Сатысы: май кышкылынын активтенуі.
Май қышқьшдары мейлі ол липогенезге жұмсалсьш немесе бетта-тотығуға ұшырасып, алдымен міндетті турде рёаццияласу қабілетіне ие активті формаға айналу керек. бета-тотығуға тусетін май кышқылдары АТФ-ке тәуелді арнайы ацил-КоА-синтетаза әсерінен кофермент А-мен (НSКоА) әрекетгесіп ацил~SКоА тузеді. Реакция барысында АТФ АМФ пен пирофосфатқа ыдырап, бөлінген энергия ацил~SКоА-дағы макроэргиялық байланыстың тузілуіне жұмсалады. Сол себепті ацил~SКоА-ны активті май қышқылы деп те атайды.
2-сатысы: ацил~SКоА-ның цитоплазмадан митохондрияға тасы маддануы
Ұзын тізбекті май кышкылдарыньщ активтенуі цитоплазмада өтетіндігіне қарамастан, олардың бетта -тотығуы митохондрия ішкі мембранасының матрикске караған бетінде өтеді. Митохондрия мембранасы ұзын тізбекті май қышқылдарын өткізбейді. Сондықтан да тузілген ұзын тізбекті Ацил-КоА - карнитин деп аталатын арнайы тасымалдаупының көмегімен цитоплазмадан митохондрияға тасымадданады. Митохондрия мембранасының сыртқы бетінде карнитинацилтрансфераза I әсерінен ацил ~КоА карнитинмен реакцияға түсіп ацилкарнитинге айналады, коферментА-ны бөліп шығарады. Түзілген ацилкарнитин митохондрияның ішкі мембрана сына жеңіл өтеді. Митохондрия ішіне тускен ацилкарнитин митохондрия ішкі мембранасының матрикске қараған бетінде орналасқан карнитинацил- трансфераза II әсерінен кофермент А-мен (НЗКоА) реакцияланып, қайтадан Ацил ~КоА мен карнитин түзеді. Ацил ~КоА бетта-тотығуға ұшырайды. Қыска тізбекті май кышкылдары карнитінге тәуелсіз турде митохондрия ішіне еніп,активті формаға айналады.
|
|
|
3-сатысы: Май кышқылының тізбегінің кысқаруы.
Карнитиннің көмегімен митохондрия матриксіне түскен ұзын тізбекті ацил~SКоА немесе митохондрия ішінде түзілген кысқа тізбекті ацил~SКоА “май кышқылдарының оксидазасы” деп аталатын мультиферменттік комплекстің катализімен жүйелі түрде ацетил ~SКоА-ны бөліп шығарып, көміртек тізбегі екіден кысқарып отырады. Әрбір ацетил~SКоА молекуласы- ның түзілуі 4 сатылы ферментативтік реакциядан түрады, оны (бета-тотығудың циклі деп атайды. Бұл циклде алдымен коферменті ФАД болып келетін ацил~SКоАдегидрогеназа ферменті ацил~SКоА-ның 2(альфа)- және 3(бетта)- көміртектерінен бір-бірден сутегін бөліп шығарып С3- мен С3-көміртегінің арасында кос байланысы бар косылыс дегидроацил-SКоА түзеді. Босаган екі сутегі ФАДН2 түрінде тыныс алу тізбегіне беріліп, эндогенді су және энергия (жылу+2АТФ) түзеді. Екінші реакцияда, дегидроацил-SКоА сәйкес гидратазаның катализдеуімен бір молекула суды қосып алып, қос байланыс үзіледі де ,бетта-гидрокси-ацил~SКоА-ға айналады. Үшінші реакцияда, коферменті НАД болатын бета-гидроксиацил~SКоАдегидрогеназа деп аталатын пиридинферментінің әсерінентүзілген (бетта-гидроксиацил~SКоА-ның бетта-көмір- тегіндегі сутек атомдары бөлініп цығып, бета-кетоацил~SКоА түзіледі. Бөлініп шыққан екі сутекгі кофермент НАД косьп алып, онан әрі тыныс алу тізбегіне тасымалдайды. Пиридинферментгерінің тьнысалутізбегіндегі орналасуреті бойьшша екі сутек отгегіне тасымалдану барысыңда 3 молекула АТФ синтезін қамтамасыз етеді. Төртінші тиолазды реакция, яғни циклдің соңғы реакциясында бетта-кетоацил~SКоА тиолазаньң эсер етуімен С2 мен С3-көміртегінің арасындағы байланыс үзіліп, ацетил~SКоА-бөлініп шығады. Сонымен бір уақытта екі көміртегіне кысқарған ацил~SКоА түзіледі,ол реакцияға қайтатүседі. Осылайша цикл төрт көміртекті фрагменті-бутирил-КоА -түзілгенше кайталана береді. Яғни, ең сонында бутирил-КоА екі ацетил-SКоА түзеді. Түзілетін ацетил~SКоА- ның саны мен бетта-тотығудың цикл саны май қылқылдарының көміртек санына (қос байланыстьщ бар-жоқтығьна, сондай-ақ көміртек тізбегінің тақ санды немесе жүп санды болуьна) қарай әртүрлі болады.
|
|
|
|
|
|
Ацетил-SКоА Кребс циклінде 2С02-ге дейін тотығып, энергия көзі ретінде 1 молекула АТФ және 8 активті сутегін (ЗНАДН2+ФПН2) береді. Активті сутектері -ЗНАДН2 мен ФПН, - онан ары тыныс алу тізбегі арқьлы оттекке тасымалданып, эндогенді су және энергия түзеді. Бөлінген энергияның біраз бөлігі жылуға айналады, ал біраз бөлігі тотығудан фосфорлану аркылы АТФ синтезіне жүмсалады.
Май қышқылдарының бетта-тотыгуында тузілетін энергия молшері.
Митохондриядағы қаныккан май қышқылдарының (бета-тотығуына пальмитатты С15Н31СООН мысал ретінде келтіреміз:
а)пальмитин қышкылының көміртек саны 16, сол себепті түзілетін
активті сірке қьшқылының (АСҚ) саны АСҚ = 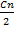 (Сп-көміртек саны);
(Сп-көміртек саны);
16/2=8 болады. Әрбір АСҚ Кребс циклі -» тыныс алу тізбегі —»тотығудан фосфорлану аркылы 12 АТФ түзетіндіктен, 8АСҚ толық тотыққанда жалпы жиыны 8x12=96 АТФ түзіледі; .
|
|
|
б)пальмитин кышқылының бетта-тотығу цикл саны= -1; 16/2-1=7 бола-
тындықтан, 7 циклде 7ФПН2 және 7НАДН2 түзіледі (бір циклде 1НАДН, және 1ФПН2 түзіледі). Тыныс алу тізбегінде реакция пиридинферменттері- нен (НАДН2) басталса ЗАТФ синтезіне, ал флавинферменттерінен (ФПН,) басталса 2АТФ синтезіне жететін энергия бөлінетіндіктен жалпы жиыны:
7ФПН2 х 2АТФ =14АТФ
7НАДН2 х ЗАТФ = 21АТФ; 14+21=35АТФ түзіледі (немесе бір циклде 1НАДН2+ФПН2= 3+2= 5 АТФ түзіледі; ал 7 циклде 5х7цикл=35АТФ).
