Рационы кормления коров опытной группы
| Корма, кг | 1 период – 10-25-й-день лактации | 2 период – 25-55-й день лактации | 3 период – 55-85-й день лактации | 4 период – 85-115-й день лактации |
| Сено | 4,5 | 4,5 | 4,5 | 4,5 |
| Силос разнотравный, | 15 | 15 | 18 | 21 |
| Патока кормовая | 0 | 1,5 | 1,5 | 1,5 |
| Комбикорм | 8,4 | 9,0 | 9,0 | 8,4 |
Таблица 45
Рационы кормления коров контрольной группы
| Корма, кг | 1 период 10-25-й день лактации | 2 период 25-55-й день лактации | 3 период 55-85-й день лактации | 4 период 85-115-й день лактации |
| Сено | 4,5 | 4,5 | 4,5 | 4,5 |
| Сенаж разнотравный | 15 | 15 | 18 | 21 |
| Патока кормовая | 0 | 1,5 | 1,5 | 1,5 |
| Комбикорм | 8,4 | 9,9 | 10,5 | 8,4 |
Таблица 46
Состав комбикормов по периодам опыта (%)
| Корма | Периоды опыта | |||||||
| Опыт | контроль | |||||||
| 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |
| Кукуруза | 8,3 | 18 | ||||||
| Ячмень | 40 | 29 | 27 | 45,4 | 40 | 40,1 | 45,4 | 45,4 |
| Пшеница | 18 | 14,1 | 14 | 25,6 | 18 | 23,2 | 25,6 | 25,6 |
| Отруби пшеничные | 13,5 | 6,2 | 10 | 13,5 | ||||
| Соевый шрот | 15,3 | 12 | 15 | 10,7 | 15 | 15 | ||
| Кукурузный жмых | 17,8 | 13,4 | ||||||
| Кукурузный глютен | 0,83 | 0,5 | 0 | 0 | 0 | |||
| Кровяная мука | 2,1 | 8 | 0 | 0 | 0 | |||
| Рыбная мука | 2,0 | |||||||
| Подсолнечный шрот | 25 | 0 | 10 | 25 | 8,9 | 10 | 10 | |
| Жир | 2,1 | 3 | 0 | |||||
| Мочевина | 0,83 | 1,5 | ||||||
| Премикс П-60-3 | 1 | 0,83 | 1 | 1 | 1 | 0,9 | 1 | 1 |
| Соль поваренная | 1 | 1,25 | 1 | 1,5 | 1,5 | 1,3 | 1,5 | 1,5 |
| Трикальций фосфат | 1,5 | 1,25 | 1,5 | 1,5 | 1,5 | 1,3 | 1,5 | 1,5 |
|
|
|
На 55-й и 8-й5 и 115 дне лактации проведены балансовые опыты, респирационные исследования, отбор проб крови, рубцового и дуоденального содержимого.
Каждая группа коров содержалась на рационах с оптимальным содержанием доступных питательных веществ в рационе (нераспадаемый протеин, фракции клетчатки, распадаемый и нераспадаемый крахмал, липиды) для данной фазы лактации и данного уровня молочной продуктивности (табл. 47, 48). В плазме артериальной крови лактирующих коров определена концентрация гормонов: инсулина, тироксина, трийодтиронина с целью выявления взаимосвязей этих гормонов с потоками метаболитов из желудочно-кишечного тракта при разном соотношении у коров. Определяли артерио-венозную разница метаболитов по молочной железе и эффективность их поглощения (глюкоза, ЛЖК, кетоновые тела, НЭЖК, триацилглицеролы, ВЖК, свободные аминокислоты и мочевина (Методы биохимического анализа, Боровск, 1997).
