосомы, – гетерогаметным. Неполовые хромосомы кариотипа называются аутосомами.
Морфологически различающиеся половые хромосомы представляют собой пару гомологов, поскольку имеют гомологичный участок, что позволяет им конъюгировать в мейозе. Однако гомологичный участок половых хромосом гетерогаметного пола обычно очень мал, поэтому большинство их аллелей присутствуют в генотипе в единственном числе. Наличие только одного аллеля в генотипе диплоидного организма называется гемизиготой.
В природе встречаются разные варианты хромосомного определения пола. Чаще гетерогаметным полом является мужской, а гомогаметным – женский, что наблюдается у млекопитающих (рис. 4.3).
У птиц (рис. 4.4) гетерогаметным полом является женский (WZ), а гомогаметным – мужской (ZZ). У некоторых насекомых самцы и самки могут различаться числом половых хромосом (либо две одинаковые, либо одна). Наконец, у пчел самки диплоидны, а самцы – гаплоидны (возникают путем партеногенеза из неоплодотворенных яйцеклеток).

Рис. 4.3. У млекопитающих гетерогаметным полом является мужской, а гомогаметным – женский
|
|
|
Кариотип человека включает 44 аутосомы и 2 половые хромосомы – у женщин XX, у мужчин ХY. Однако половой кариотип не исчерпывает вопрос детерминации пола. Этот вопрос далее рассмотрен отдельно.
Половые хромосомы всегда несут различные гены, не связанные с формированием пола (например, цвет глаз у дрозофилы). Наследование генов, локализованных на половых хромосомах, получило называние сцепленного с полом наследования. Такие гены обычно обозначают в виде верхнего индекса соответствующей половой хромосомы(ХА, ХВ, Yс+ и т. д.).

Рис. 4.4. У птиц гетерогаметным полом является женский, а гомогаметным – мужской
У млекопитающих Х‑хромосома имеет довольно много генов, а Y‑хромосома, наоборот, мало. Так, у человека, по различным данным, Х‑хромосома несет более 700 генов, а Y‑хромосома – около 80. У самцов рецессивные гемизиготные гены Х‑хромосомы могут проявлять свой фенотипический эффект. У самок также одна из двух Х‑хромосом подвергается гетерохроматизации в раннем эмбриогенезе и инактивируется. Биологический смысл этого явления получил объяснение в гипотезе М. Лайон через механизм «дозовой компенсации», приводящий в соответствие дозы генов Х‑хромосом у разных полов. Процесс гетерохроматизации Х‑хромосом носит случайный характер, поэтому в разных клетках женского организма инактивированы разные Х‑хромосомы(либо отцовская, либо материнская), а значит, могут функционировать разные аллели гомологичных генов.
|
|
|
В генетике пола выделяют также такое понятие, как наследование, ограниченное полом. Оно обусловлено генами, локализованными на аутосомах, но фенотипически проявляющимися у разных полов по‑разному.
Формирование половых признаков, полового поведения – это сложный, многоступенчатый процесс, происходящий во время онтогенеза. Подробно он рассматривается в курсе биологии развития.

Глава 5. Изменчивость
Храбреца не разыгрывай перед судьбой,
Каждый миг она может покончить с тобой.
Твой доверчивый рот, услаждая халвою,
Что ей стоит подсыпать отравы любой?
Омар Хайям (1048–1123), персидский философ и поэт
Всем живым организмам свойственна изменчивость, под которой понимают свойство приобретать новые признаки. Изменения в генетическом материале организмов служат основой разнообразия жизни на Земле.
В отечественной традиции принято рассматривать генотипическую (наследственную) и модификационную (ненаследственную) изменчивость в едином разделе генетики. Среди генотипической изменчивости выделяют мутационную и особую комбинативную изменчивость – процесс формирования новых комбинаций генов.
|
|
|
В западной литературе эти явления чаще рассматриваются в самостоятельных разделах: «мутации», «рекомбинации», «модификации». Понятие «изменчивость» употребляется обычно при анализе эволюционной теории.