Сонымен, 96+35=131АТФ түзіледі
в)митохондрия сыртында 1 молекула пальмитин қышқылын активтендіру үшін 1АТФ (АТФ->АМФ+ФФ) жүмсалады. Сондықтан барлыгы 96АТФ+35АТФ-1АТФ= 130АТФ болады.
70. Кетон денелерінің синтезі, реакциялар ретін сөзбен көрсету, өтетін орны. ГМГ-КоА мен кетон денелерінің формуласын жазу. Кетон денелерінің ағзада пайдаланылуы. Кетон денелерінің тотығуының энергиялық балансы. Кетогенездің күшеюінің себептері, осы құбылыстың негізгі салдарлары.
КЕТОН ДЕНЕЛЕР1ЖӘНЕ ОЛАРДЫҢ ТҮЗІЛУІ
Кетон денелерінің синтезі-кетогенез. Кетон денелері деп бетта-гидрокси- бутират ( бетта-гидроксимай қышқылы), ацетоацетат және ацетон - осы үш қосылысты атайдьі. Қалыгпы жағдайда <0,2 мМоль/л көлеміндегі кетон денелері қан кұрамында үнемі кездесіп отырады. Бірақ организмде май кышкылдарының тотығуын шамадан тыс күшейгетін кез-келген себептер кетонденелерінің артық мөлшерде түзлуіне әкеліп соғады. Ацетил~SКоА (активті сірке қышкьшы-АСҚ) кетон денелерінің негізін салуш қосылыс. Кетон денелері негізінен бауырда, аз мөлшерде бүйрекге де синтезделеді. Кетон денелерінің түзілуіне қатысушы ферменттердің барлығы митохондрия ішіне шоғырланғандықтан, олардың синтезі де осы жерде өтеді . Кетон денелерінің синтезі келесі сатылы реакциялардан тұрады.
Бірінші реакция:
2 молекула ацетил-SКоА 3-кетотиолаза әсерінен өзара конденсация- ланып, НSКоА-ны бөліп шығарады да ацетоацетил~SКоА түзеді.
Екінші реакция:
Түзілген ацетоацетил~SКоА (бетта-гидрокси- бетта-метилглутарил~SКоАсинта- заның өсерінен тағы бір активті сірке қышқылымен конденсацияланып бетта- гидрокси- бетта-метилглутарил~SКоА-ға (ГМГ~SКоА) айналады. Бұл реакция кетон денелерінің синтезіндегі бірінші қайтымсыз реакция болып есептеледі. ГМГ~SКоА кетон денелерінің тікелей негізін салушы қосылыс.
Үшінші реакция:
ГМГ~SКоА онан әрі бауыр жасушаларының митохондрия ішіңде ғана болатын ГМГ~SКоА - лиазаның әсерінен активті сірке кышқылын бөліп шығарып ацетоацетат (ацето- сірке кышқылы) деп аталатын бірінші кетон денесін түзеді. Ацетоацетат басқа екі кетон денелерінің тікелей негізін салушы кетон денесі.
Ацетоацетат митохондрия ішкі мембранасымен берік байланысқан бетта- гидроксибутират - дегидрогеназаның әсерінен НАДН2 арқьілы тотыксьз данып бетта-гидроксибутиратқа (бетта-гидроксимай қышқылы) айналады. бетта-гидроксимай қышқылы екінші кетон денесі. Келесі кетон денесі-ацетон ацетоацетат декарбоксилдену аркылы түзіледі. Бұл реакция ферменттің қатысуынсыз қайтымсыз жүретіндіктен, ацетонның түзілуі тек ацетоацетат ерекше көп түзілгенде ғана байқалатын қүбылыс. Бүған коса ацетон адам организміңце энергетикалық субстрат ретінде пайдаланылмай өзгеріссіз өкпе арқылы ауаға бөлінеді. Сол себепті қантты диабеттің ауыр түрінде науқастын демінен алмұрттың иісі тәрізді иіс иығады.
Бауыр кетон денелерін энергия ретінде пайдаланбайды. Кетон денелері бауырдан басқа тіндерде (бұлпық ет, бүйрек және т.б.) энергия көзі ретінде пайдаланылады.
Кетон денелерінің тотығуы
Кетон денелері тотыкқаңда органдар мен тіңдерде 24 немесе 27 АТФ түзіледі. бетта-гидроксимай қышқылы алдымен НАД-тәуелді дегидрогеназа ферменгінің қатысуымен ацетоацетатқа дейін тотығады. Ал ацетоацетат НSКоА- мен әрекетгесіп ацетоацетил~SКоА тузеді. Түзілген ацетоацетил- НSКоА- мен әрекетгесіп тиолазаның катализдеуімен екі АСҚ- тузеді. Әрбір АСҚ Кребс цикліңде, биологиялық тотығу тізбегінде, тотығудан фосфорлануда толық тотығып 12 АТФ түзеді. Екі молекула АСҚ 24 молекула АТФ береді. Бетта –гидроксимай қышқылы тотыққанда тузілген НАДН2. Ал косымша 3 молекула АТФ тузуге жеткілікгі энергия береді, сондықтан 27 молекула АТФ береді.
Жиыны: бетта-гидроксимай кышкылы тотыкканда 3+24=27АТФ түзіледі. Ацетоацетат тотыкканда 2АСҚ түзіледі, ол 24 молекула АТФ береді.
71.Холестериннің синтезі, реакцияларретінсөзбенкөрсету, үрдістіңөтетінорны. ГМГ-КоА мен холестеринніңформуласынжазу. Мевалонқышқылының ГМГ-КоА-дан түзілуреакциясынжазу. Осы реакцияныңреттелуі.
Холестерин синтезі бауырда, ішекте, теріде, бүйрек үсгі бездерінде, үрықта, плацентада, аналық бездерде жүреді. Холестериннщ 80%-ы бауырда түзіледі. Холестерин синтезіүш сатыдан тұрады.
I сатысы. Ацетил-КоА-ның 3 молекуласынан мевалон қышқылының түзілуі.
АСҚ цитоплазмаға цитрат түріндетүседі. Холестеринсинтезінщалғашқыреакциялары кетон денелерісинтезініңреакцияларыменүқсас, бүлреакциялар ми- тохондриядажүреді (кетон денелерінің синтезіндегі ГМГ-КоАтүзілгенгс дейінп реакцияларды караңыз).
Түзілген ГМГ-КоАцито плазмада мевалонатредуктаза ферментінің (коферменті НАДФНр әсерінен мевалонатқа тотықсызданады. Бұл реакция холестерин синтезіндеп реттеуші реакция болыгі табылады, осы үрдістің жылдамдығын реттейді.
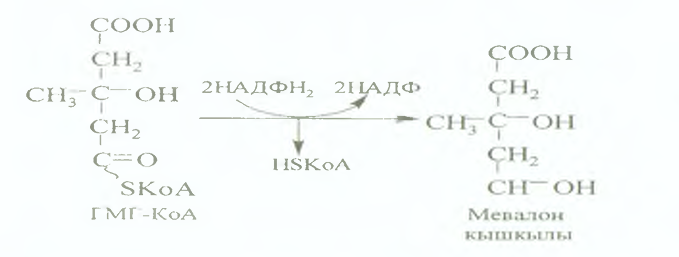
II сатысы. Мевалонаттың 6 молекуласынанскваленніңтүзілуі.