Таблица 47
Состав рационов по фактической поедаемости (г)
|
|
|
| № рационов | СВ (кг) | Зола | Сырая клетчатка | Жир | Лигнин | Гемицеллюл. | Целлюлоза | СП | НДК |
| 1-к | 16,5 | 1253,9 | 3715,6 | 436,8 | 1222,1 | 3139,8 | 2973,2 | 2182,7 | 7335,2 |
| 1-о | 16,4 | 1248,6 | 3690,6 | 435,5 | 1214,5 | 3125,0 | 2956,8 | 2178,8 | 7296,4 |
| 2к | 18,9 | 1033,7 | 3883,3 | 614,3 | 1315,7 | 3105,5 | 3030,3 | 2527,5 | 7451,5 |
| 2о | 18,3 | 1027,1 | 3878,2 | 699,9 | 1347,1 | 3006,4 | 2972,5 | 2477,0 | 7326,0 |
| 3к | 19,8 | 1478,7 | 4272,9 | 479,7 | 1348,2 | 3486,0 | 3262,5 | 2698,7 | 8096,6 |
| 3о | 19,1 | 1321,4 | 4359,6 | 780,9 | 1298,4 | 3541,7 | 3250,5 | 2919,9 | 8090,6 |
| 4к | 19,4 | 1510,3 | 4539,6 | 485,1 | 1473,0 | 3620,7 | 3502,2 | 2555,9 | 8595,9 |
| 4о | 19,1 | 1472,8 | 4401,2 | 477,2 | 1432,1 | 3554,1 | 3425,8 | 2535,9 | 8411,9 |
Таблица 48
Содержание питательных веществ по периодам опыта от СВ рационов (%)
| № рационов | Зола | Сырая клетчатка | Жир | Лигнин | Гемицеллюл. | Целлюлоза | СП | НДК |
| 1-к | 7,6 | 22,5 | 2,6 | 7,4 | 19,0 | 18,0 | 13,2 | 44,5 |
| 1-о | 7,6 | 22,5 | 2,7 | 7,4 | 19,1 | 18,0 | 13,3 | 44,5 |
| 2-к | 5,5 | 20,5 | 3,2 | 6,9 | 16,4 | 16,0 | 13,4 | 39,3 |
| 2-о | 5,6 | 21,2 | 3,8 | 7,4 | 16,5 | 16,3 | 13,5 | 40,1 |
| 3-к | 7,5 | 21,6 | 2,4 | 6,8 | 17,6 | 16,5 | 13,6 | 40,9 |
| 3-о | 6,9 | 22,8 | 4,1 | 6,8 | 18,5 | 17,0 | 15,3 | 42,4 |
| 4-к | 7,8 | 23,4 | 2,5 | 7,6 | 18,7 | 18,1 | 13,2 | 44,3 |
| 4-о | 7,7 | 23,0 | 2,5 | 7,5 | 18,6 | 17,9 | 13,3 | 44,0 |
Базовый рацион подопытных животных состоял из сена 20%, силоса –30% и концентратов-50% по питательности (табл.49).
|
|
|
Таблица 49
Структура рационов (% от СВ)
| Корма | Периоды опыта | |||||||
| Опыт | Контроль | |||||||
| 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |
| грубые корма | 55,1 | 49,8 | 53,5 | 57,5 | 55,1 | 47,6 | 49,9 | 57,5 |
| концентраты | 44,9 | 45,5 | 42,1 | 38,2 | 44,9 | 47,8 | 45,9 | 38,2 |
| патока | 0 | 4,67 | 4,36 | 4,2 | 0 | 4,46 | 4,07 | 4,2 |
Рационы коров контрольной и опытной группы до 25-го дня лактации были идентичными, в состав комбикорма включали компоненты с высокой доступностью для переваривания в рубце. В результате на 25-й день лактации потребление сухого вещества составило 3,3кг на 100кг живой массы.
В дальнейшем коровы контрольной группы получали рацион согласно норм потребности, разработанных ВНИИФБиП (Физиологические потребности в питательных веществах и нормирование питания молочных коров., Боровск., 2001) . Рацион коров опытной группы был составлен с учетом субстратной характеристики обменной энергии и аминокислотного состава обменного протеина (приложение 1,2). Коровы обеих групп получали одинаковое количество обменного протеина, а в опытной группе валовой и обменной энергии было на 6% меньше. В составе ОЭ рационов опытной группы больше содержалось ВЖК (на 40%), аминокислот и глюкозы (на 3-5%) и меньше ЛЖК на 8%. Это обеспечивало поступление в организм коров, при меньшем содержании комбикорма в рационе (на 10%) одинаковое количество аминокислот и глюкозы (табл.50, 51).