Мутации
Теория мутаций составляет одну из основ генетики. Ее основные положения были разработаны голландским ученым Г. де Фризом еще в начале XX в.
Мутации – это наследственные изменения генетического материала. Они характеризуются как редкие, случайные, ненаправленные события. Большая часть мутаций приводит к различным нарушениям нормального развития, некоторые из них летальны. Однако вместе с тем именно мутации являются исходным материалом для естественного отбора и биологической эволюции.
Частота мутаций возрастает под действием определенных факторов – мутагенов, способных изменять материал наследственности. В зависимости от их природы мутагены делятся на физические (ионизирующее излучение, УФ‑излучение и др .), химические (большое число различных соединений ), биологические (вирусы, мобильные генетические элементы, некоторые ферменты ). Весьма условно деление мутагенов на эндогенные и экзогенные. Так, ионизирующее излучение, помимо первичного повреждения ДНК, образует в клетке нестабильные ионы (свободные радикалы ), способные вторично вызывать повреждения генетического материала. Многие физические и химические мутагены являются также канцерогенами, т. е. индуцируют злокачественный рост клеток.
|
|
|
Частота мутаций подчиняется распределению Пуассона, применяемому в биометрии, когда вероятность отдельного события очень мала, а выборка, в которой может возникнуть событие, велика. Вероятность мутаций в отдельном гене довольно низкая, однако число генов в организме велико, а в генофонде популяции – огромно.
В литературе можно встретить различные классификации мутаций: по проявлению в гетерозиготе (доминантные, рецессивные ), по инициирующему фактору (спонтанные, индуцированные ), по локализации (генеративные, соматические ), по фенотипическому проявлению (биохимические, морфологические, поведенческие, летальные и др. ). На мой взгляд, эти показатели представляют собой скорее рабочие характеристики конкретных мутаций, а не основу для классификации.
Классифицируются мутации по характеру изменений генома. По этому показателю выделяют 4 группы мутаций, каждая из которых имеет многочисленные разновидности.
Генные мутации представляют собой изменения нуклеотидного состава ДНК отдельных генов. Мутационные изменения генов могут происходить в одной точке (односайтовые мутации ) либо в нескольких разных точках (многосайтовые мутации ). Термин «сайт » в генетике подразумевает определенное место в цепи молекулы ДНК. Современные методы молекулярной генетики позволили определить два основных процесса формирования генных мутаций: замена нуклеотидов и сдвиг рамки считывания, каждый из которых имеет свои варианты (рис. 5.1).
Транзиции – при замене сохраняется месторасположение пуриновых и пиримидиновых нуклеотидов (АТ → ГЦ, ГЦ → АТ и т. п.).
Трансверсии – при замене пуриновые и пиримидиновые нуклеотиды меняются местами (АТ → ЦГ, АТ → ТА и т. п.).
Мутации вследствие сдвига рамки считывания встречаются более часто. Они проявляются в двух вариантах: инсерция (вставка) и делеция (утеря) одного или нескольких нуклеотидов. Необходимо отметить, что вставка сдвигает рамку считывания в одном направлении, а делеция – в противоположном.
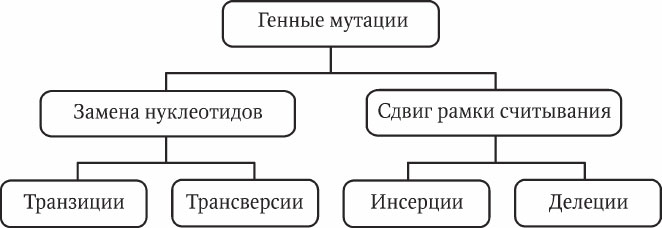
Рис. 5.1. Генные мутации
Механизм возникновения генных мутаций наиболее детально изучен у вирусов и бактерий. Согласно концепции американского генетика Р. фон Борстела, генные мутации возникают в результате ошибок «трех Р»: репликации, репарации и рекомбинации.