Осы сатыдамевалонаттан 5 көміртегі бар активтіизопрендер (1 молекула мевалонаттүзілуүшін 3 молекула АТФ қажет) түзіледі. Оларконденсацияланып, құрамында 30 көміртегі бар скваленгеайналады.
III сатысы. Холестеринніңтүзілуі.
Скваленненланостериндегенциклдікқосылыстүзіледі. Олмодификацияғаұшырап, 3 көміртекато- мынжоғалтып, холестерингеайналады.
Сквалентүзілуінедейінгіаралықреакцияларцитозольдежүреді. Скваленжәнебасқа да өнімдер суда ерімейдіжәнемикросомалдытотығуферменттерініңқатысуыменэндоплазматикалықретикулумныңмембраналыққабатындатүзіледі.
Эстрогендер ГМГ-КоА-редуктазаныңактивато- рыболыптабылады. Тағамқұрамындағы холестерин мөлшерікөпболғанжағдайдабауырдағы холестерин синтезітежеледі.
72.Тағамнәруыздары. Ағзаүшіннәруыздардыңмаңызы. Нәруыздаралмасуыныңерекшеліктері. Азотты баланс, оныңтүрлері. Нәруыздардыңқорытылуы. Нәруыздардыңқорытылуынақатысатынферменттер. Нәруызалмасуыныңбұзылысы – нәруызжетіспеушілігі (квашиоркор, маразм).
1. Белоктаркөмірсулар мен липидтерсияқтыорганизмдеқорретіндежиналмайды. Тек канплазмасынынбелоктары аз мөлшердебауырдақорғажиналып, же- дел қан кету жағдайындақанғагүседі. 2. Тіндердіңүнеміжаңаруы мен ферменттердің, гормондардың, биологиялыкактивгізаттардың (БАЗ) түзілуі, организмніңқалыптытіршілікәрекетіүыпнқүндыбелоктардыңжүйелітүрдетағамментүсігіотыруынқажететеді. Белоктардыңтапшылығыкезін- де организмдегормондардың, ферменттердің, БАЗ-ң синтезібұзылады. Егер белок тағамментүспесе, ондабелоктардыңжаңалануынатіршілікүшінмаңызызормүшелердің (ми, жүрек, бүйрек, бауыр) жәнебұлшықеттіңбелоктарыпайдаланылады. 3. Белоктарорганизмдеөтемаңыздыкөпсалалықызметтерді: реттеуші, тасымалдау, құрылымдық, катализдікжәнет.б.қызметтердіатқарадыбұлаталғанқызметтердімайлар мен комірсуларатқараалмайды.
Тағамдабелоктардыңжетіспеушілігіорганизмде, әсіресеосінкслежатқанорганизмде, жүктіліккезіндеауырзардапқашалдықтырады. Белоктардыңорганизмгеқажеттілігіорганизмніңжасына, энергия шығынына тәуелді болады:
- Ересекденісауадамның 1 кг денесалмағынаесептегендетәулігіне 0.8 г/кг белок;
- жаңатуылғаннәрестелерүшін- 2,0 г/кг;
-бес жасгағыбалаларғатәулігне - 1,0 г/кг жүйелігүрдетағамменгүсіпотыруықажет.
Белоктардың биологиялық құндылығы оның амин қышқылдық құрамына байланысты. Организмгеқұрамындабарлықсегізалмастырылмайтын амин қышқылы бар құнды белок түсіптұруықажет. Құндыбелоктыңхалықаралық«шарттыүлгісі» бар, бұлүлгібойынша белок құрамындағыалмастырылмайтын амин қышқылы 31,4%-ды (сүт, нан, жұмырткабелогынқосаалғанда) күрайды.
Белокқадегенқажеттілік энергия шығынына да тәуелдіекенінескеруқажет. Егер 10500 кДж энергия (ой еңбегі, механикаландырылғанеңбек) жұмсалса, онда 100-120 г белок қажет. Ал энергия шығыныәрбір2100 кДж артқансайын 1 Огбелоктанқосылыптұруытиіс.Организмгетүскен белок мөлшеріншжеткіліктілі- гінанықтауүшін«азотты баланс» гүсінігіенгізілген.
Азотты баланс - организмгетағамментүскеназоттың (белок ретінде) организмнензәр, нәжісарқылышығарылатыназогқа (мочевина, несепқышқылы, кре- атининжәнег.б.) аракатынасы. Осығанбайланыстыазоттыбаланстыүштүрге бөлуге болады: Он азотты баланс тағамарқылытүскен азот организмненэкскрецияланатыназоттанкөпболғандабайкалады. Бұл өсіпкележатқан организмде, жүкті әйелдерде байкалады.
Теріс азотты баланс организмге тағамарқылы түскеназоттың организмненбөлінген зәржәненәжісазотынан аз болғанда байқалады. Олқаргадамдар- да, емшектегібалаларға белок жеткіліксізмөлшердетүскенде, ашығукезінде, үлкенаймақтыкүйікте(белоктардыңсіңірілуібұзылады), тінбелоктарыныңыдырауыкезіндебайқалады.
Нольдік азотгы баланс (азотты тепе-тенлік) денісаудұрыстамақтанғанересекадамдардаболады.
Белоктардьіц ас қорытужолдарындақо-рытылуынақандайферменттерқатысады?
Белоктардыңқорытылуыалдыменасқазанда, сонансоңащыішекқуысында (қуыстыққорытылу), одан кейін ішектің қабырғалыққабатында жәнеішекэпителийінің жасушаларында қабырғалық қорытылу жүреді. Ауызқуысындапептидгидролазаларферментіжоқ, асқазандаэндопептидазалар (пепсин, гастрик- син) асқазансөліндегі тұзқышқылыныңәсерінен активтеніп, белоктарды полипептидтерге дейін гидролиздейді.
Ішектетағамбелоктарыныңгидролизденуінеэндопептидазалар (ұйқыбезі сөлінің пептидазалары: трипсин, химотрипсин, эластаза) қатысады, олардың әсеріненбелоктарполипептидтерге, ал ішексөліэкзопептидазаларының (аминопептидазалар, дигрипептидазалар) жәнеұйқыбезісөлінщкарбок- сипептидазасыәсеріненполипептидгер бос амин қышқылдарынадейіныдырап, сіңірілебастайды. Амин қышқылдарыәртүрліжылдамдықпенсіңіріледі. Амин қышқылдарыныңсіңірілуі АТФ энергиясыныңесебіненNa,К,АТФазажәне ү-глутамилтрансферазаферментініңкөмегіменекіншілікактивтігранспортарқылыжүреді. Амин қышқылдарынішекқуысынанішекжасушаларынатасымалдайтын бес түрлігасымалдаужүйесі бар:
бейтарап амин қышқылдарын аарналған;
қышқылдық амин қышқылдарынаарналған;
негіздік амин қышқылдарынаарналған;
циклді амин қышқылдарынаарналған;
цис, орн, арг, лиз амин қышқылдарынаарналған.