|
|
|
Таблица 50
Образование субстратов в пищеварительном тракте коров по периодам опытов из кормов рациона (г)
| Периоды опыта, день лактации | Образованные субстраты, г | |||||
| ацетат | пропионат | бутират | глюкоза | аминокислоты | ВЖК | |
| 1 период, 25 –й день | 3034,1 | 1523,0 | 688,4 | 601,1 | 1158,7 | 331,9 |
| 2 период, 55 –й контроль | 3579,4 | 1718,2 | 820,8 | 756,3 | 1709,0 | 415,6 |
| 2 период, 55 –й опыт | 3224,2 | 1550,5 | 778,4 | 763,8 | 1725,5 | 518,7 |
| 3 период, 85 –й контроль | 3853,6 | 1939,3 | 926,6 | 872,0 | 1740,6 | 349,5 |
| 3 период, 85 –й опыт | 3288,0 | 1621,6 | 774,2 | 797,6 | 1741,8 | 514,0 |
| 4 период, 115-й день | 3755,2 | 1811,9 | 880,2 | 690,6 | 1573,4 | 337,7 |
Состав обменного протеина у коров опытной группы обеспечивал потребность коров в незаменимых аминокислотах, а в контрольной обеспеченность составляла 80-90%, за счет разного состава комбикормов. Ранее проведенные исследования показали, что увеличение в составе ОЭ энергетической доли ВЖК приводит к повышению эффективности молокообразования. Поэтому рацион коров опытной группы содержал на 4,8 МДж ОЭ меньше. На 55-й, 85-й и 115-й день лактации проводили балансовые опыты по изучению переваримости в преджелудках и кишечнике, обмена энергии, определяли поглощение метаболитов крови молочной железой и исследовали гормональный статус. С 85-го по 115-й день лактации коровы обеих групп получали идентичные рационы.
Таблица 51
Образование субстратов в пищеварительном тракте коров по периодам опытов из кормов рациона (% по энергии от суммы всосавшихся субстратов)
| Периоды опыта | Образованные субстраты, % | ||||||
| ацетат | пропионат | бутират | глюкоза | аминокислоты | ВЖК | ||
| 1 период, 25 –й день | 30,61 | 21,86 | 11,85 | 6,80 | 19,71 | 9,17 | |
| 2 период, 55 –й контроль | 29,11 | 19,89 | 11,39 | 6,90 | 23,45 | 9,26 | |
| 2 период, 55 –й опыт | 26,99 | 18,47 | 11,12 | 7,17 | 24,36 | 11,90 | |
| 3 период, 85 –й контроль | 29,49 | 21,12 | 12,10 | 7,48 | 22,47 | 7,33 | |
| 3 период, 85 –й опыт | 27,05 | 18,98 | 10,87 | 7,35 | 24,17 | 11,59 | |
| 4 период, 115 | 30,81 | 21,15 | 12,32 | 6,35 | 21,77 | 7,59 | |
После того как группы коров стали получать различные рационы, молочная продуктивность в опытной группе стала возрастать по сравнению с опытной. На пике молочной продуктивности удой в опытной группе составил 28,9±0,87 кг, а в контрольной 27,6±1,63 кг (4,7%). На 55-й день разница в продуктивности составила 15,7%, на 85-й-12,5%, на 115-й день при идентичных рационах разница сохранилась (12,5%) и даже на 153-й день (19,6%) в пользу опытной группы (рис.9).
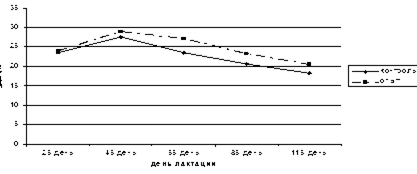
Рис.9. Молочная продуктивность коров в начале лактации.
Коровы контрольной группы в течение двух месяцев лактации теряли по 15 кг живой массы, а затем восстанавливали ее по 0,63кг в сут в течение третьего месяца и по 0,13кг в четвертый. Коровы опытной группы за второй месяц потеряли всего 5,5 кг, а восстанавливали массу по 0,4кг в сут за третий месяц и по 0,06кг за четвертый (рис.10). Таким образом, динамика изменения живой массы коров опытной группы была более сглаженной.

Рис.10 Динамика изменения живого веса коров в опыте
Установлено, что на 55-й день в крови коров обеих групп концентрация инсулина в сыворотке крови коров существенно возрастала, как после первого, так и после второго приема корма (рис.11). Это свидетельствует, что к концу второго месяца лактации наступила активация функциональной активности инсулярного аппарата поджелудочной железы, что оказало влияние на перераспределение потоков метаболитов в инсулинзависимые органы и ткани лактирующих коров. Отмечено также существенное снижение концентрации трийодтиронина в крови коров, что свидетельствует о торможении мобилизации жировых депо. В то же время если у контрольных животных выброс инсулина после приема корма превышал исходный уровень в 2,6 раз, то у опытных – только в 1,38 раза.