В процессе репликации возможна замена нуклеотидов вследствие некоторой неоднозначности принципа комплементарности. Азотистые основания нуклеотидов ДНК могут существовать в нескольких таутомерных формах. Таутомеризация – это изменение положения водорода в молекуле, меняющее ее химические свойства. Некоторые таутомеры нуклеотидов меняют способность формировать водородные связи с другими нуклеотидами. У аналогов нуклеотидов таутомерия происходит значительно чаще, чем у типичных форм, что объясняет их мутагенный эффект. Примером может служить аналог тимидина 5‑бромурацил, который способен в некоторых таутомерных формах вступать в связь с гуанином. Вследствие вырожденности генетического кода замены нуклеотидов могут иметь различные фенотипические проявления. Они могут не вызвать замены аминокислот или привести к появлению аминокислоты, близкой по своим свойствам, но могут привести к замене на аминокислоту с другими физико‑химическими свойствами или к образованию стоп‑кодона.
Большинство мутаций со сдвигом рамки считывания обнаружены в участках ДНК, состоящих из одинаковых нуклеотидов. Существует гипотеза возникновения этих мутаций вследствие диссоциации и неправильного восстановления нитей в данных участках.
Резкий рост мутаций при нарушении системы репарации и взаимосвязь мутационного и рекомбинационного процессов продемонстрированы в многочисленных исследованиях. Процессы репарации и рекомбинации мы рассмотрим ниже.
Хромосомные мутации (аберрации ) – это изменения структуры хромосом: внутрихромосомные или межхромосомные перестройки, возникающие при разрывах хромосом. Хромосомные перестройки обычно приводят к различным фенотипическим проявлениям. Выделяют следующие виды аберраций (рис. 5.2).
Дупликация – дублирование участка хромосомы.
Амплификация – многократное повторение участка хромосомы.
Повторы генетического материала не оказывают такого отрицательного влияния на организм как делеции и дефишенси. Показана значительная роль дупликаций в эволюции генома, поскольку они создают дополнительные участки генетического материала, доступные для мутирования, изменения функций генов и естественного отбора.
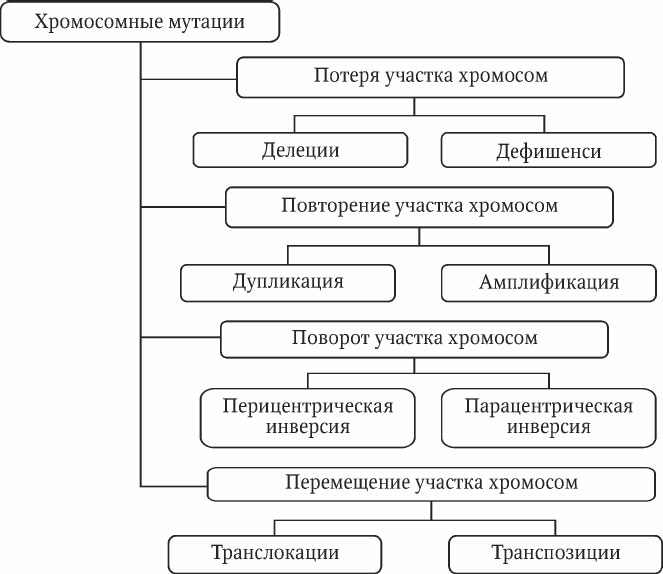
Рис. 5.2. Хромосомные мутации
Явление амплификации можно наблюдать при культивировании клеток с различными повреждающими агентами, но оно встречается и в природе как закономерный процесс онтогенеза, когда необходимо резко увеличить экспрессию каких‑либо генов. В последнем случае возможны два варианта: либо амплифицированная ДНК остается связанной с хромосомой, образуя многочисленные репликативные вилки (например, в фолликулярных клетках дрозофилы); либо синтезированная ДНК отделяется от материнской и многократно реплицируется (как ДНК, содержащая геныр‑РНК ооцитов амфибий).