Амин қышқылдары негізінен екіншілік активті тасымалдаужолы:
1.тасымалдаушы АТФ-азалардыңқатысуымен;
2. ГГТ (ү-глутаминтрансфераза) қатысуыменсіңіріледі.Дижәнетрипепгидтер мен кейбірбелоктар (ботулизм токсині, анасүтініңантиденелері).
Нәруыз алмасуының бұзылыстары: ақуыз-энергня жетіспеушілігі (квашиоркор, алиментарлық маразм, ане- мия, кахексия, алиментарлық қортықтық).
«Квашиоркор» емшектен шығарылған бала" немесе басқа түсінікте - "қызыл бала" деген магынада. Ауру ерте балалық кезеңде және емшектен шығарғаннан кейін 5 жасқа дейін байқалады. Ауруга ұйқышылдық, селқостық немесе қозғыштық, өсуі мен дамуының баяулауы, салмағыньгң азаюьг, терісі мен шашының түсі озгеруі, беті, аягы, шабы қьгзғьглтьгм тартуы, депигментация, шашының агаруы, ет тінінің атрофиясы, диспепсиялық құбылыс жэне тұрақты түрде іш өтуі, алдьгмен ішкі мүшелерінің ісуі және кейінірек - беті мен аяқ- қолының ісуі, бауьгрыньгң үлкеюі, атторексия тән. Ауыр жағдайларда ақыл-есі ауытқиды, жұқпалы және паразиттік аурулар қосылады.
Алиментарлық маразм — атрофия, әбден жүдеу. Дене массасы әбден жүдегенге дейін томендейді, терісі әжімденеді, жеңіл қатпарланады. Іші созылып немесе кеуіп тұрады, ішектің нұсқасы (жобасы) байқалады. Негізгі алмасуы томендейді, дененің температурасьт томендеу боладьт, жиірек іш қатады, кейде аштықтан диарея болуы да мүмкін.
1.Нәруыз түзілуінің төмендеуі (инсулиннің тапшылығынан аминқышқылдарының трансмембраналық тасымалдануы бұзылуы, полисомалардың деградациясы, трансляция үрдісінің бұзылуы) ® гипераминацидемия, аминацидурия
2.Нәруыз ыдырауының әсерленуі ® теріс азоттық тепе-теңдік, гиперазотемия
3.Антидене түізуінің және организмнің жұқпаға төзімділігінің төмендеуі
73. Асқазан сөлі, асқазан сөлінің физикалық-химиялық қасиеттері, патология кезіндегі өзгеруі. Асқазан сөліндегі тұз қышқылының маңызы, патология кезінде өзгеруі. Патология кезіде асқазан сөлінің физико-химиялык қacиeттepi езгереді 1. Мөлшерінің артуы асказан селінің секрециясы жоғарылаганда немесе сфинктерді спазмасы және стенозы кезіде тағамның ішекке баяу тycyi кезінде, ал мөлшерінің азаюысекрециясы темендегенде, тағам ішекке тез тускенде, сфинктер толық жабылмағанда байқалады. 2. Иici. «Жагымсыз май» иici ұшқыш май қышқылдарының болуына және май кышкылдарының микроорганизмдер әсерін тотығуының күшеюіне байланысты.Шіріген Иiс микрофлора ферменттерінің әсерінен белоктардың асказанда шіруі кезіде, қатерлі iciK ыдыраганда, тағамның асказаннан imeKKe Tycyi бузылганда байкалады. 3. Tyci: патология кезіде асказан сөлінде ет қоспасының болуына байланысты ол сары немесе жасыл TycTi болады. Егерасказан сөлінде туз кышкылыжокболса, ондаолсаргыш түстеболады. Асказан сөлі түсініңсарыболуыбилирубиннің (етпигмент) болуынабайланысты. Ал жасылдау түсасказан сөліндееттіңболуына (туз кышкылы) байланысты. Асказанселшдеканболганда, оныңтyciкызылнемесе коңыртустіболады. Егерасказан сөліде туз кышкылы бар болса, ондаасказан сөлініңTyci коңырнемесе кофе тәріздіболады. Ce6e6i туз кышкылы канның кұрамындагыгемоглобингеәсер етіп, оны коңыртусті гематингетотыктырады. Ал асказан сөлінде туз кышкылыболмаса, ондақан болған кездеасказан сөлініңтyciқызылдауболады. 4. Асказан сөлінің консентрациясы да езгеруі мумкін. Асказан сөлі өте тұтқыр болуы мумкін, кейде оның кұрамынан белгілі 6ip бөлігін зерттеу ушін бөліп алуға болмайтындай тұтқыр болады. Шырыш мөлшерінің артуы асказанда кабыну үрдістерінің бар екенін көрсетеді Асқазан сөліндегі тұз қышқылының маңызы, патология кезінде өзгеруі. Бос тұз қышқылының дебит-сағаты (1 сагат ішінде бөлген бос HCI-ц мөлшерін )және тұз қышқылының дебит-сағаты (1 сагат ішнде бөлген жалпы қышқыл өнімдерінің мөлшері) деген көрсеткіштер бар. Тұз қышқылының дебит- сағаты асқазанның қышқыл түзу функциясын дурыс сипаттайды деп есептелінеді. Асказан сөлінің секрециялануында келесі патологиялық жагдайлар байқалады:
1. Гиперглогидрия - жалпы қышқылдық пен бос HCI мөлшерінің артуы (асказан және 12 елі ішектің жарасы кезінде). 2. Гипохлоргидрия - жалпы кышкылдык пен бос HCI мөлшерінің төмендеуі 3. Ахлоргидрия - бос HCI-ц болмауы, жалпы қышқылдықтың төмен болуы. 4. Ахилия - асқазан сөлі секрециясының және ферменттердің болмауы.
74. Қалыпты жағдайдағы асқазан сөлінің химиялық құрамы. Асқазан сөлінің қышқылдылық түрлері, қалыпты шамалары. Асқазан сөлінің патологиялық құрамдас бөліктерінің пайда болу себептері. Патологиялық компоненттерге сапалық реакцияларды атап өту.-
Қалыпты асқазан сөлінің химиялық құрамы.Асқазан сөлі 99,4-99,5% судан және 0,5-0,6% құрғақ қалдықтан тұрады. Органикалык заттардьң улесне 0,1%, бейорганикалык заттардың үлесне 0,4-0,5% тиеді Органикалык заттарын белоктар жэне органикалык кышкылдар кұрайды. Асказан сөлінің белоктары: а) ферменттер (пепсин, гастриксин, липаза); б) қорғаныш белоктары - м у ц и н д е р , олар асказанның кілегей кабатын жауып, оны туз кышкылы мен ферменттерді әсерінен қорғайды; в) гастромукопротеид немесе Каслдың ішкі факторы, ол В12 витаминін бузылудан сактап, оның aғзаға сіңірілуіне септігін тигізеді г) асказан сөлінің кұрамында органикалык кышкылдар (май кышкылы, сут кышкылы) өте аз мелшерде кездеседі Асказан cөлінің бейорганикалык компоненттерне: туз кышкылы (бос жэне белоктармен байланыскан), калий, кальций туздары, сульфаттар, аммоний, кышкылдык фосфаттар жэне баска заттар жатады.