Рис. 11 Динамика концентрации инсулина в сыворотке крови коров
Указанные различия в темпах мобилизации у коров контрольной и опытной групп связаны с разным составом обменной энергии, в основном за счет большей долей ВЖК в опытной группе. В результате в артериальной крови у них всегда наблюдали повышенное содержание триацилглицеролов по сравнению с контрольными животными (рис.12).
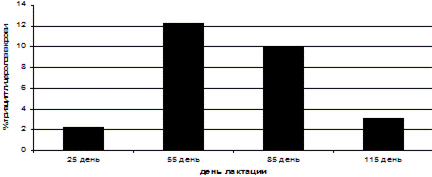
Рис. 12 Процентное отношение среднесуточной артериальной концентрации триацилглицеролов в опытной группе к контрольной.
В предыдущих исследованиях нами показано, что по ходу лактации у коров снижается использование ВЖК в энергетическом обмене и увеличивается доля использования ацетата. В текущих исследованиях отмечена аналогичная закономерность по использованию этих субстратов и молочной железой. По ходу лактации поглощение триацилглицеролов и НЭЖК снижается, а ацетата и бутирата увеличивается, в основном за счет изменения эффективности поглощения этих метаболитов.
За счет более равномерной мобилизации жировых депо из организма коров опытной группы, доля неэстерефицированных жирных кислот (НЭЖК) от суммы поглощенных молочной железой липидных компонентов во все периоды лактации была на уровне 25%, в то время как у коров контрольной группы достигала на 55-й день 35%, а к 115-му дню снижалась до 15% (рис.13).
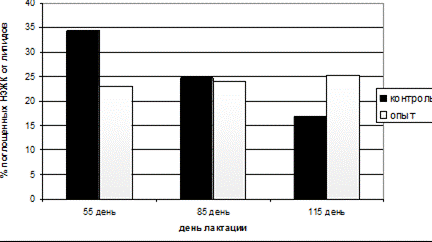
Рис.13 Относительное поглощение НЭЖК молочной железой от суммы поглощенных триацилглицеролов и НЭЖК
При этом жирнокислотный состав молока коров контрольной и опытной групп не различался, т.к. увеличенное поглощение триацилглицеролов молочной железой у коров опытной группы компенсировалось меньшим поступлением НЭЖК и большим выделением липидов с молоком.
Балансовые обменные опыты показали, что в период 85-го дня лактации наблюдалась уже отложение энергии в тканях (11,6 ± 0,55 МДж/сут), при этом также избыток обменного белка шел на отложение в тканях (63,9±12,7). На торможение липолиза и активацию липогенеза указывает повышение дыхательного коэффициента у коров контрольной группы до 0,88 и сохранения у коров опытной группе на уровне первого месяца лактации -0,78.
За счет использования в рационах коров опытной группы белковых кормов с разным набором незаменимых аминокислот в нераспадаемом протеине, в химусе, поступающем в кишечник содержание метионина было выше на 42%, лейцина на 17%, фенилаланина на 17%, лизина на 24,6%, гистидина на 76,3%, чем у коров контрольной группы (табл.52).
Таблица 52
Содержание незаменимых аминокислот в химусе двенадцатиперстной кишки, % от суммы кислот
| Аминокислоты | Предварительный период -25-й день лактации | Контроль 25-85-й день лактации | Опыт 25-85-й день лактации | Заключительный период 85-115-й день лактации |
| Метионин | 1,67 | 1,97 | 2,80 | 2,24 |
| Лейцин | 7,94 | 7,47 | 8,75 | 7,60 |
| Фенилаланин | 5,38 | 5,33 | 6,37 | 5,47 |
| Лизин | 6,60 | 6,00 | 7,48 | 6,13 |
| Гистидин | 1,98 | 1,44 | 2,54 | 1,56 |
В результате этого в составе обменного белка концентрация этих аминокислот была также увеличена –метионина на 32,6%, лейцина на 2,1%, фенилаланина на 13,4%, лизина на 21,4%, гистидина на 74,4% (табл.53).
Таблица 53
Дата добавления: 2018-11-24; просмотров: 253; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!