Инверсия – поворот участка хромосомына 180°. Инверсия приводит к изменению линейной последовательности генов. Она встречается в двух вариантах: перицентрическая инверсия (центромера входит в инвертированный участок) и парацентрическая инверсия (центромера не входит в инвертируемый участок). Негативный эффект инверсии зависит от локализации точек разрывов, их близости к жизненно важным генам. Необходимо отметить, что инверсии встречаются в природных популяциях чаще других хромосомных перестроек. Они представляют собой распространенный путь преобразований генетического материала в эволюции, являясь факторами изоляции и дивергенции новых форм в пределах вида. Реципрокные транслокации – обмен участками хромосом между негомологичными хромосомами. В результате такой транслокации изменяется характер сцепления генов – гены, принадлежащие к разным хромосомам, могут наследоваться как одна группа сцепления. Характер конъюгации при транслокации меняется – вместо бивалентов образуется квадривалент в виде «фигуры креста» (рис. 5.3).
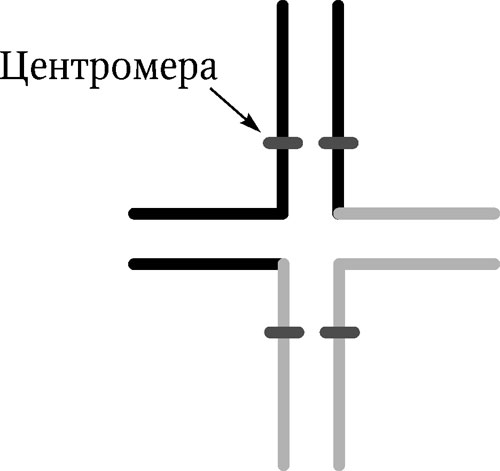
Рис. 5.3. Синапсис хромосом при реципрокной транслокации в профазе мейоза. На каждой хромосоме отмечена центромера
Гетерозиготы по реципрокным транслокациям обладают пониженной плодовитостью, так как продуцируют дефектные гаметы. Только у двух видов гамет из шести возможных при разных способах расхождения хромосом имеются полные комплекты генов. Остальные несут дупликации и нехватки и не могут дать жизнеспособного потомства. У животных реципрокные транслокации встречаются редко, обычно с негативным эффектом, но они широко распространены у растений. Благодаря специальным механизмам, транслокация обеспечивает изоляцию новых форм.
Участок хромосомы может менять свое положение и без реципрокного (взаимного) обмена, оставаясь на той же хромосоме или включаясь в другую. Такое явление называется транспозицией. Транспозиции будут рассматриваться ниже как важный самостоятельный раздел современной генетики.
Вероятно, все типы хромосомных перестроек имеют единый механизм и обусловлены лабильностью генома.
Причиной изменения фенотипа при различных хромосомных перестройках часто является изменение расположения гена. Этот феномен получил название эффект положения гена. Он показан для многих генов и обычно влияет на их регуляторную систему. Например, при перемещении гена из эухроматина в гетерохроматиновую область его активность утрачивается, хотя сам ген не изменяется.
Геномные мутации – это изменения числа хромосом. Среди геномных мутаций также выделяют несколько разновидностей (рис. 5.4).
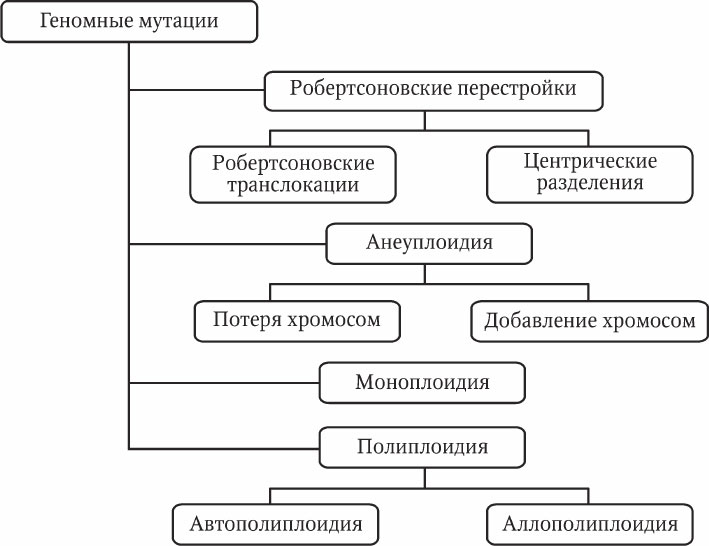
Рис. 5.4. Геномные мутации
Робертсоновские перестройки – слияния и разделения хромосом в области центромеры. Названы они по имени В. Робертсона, который предложил свою гипотезу механизма таких мутаций. Центрические слияния (робертсоновские транслокации ) представляют собой слияния двух негомологичных акроцентрических хромосом с образованием одной субметацентрической хромосомы. При разделении, наоборот, одна субметацентрическая хромосома делится на две акроцентрические хромосомы. При этом должна образоваться новая центромера, иначе хромосома без центромеры будет потеряна при митозе.