Асқазан сөлінің патологиялық кұрамдас бөліктеріАсқазан сөлінің секрециясы бұзылганда, онда патологиялык курамдас бөліктер (кан, ет, сут кышкылы, ұшкыш май кышкылдары), ягни калыпты асказан сөлінде кездеспейтін немесе тек iздepi гана болатын заттар пайда болуы мумкін.
ӨТ. Кышкылдык төмендегенде немесе кышкыл мулдем жок болганда өт пайда болады. Бул жагдайда асказан сфинктерінің кызметі бұзылады. ол босаңсиды да. iшектегі заттар асказанга eрiп кетуі мумкін. Бұған сондай-ак сфинктердеп тыртықтық урдістерге байланысты оныңтығыз жабылмауы ықпал етуі де мумкін. Асказан сөлі туснің өттің әсерінен сары немесе жасыл туске боялуы ондагы туз кышкылыныңболуына байланысты. Егер туз кышкылы болса, онда асказан солі жасылдау тусті болады, өйткені билирубин биливердинге тотыгады.
ҚАН. Асқазан сөлнің Tyci кызыл туске езгерсе, оның кұрамында канның бар екенін керсетеді Кан асказанда травма болганда, ауыз куысыныц,Өңештің кілегейлі қабатынын травмасында, өңештің вена кан тамырларының варикозды кеңейю кезінде, мурыннан кан кеткенде, зонд жутканда болады. Бул жагдайда кан ұсак жіпшелер тәрізді ұйыған турде болады. Сонымен катар кан асказан жарасында, iciктің ыдырауы кезіде пайда болуы мумкін. Бұл жагдайда асказан сөлінің Tyci «кофенің» тусіндей болады, ce6e6i туз кышкылының әсернен гемоглобин қара коңыр тусті туз кышкылды гематинге тотыгады. Ахлоргидрия немесе ахилия жагдайында асказан сөлініңде кан болса, онда ол әр турлі кызыл тустерге боя алады.
Сүт қышқылы.Сут кышкылы асказанда асказанның катерлі iciri кезіде, ахлоргидрияда жзне асказаннан тагамның ішeKKe өтуі баяулаганда пайда болады. Наукастардың асказанында тагам турып калады да, көмірсулар ашып, сут кышкылы тузіледі
Ұшқыш май қышқылдары да асказан сөлінің патологиялык курамдас бөлктеріне жатады, олардың пайда болу себебі асказан сөлінің кышкылдытының темендеуі немесе туз кышкылыныңмулдемболмауы. Қалыпты асқазан сөлінің қасиеттері. Аш қарындағы асказан сөлінің мөлшері 50 мл-ден аспайды (балаларда 30 мл-ден аспайды). Калыпты асказан сөлі - туссіз, иіссіз, күшті кышкылдык реакция керсететі (pH-1,5-2,0; жаңа туган балаларда рН-4,0-5,0) суйыктык. Меншікті салмағы 1,008-1,010. Калыпты жагдайда тәулігіне 2 литрге жуык асказан сөлін бөледі.
75. Белоктардың ішекте шіpyi. Улы шіpy өнімдерін залалсыздандыру. Залалсыздану орны жане жолдары.
Ыдырамаған белоктар тоқ ішекте микроорганизмдер ферменттерінің әсерінен ыдырайды. Бұл жағдайда амин қышқылдары анағұрлым терең ыдырауға ұшырайды, бұл үрдіс белоктардың шipyi деп аталады. Белоктардың қорытылуы мен шipyi арасында келесі айырмашылықтар бар:
1. Қорытылу ас қорыту жолдарының өздерінің ферменттері әсерінен, ал шipy тоқ ішектің микрофлорасы ферменттерінің әсерінен журеді
2. Қорытылу амин қышқылдары тузілгенге дейін жүрсе, ал шipy кезінде амин қышқылдары ары қарай өзгерістерге ұшырайды.
3. Белоктардың қорытылуы кезінде амин қышқылдарының қоспасы түзіледі де, олар сіңіріледі, ал шipy кезінде аминдер және т.б. шipy өнімдері түзіліп, олар ағзадан шығарылады.
Амин қышқылдарының ыдырауы eкі жолмен жүреді:
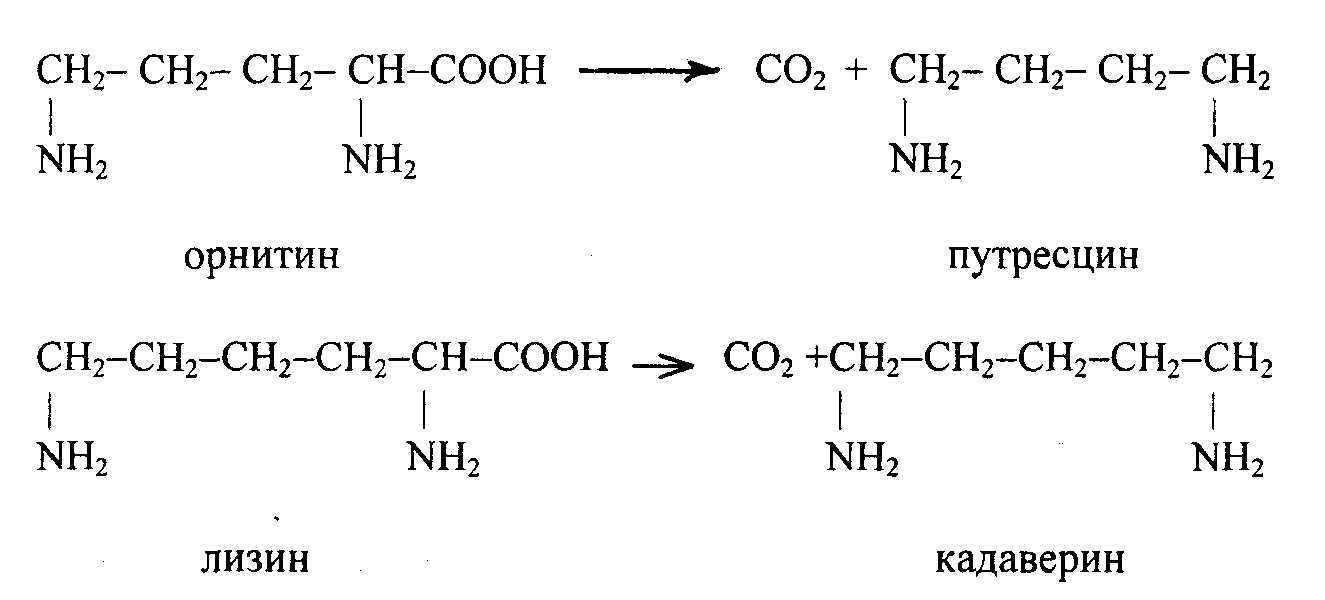
1.Декарбоксилдену , бұл кезде ациклді амин қышқылдарынан сәйкес аминдер түзіледі, орнитин мен лизиннен сәйкес путресцин мен кадаверин түзіледі, олардың улылыгы аз, сондықтан залалсызданбай, сол күйінде ағзадан шығарылады:
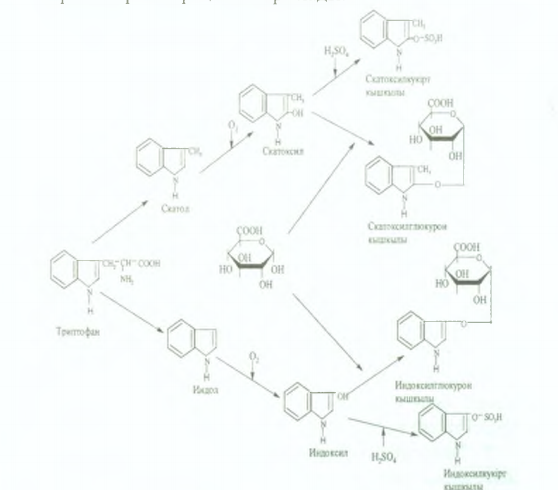
2. Циклді амин қышқылдары бүйір тізбегінің тотығуы мен бұзылуы арқылы ыдырайды. Тирозиннен крезол мен фенол түзілсе, триптофаннан скатол мен индол түзіледі.