Робертсоновские перестройки приводят к изменению числа хромосом в кариотипе, не влияя на общее количество генетического материала в клетке. Оба варианта перестроек представлены в природе, но робертсоновские транслокации встречаются значительно чаще. Они являются одним из магистральных путей эволюции кариотипов.
Анеуплоидия – изменение числа хромосом, не кратное гаплоидному набору. Как правило, представляет собой добавление или потерю одной или двух хромосом диплоидного набора. У животных анеуплоидия обычно приводит к тяжелым аномалиям или летальности. Однако у растений трисомия (наличие трех гомологичных хромосом) может служить фактором генетического разнообразия. Причиной анеуплоидии является нерасхождение хромосом в мейозе и образование несбалансированных гамет.
Моноплоидия (гаплоидия) – гаплоидное состояние диплоидного организма. Эта мутация интенсивно изучается у растений, так как позволяет видеть проявление рецессивных аллелей. У животных моноплоидия обычно приводит к летальному исходу.
Автополиплоидия – наличие в клетке более двух одинаковых гаплоидных наборов. Эта разновидность мутации довольно широко представлена в природе у протистов, грибов и растений. Плоидность макронуклеуса инфузорий может достигать нескольких сотен. У животных встречается редко и обычно приводит к летальному исходу на ранних стадиях эмбриогенеза. У культурных растений сбалансированные полиплоиды (т. е. кариотипы с четным числом гаплоидных наборов – 4n, 6n, 8n и т. п.) получают искусственным путем из‑за их более крупных размеров. Несбалансированные полиплоиды (3n, 5n, 7n и т. п.) растений часто имеют пониженную фертильность вследствие нарушений мейоза. Тем не менее некоторые растения‑триплоиды (3n) обладают большими размерами и продуктивностью по сравнению с диплоидными (2n) и тетраплоидными (4n).
Аллополиплоидия – объединение в клетке разных геномов посредством гибридизации. В природе для многих цветковых растений описаны полиплоидные ряды различной степени плоидности. Эти ряды возникают путем гибридизации разных видов и последующего удвоения родительских гаплоидных наборов. Так преодолевается барьер бесплодия при скрещивании разных видов.
Цитоплазматические мутации – это изменения неядерных генов. Генетическая информация органелл (в отличие от хромосомной) представлена в зиготе в разном объеме от каждого из родителей. Чаще всего преобладает вклад материнского организма, но у некоторых видов наследуются внеядерные геномы от обоих родителей. Мутации генома органелл обычно представлены точечными мутациями и делециями. Основными носителями внеядерной генетической информации в клетках высших эукариот являются хлоропласты и митохондрии.
В хлоропластах фенотипическое проявление мутаций выражается нарушением фотосинтеза, изменением чувствительности к температуре и устойчивости к антибиотикам. Генетика хлоропластов разработана хуже, чем генетика митохондрий.
У позвоночных митохондрии служат единственным хранилищем внеядерного генетического материала. Источником митохондрий в зиготе, вероятно, служит только яйцеклетка. Хотя 95 % митохондриальных белков кодируется в ядре, мутации мт‑ДНК происходят в 10 раз чаще, чем в ядерных генах, поэтому их вклад ощутим.
Фенотипическое проявление мутаций мт‑ДНК во многом зависит от уровня потребления АТФ той или иной тканью. Поскольку наиболее энергозависимыми являются нервная и мышечная системы, такие мутации наиболее часто проявляются в форме различных нейропатий и миопатий.
Рекомбинация
Рекомбинация генетического материала в природе происходит на разных уровнях.
Рекомбинация несцепленных генов происходит благодаря механизму независимого распределения хромосом в мейозе и формирования новых генотипов при оплодотворении. Даже единичные мутантные аллели в популяции резко увеличивают генотипическое разнообразие. Именно этот фактор и является главным эволюционным преимуществом полового размножения.
Замечено, что чем выше уровень организации организмов, тем большее значение приобретает рекомбинация. Если у прокариот жизненно важные функции могут определяться единичными генами, то у высших животных фенотипические признаки обычно определяются сложными сочетаниями генов. Некоторые новые сочетания генов могут превысить предковый генотип по адаптационной ценности. Как сказал известный эволюционист В. Грант, «рекомбинация – это механизм, который производит сборку генных сочетаний » (Грант В., 1980).

Рис. 5.5. Генетическая карта хромосомы. Частота кроссинговера между генами А и В меньше, чем между генами А и С
Кроссинговер представляет собой реципрокную рекомбинацию сцепленных генов, лежащих в гомологичных локусах гомологичных хромосом. Происходит кроссинговер после процесса репликации на стадии четырех нитей. Механизм кроссинговера весьма сложен и рассматривается в специальных курсах. Современные представления о механизмах кроссинговера явились итогом долгих исследований, хотя и сейчас еще остается ряд нерешенных вопросов.
Как уже говорилось выше, частота кроссинговера является мерой расстояния между генами и служит основой для построения генетических карт. Если два сцепленных гена занимают локусы, расположенные рядом, кроссинговер между ними наблюдается редко. Наоборот, если локусы находятся далеко друг от друга, то частота кроссинговера между ними велика (рис. 5.5). При достаточном удалении сцепленные гены рекомбинируют с той же частотой, что и несцепленные.
На частоту кроссинговера оказывают влияние различные генетические факторы. Известны мутации, повышающие и понижающие частоту кроссинговера. Обычно эта частота ниже у гетерогаметного пола.
Сайт‑специфическая рекомбинация – это рекомбинация между молекулами ДНК, имеющими небольшие участки гомологии.
Впервые этот механизм был изучен при взаимодействии фага ë и кольцевой ДНК E. coli. Интеграция и эксцизия (вырезание) фага всегда происходят в области общей гомологии ДНК фага и бактерии, размером всего в 15 п. н. Этот процесс контролируют два фаговых гена.
Таким же путем происходит перемещение мобильных генетических элементов, которые мы рассмотрим в следующем разделе.
Сайт‑специфическая рекомбинация играет важную роль в процессе образования антител. Формирование многообразия антител является ключевым вопросом иммуногенетики – бурно развивающимся разделом современной биологии, находящимся на стыке иммунологии и генетики.
Не углубляясь в механизм сложных процессов интеграции и эксцизии, отметим, что эти процессы у вирусов не связаны с дополнительной репликацией ДНК. Наоборот, при транспозиции почти всегда наблюдается дополнительная репликация.
Транспозиции
Традиционные представления о стабильности геномов, сложившиеся в рамках классической генетики, были существенно поколеблены после открытия мобильных (мигрирующих) генетических элементов (МГЭ). МГЭ – это структуры, которые могут перемещаться в пределах генома и переходить из генома в геном. Они могут встраиваться в различные области хромосом эукариот, ДНК и плазмид прокариот. Процесс перемещения МГЭ получил название транспозиции.
МГЭ были открыты Б. Мак‑Клинток у кукурузы еще в 1940‑е гг. (рис. 5.6), однако ее сообщение было встречено с большим недоверием генетиков. Развитие молекулярной генетики подтвердило наличие подобных структур сначала у прокариот, а затем и у эукариотических организмов. Только в 1983 г. за свое выдающееся открытие Б. Мак‑Клинток была удостоена Нобелевской премии.

Рис. 5.6. Впервые мобильные генетические элементы были обнаружены у кукурузы
У прокариот выделяют два вида МГЭ.