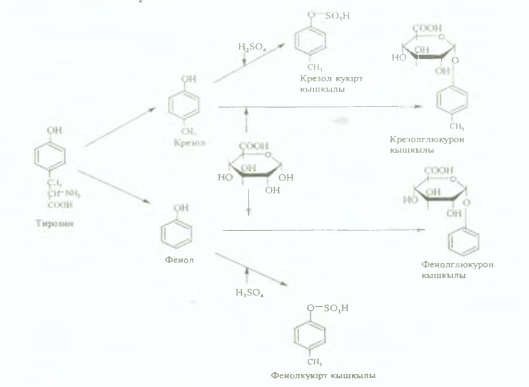
Бұл - улы заттар қан ағысымен бауырға түсіп, сол жерде күкірт жзне глюкурон қышқылдарымен конъюгациялану реакциялары арқылы залалсызданады. Бұл жағдайда усыз кукірт және глюкурон қышқылдарының эфирлері тузіледі, олар жұп қышқылдар деп аталады. Скатол мен индол алдымен гидроксилденіп, содан кейін ғана коньюгацияланады. Олар бүйрек арқылы зәрмен 6ipгe сыртқа шығарылады.
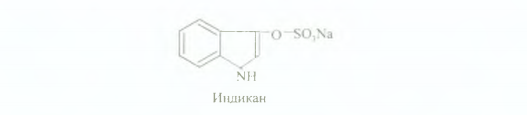
Клиникада қаннан жэне зәрден жануар индиканын аньқтайды, бұл индоксилкүкірт қышқылының калий немесе натрий тұздары. Формула
Индиканның мөлшері артса, онда ішектегі шipy үрдістері күшейеді , бұл iш қату және ішектің өткізгіштігі төмендегенде(іріңді ауруларда)байқалады. Ал бауырдың антитоксикалық қызметі бұзылған кезде индикан мөлшерінің төмендеуі байқалады.
76. Креатиннің түзілуі, тағдыры, маңызы. Жануар организмінде кейбір амин қышқылдарынан креатин синтезделуі мүмкін. Креатин синтезі 2 сатыдан тұрады:
1 сатысында буйректе аргинин мен глицин әрекеттесіп, гуанидин cipкe қышқылын (гликоциамин) түзеді, ол қан ағысы арқылы бауырға келіп түседі.
2 сатысы: гуанидин сірке қышқылы бауырда метилтрансфераза ферментінің қатысуымен (коферменті ТГФҚ (витамин Вс)) тотығудан метилдену (метил тобының доноры - метионин), креатин (метилгуанидин cipKe кышкылы) тузеді.
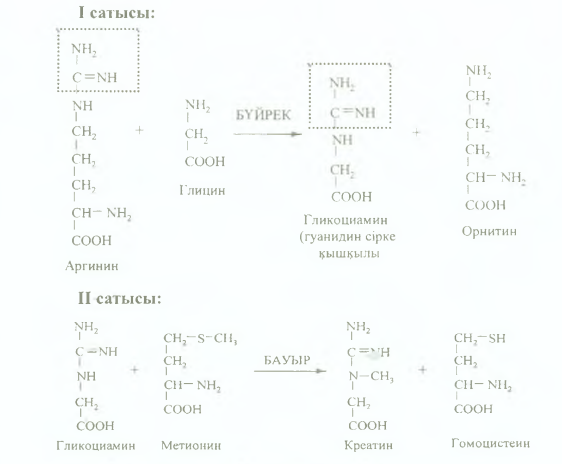
Түзілген креатиннің басым бөлігі бауырдан бұлшық етке (creas - ет) түседі де, АТФ-пен фосфорланып, макроэргиялық байланысы бар креатинфосфатқа айналады, бұл реакцияны креатинфосфокиназа (КФК) ферменті катализдейді.
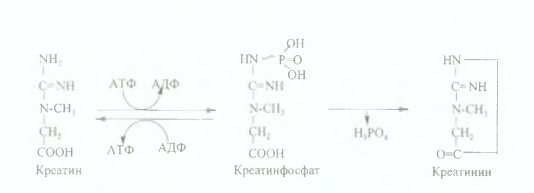
Креатинфосфат бұлшық ет жиырылуының химизміне қатысады, бұлшық етте АТФ-ң ресинтезделуіне бағытталған қайта фосфорлану (қайта эфирлену) реакциясына жұмсалады. Бұлшық еттің жұмысы (жиырылуы) және көптеген қайтымды қайта эфирлену реакциялары нәтижесінде креатинфосфаттың 6ip бөлігі ферментсіз дефосфорланып, креатинин тузеді Креатинин белоктардың алмасуының сонғы өнімi болғандықтан, ол қанға түсіп, бүйрек арқылы зәрмен сыртқа шығарылады.
77.Амин қышқылдарының тікелей емес тотығудан дезаминденуінің реакциясын жазу.Аминқышқылдарының азотсыз қалдықтарының пайдаланылуы туралы түсінік бер.
Жануар организмінде амин кышкылдарынын ыдырауы негізінен тотығудан дезаминдену аркылы жүреді.
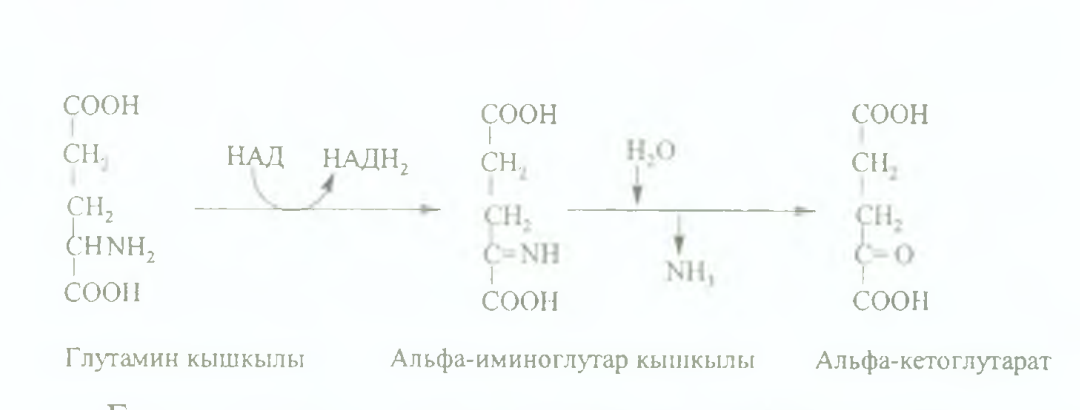
Амин қышкылдарының азотсыз қалдықтарының пайдаланылуы. Амин кышкылдарынын көміртек каңкасының катаболизмі нэтижесінде 7 түрлі өнім: ПЖК, ацетил- КоА, ацетоацетил-КоА, альфа-кетоглутарат, сукцинил- КоА, фумарат, оксалоацетат түзіледі.