Инсерционные последовательности (IS) – небольшие последовательности ДНК (700–2000 п. н.), имеющие на концах короткие (10–40 п. н.) схожие последовательности, расположенные в обратном порядке (инвертированные повторы).
IS несут гены, связанные только с транспозицией. При транспозиции участок ДНК хозяина в сайте‑мишени (5–9 п. н.) удваивается и окаймляет IS прямыми повторами, т. е. одинаково ориентированными (рис. 5.7).

Рис. 5.7. Схема внедрения IS в ДНК хозяина:
1 – удвоенный сайт‑мишень ДНК хозяина; 2 – инвертированные повторы; 3 – центральная область IS; 4 – ДНК клетки‑хозяина
Бактериальная клетка обычно имеет множество IS. Наличие идентичных IS в различных бактериальных генетических структурах создает основу для их взаимодействия, интеграции и генетического обмена по принципу сайт‑специфической рекомбинации.
Транспозоны (Tn ) – более сложные структуры (2000–20 000 п. н.), в средней части имеющие различные гены, не обязательно связанные с транспозицией.
Tn с каждой стороны имеют плечи (или модули), которые являются IS, в прямой или инвертированной ориентации (рис. 5.8). Поскольку IS всегда имеют инвертированные повторы, то и Tn имеют их на своих краях. Большие транспозоны группы TnА плеч не имеют, но инвертированными повторами (38 п. н.) все равно окаймлены.
Вероятно, Tn возникли путем объединения двух первоначально независимых IS и области между ними (Shapiro J., 1983). Многие транспозоны несут гены устойчивости к антибиотикам. Процесс транспозиции у бактерий включает дупликацию транспозона, при этом одна копия остается на старом месте, а другая возникает на новом.
Термин «транспозоны» в настоящее время обычно применяют ко всем МГЭ.

Рис. 5.8. Прямая (а) и инвертированная (б) ориентация плеч транспозонов
Структуры, аналогичные транспозонам бактерий, обнаружены у дрожжей, кукурузы, дрозофилы и других эукариотических организмов. Для них характерны типичные признаки транспозонов – инвертированные повторы и дупликация сайт‑мишени. У дрожжей структуры Ty (Transposon yeast ) встречаются в среднем в количестве 30–35 на клетку. Они имеют типичную структуру бактериального транспозона с плечами по 330 п. н. Такую же структуру имеют и самые первые из открытых МГЭ у кукурузы (около 5000 п. н.), получившие при открытии название «контролирующие элементы». У дрозофилы были обнаружены многочисленные транспозоноподобные структуры – МДГ (мобильные диспергированные гены ). Одной из возможных функций МДГ является создание изолирующих барьеров для скрещивания на пути к видообразованию.
Развитие методов молекулярной генетики показало широкое распространение МГЭ в геномах высших животных, в том числе и у человека. Их доля в геномах оказалась значительно выше, чем предполагалось ранее. В своем большинстве они являются эволюционным наследием и обусловлены внедрением вирусного или бактериального генома в клетки эукариот миллионы лет назад. В первую очередь, это относится к ретровирусам , способным строить на своей РНК комплементарную ДНК и таким образом внедряться в чужой геном. Встраиваемая в геном ДНК‑копия называется провирусом. Подавляющее большинство провирусов не функционируют (не экспрессируются), но при определенных воздействиях некоторые из них способны активироваться, что может представлять опасность для макроорганизма.
Хотя наличие МГЭ и у прокариот, и у эукариот указывает на их общебиологическое значение, окончательно не решен вопрос об их функциях. Феномен транспозиции представляет большой теоретический интерес, независимо от конкретной функции МГЭ. Их важная роль в эволюции уже не подлежит сомнению. Так, показана близость сайтов внедрения МГЭ и хромосомных перестроек. Поскольку МГЭ могут создавать диффузные области гомологии, их копии в разных местах генома обеспечивают возможность реципрокной сайт‑специфической рекомбинации. Такие обмены могут приводить к различным хромосомным перестройкам (делециям, инверсиям, транслокациям), изменять систему регуляции генов. Стимулируя хромосомные перестройки, транспозоны могут изменять экспрессию соседних генов. Такая реорганизация играет существенную роль в эволюции геномов.
Показан резкий рост числа транспозиций в популяциях дрозофилы при стрессовых воздействиях, что можно рассматривать как модель быстрой регуляции активности генома при изменении условий существования (Ратнер В. А., 2002).
В настоящее время наметилась тенденция оставить понятие «мутация» только за генными мутациями, а хромосомные и геномные мутации рассматривать как варианты генетической рекомбинации. Терминологическая проблема возникла из‑за традиционного понимания мутации как нерегулярного события, а главное, как отклонения от нормы. Однако дупликации, инверсии, транслокации, центрические слияния, полиплоидия неразрывно связаны с эволюцией геномов. С эволюционной точки зрения хромосома – это непостоянная структура, а лабильность – естественное состояние генома (Хесин Р. Б., 1984).
Все вирусы представляют собой «мобильные гены». Вирусы, плазмиды, транспозоны можно рассматривать в русле единого явления в эволюции генетической системы. Все эти структуры не только сами способны перемещаться по геному, но и могут «перетаскивать» соседние участки генома клетки‑хозяина. Такое явление получило название трансдукции. Многообразие мобильных генетических структур послужило основой для гипотезы горизонтального переноса генов в эволюции. Одни генетики, допуская возможность горизонтального переноса в природе, не считают, что он играл заметную роль в эволюции. Другие, наоборот, придают ему решающее значение. Проблема горизонтального переноса генов будет рассмотрена далее.
Репарация
Выявляемая частота мутаций не отражает истинную величину потенциальных повреждений ДНК. Повреждения ДНК сводятся к минимуму благодаря наличию в клетке особых систем репарации, которые узнают эти повреждения и исправляют их. Системы репарации возникли в процессе эволюции для поддержания стабильности генетической организации организмов. Некоторые репаративные системы обладают специфичностью, другие не специфичны в отношении каких‑то определенных типов повреждений – они узнают изменения в структуре ДНК как сигналы к действию. Репаративные системы представляют собой ферментативные механизмы, обнаруженные в клетках различных организмов.
Среди нарушений структуры ДНК, создающих препятствия для процессов репликации и транскрипции, наиболее хорошо изучен вариант образования тиминовых димеров (Т‑Т) под действием УФ‑облучения. Тиминовый димер образуется в результате возникновения ковалентных связей между смежными основаниями. Именно такие нарушения конформации служат мишенью для большинства систем репарации.
Выделяют три основных механизма репарации ДНК.
Фотореактивация – восстановление молекул ДНК под действием фермента фотолиазы, индуцируемого видимым светом после появления димеров. С неповрежденной ДНК фотолиаза не связывается.
Эксцизионная репарация – многоэтапный процесс удаления поврежденных нуклеотидов и синтеза новой последовательности ДНК. «Узнает» место повреждения фермент эндонуклеаза. Последующие этапы вырезания, удаления (эксцизии), ресинтеза ДНК, сшивания цепи осуществляют специальные ферменты. Различные виды эксцизионной репарации широко распространены как у прокариот, так и у эукариот.
Пострепликационная репарация – использование материала одной молекулы ДНК для восстановления другой. Этот механизм наименее специфичен, так как здесь отсутствует этап узнавания повреждения. Он также является наиболее быстрым способом репарации. Правда, повреждения остаются в исходной родительской ДНК, а исправляются только дочерние молекулы.
У человека известно наследственное заболевание – пигментная ксеродерма , – проявляющееся в гиперчувствительности к солнечным лучам, особенно к ультрафиолету. Причиной заболевания является нарушение системы репарации клетки, неспособность ликвидировать тиминовые димеры. Нередко этот дефект приводит к развитию рака кожи.
В рамках трех названных механизмов у прокариот и эукариот встречаются разнообразные системы репарации. Эволюция этих систем обусловлена особым значением сохранности генетической информации.
Дата добавления: 2019-02-12; просмотров: 391; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!