Аталған заттар метаболизмге түciп, олардың кeйбipeyлepi глюконеогенезге, липонеогенезге жұмсалады немесе ҮКЦ және БТ тотығып, энергия беледі, көмірқышқыл газы мен су тузеді, кейбір жагдайларда алмастырылатын амин қышқылдарының түзілуіне жұмсалуы мүмкін.
Амин кышкылдарының азотсыз қалдықтарының пайдаланылуы.
- ПЖК: қайта аминдену реакцияларына, глицерофосфат аркылы липидтер түзілуіне жұмсалады, тотығудан декарбоксилдену аркылы АСҚ-на айналады, ол өз кезегінде ҮКЦ-не түciп, БТ тотығып энергия береді немесе холестерин, кетон денелері, БМК синтезіне жұмсалады.
- ҚСҚ - ҮКЦ-ң субстраты: кайта аминдену реакциясына, глюконеогенезге, БМК синтезіне, шөрнек механизміне жұмсалады.
- альфа-кетоглутарат: қайта аминдену, алмастырылатын амин кышкылдары түзілетін тотықсыздана аминдену реакциясына жұмсалады, ҮКЦ-ң субстраты болып табылады.
- фумар қышқылы: ҮКЦ-ң субстраты.
- сукцинил-КоА: ҮКЦ-ң субстраты, гем синтезіне жұмсалады.
- ацетосірке кышкылы: кетон денелері мен БМҚ синтезіне, АСҚ түзілуіне жұмсалады.
78.Ағзада аммиактың түзілуі және оны пайдалану жолдарын атап шығу. Амидтену және дезамидтену,аммонийгенез реакцияларын жазу, үрдістердің өтетін орны,маңызы.
Аммиак өте аз мөлшерде физиологиялық тітіркендіргіш, ал көп мөлшерде болса, улы зат,
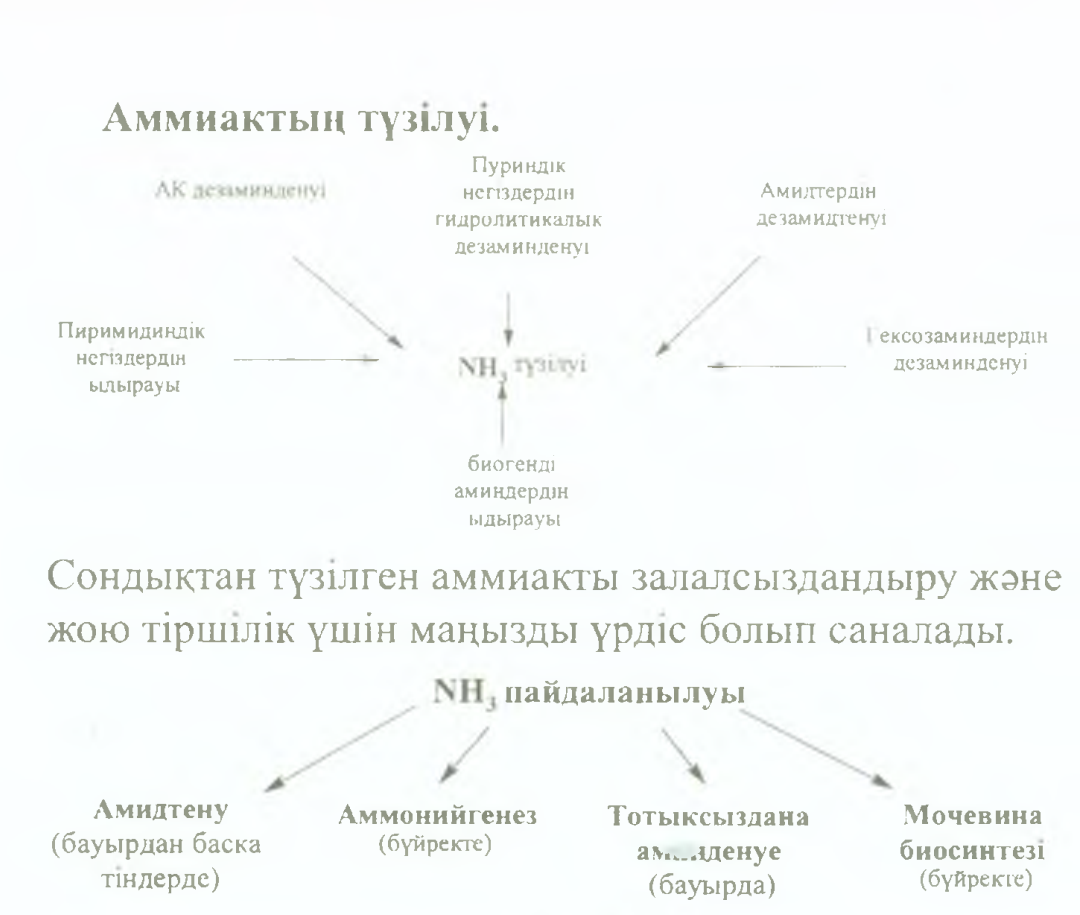
Аммиак амидтердің түзілуіне жұмсалады, бұл реакцияны амидтену деп атайды. Тіндерде түзілген амидтер (глутамин (глн), аспарагин (асн)) қан аркылы бауырға түсіп, осы жерде дезамидтеніп, глутамин кышкылы мен аммиакка дейін ыдырайды. Тура жэне Kepi реакциялар АТФ энергиясын жұмсай отырып журеді. Адам ағзасында амидтену үрдісі глутамин кышкылы аркылы, ал сирек жагдайда аспарагин кышкылы аркылы жүреді.
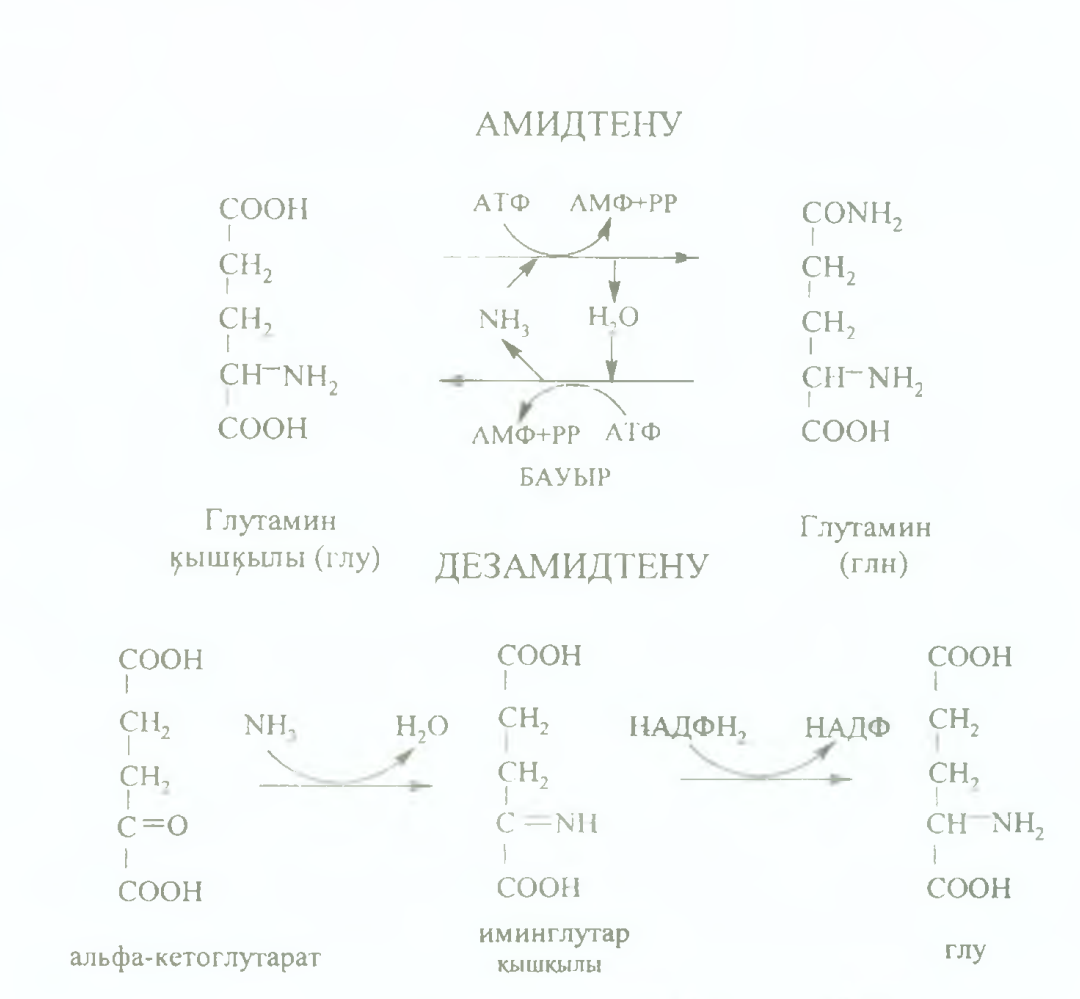
Амидтену реакциясы нәтижесінде түзілген амидтер аммиактың уакытша залалсыздандыру жолы және оның тасымалдану формасы болып табылады. Бүдан басқа амидтердің түзілуін аммиактың жұмсалу жолы деп есептеуге де болады, өйткені түзілген глутамин мен аспарагин белок синтезіне құрылыс материалы ретінде қолданылады, гексозаминдердің синтезі үшін амин тобыныц көзі және пуриндік, пиримидиндік негіздердің синтезіне қолданылуы мүмкін.
Түзілген аммиактың біраз бөлігі бүйректе қышқыл өнімдерді бейтараптауға жұмсалады да, организімнен аммоний тұздары түрінде зәрмен бөлініп кетеді.
Аммоний тұздарының түзілуінің мацызы зор: органикалык және бейорганикалык қышқылдардың артық мөлшері аммоний тұздары түрінде организмнен шығарылып, натрий, калий, кальций катиондарының сақталуына және қышқыл-сілтілік тепе-теңдікті сақтауға қатысады.
79.Мочевина синтезінің реакцияларын сөзбен жазу, үрдістің өтетін орны, маңызы. Мочевинаның формуласын жазу. Мочевина синтезі бауыр жасушаларында Кребс теориясына сәйкес өтеді (бұл теорияны 1932 жылы Кребс пен Хенселайт ашкан). Оны орнитин циклі немесе цитруллин теориясы деп те атайды. Мочевина синтезі 5 сатыдан турады: Карбомоилфосфаттыц тузшуі, митохондрияда журеді Карбомоилфосфат АТФ энергиясын пайдалану арқылы заттар алмасуының соңғы өнімдерінен (С 02, Н20 , NH3) тузіледі
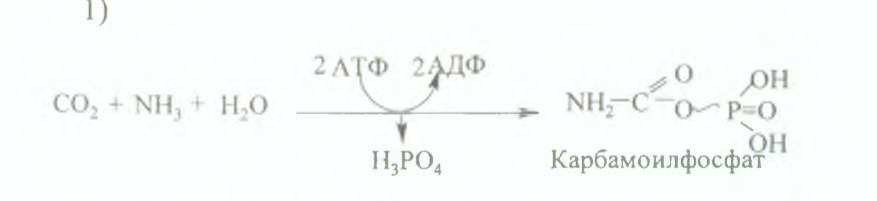
2) Цитрулиннің тузілуі (митохондрияда өтеді). қалган реакциялары цитоплазмада жұреді 3) Аргинин-янтарь кышкылының (аргининосукцинат) тузшуі, бұл саты үшінде АТФ молекуласы кажет, ол цитрулин аспарагин кышкылымен конденсациялану реакциясына жұмсалады, реакция нәтижесінде аргинин-янтарь қышқылы тузілді 4)Аргининнің түзілуі Аргинин-янтарь қышкылынан фумарат бөлініп кетеді, аргинин тузіледі 5) Мочевинаның тузілуі Соңғы сатысында аргиназа ферментінің әсерінен аргинин мочевина мен орнитинге ыдырайды. Мочевина алдымененольдыформадаболады да, соданкейінкето-форматқаайналады.
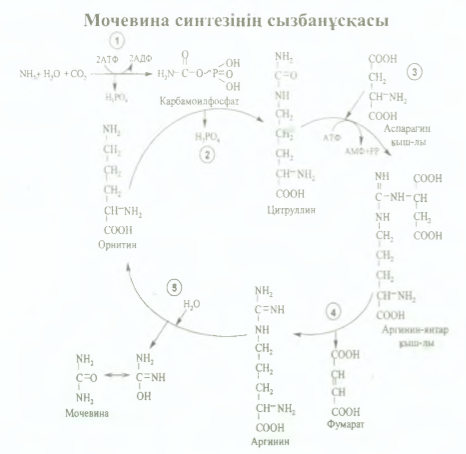
Мочевина молекуласындагы 6ip азот бос аммиактан, ал eкіншісі аспартаттан алынады. Аммиак көп мөлшерде глутаматтан (тотыгудан дезаминдену есебшен) түзіледі, сонымен катар аспартат та глутаматтан (оксалоацетаттыцтрансаминденушен) түзіледі Сонымен, мочевина молекуласындагы азоттың екі атомы да глутамин кышкылынан алынады, ал ол оз кезепнде аммиакты әр турлі амин кышкылдарынан жинайды. Мочевина синтезі аммиакты залалсыздандырудың негізі және түпкілікті жолы, ол - белоктар алмасуының соңғы онімі Мочевина синтезі- энергияны көп кажет ететін үрдіс, 6ip молекула мочевина тузу ушін 3 АТФ жумсалады.
80. Гемоглобин синтезі туралы түсінік беру, гемоглобин гемінің синтезінің реакцияларын атап шығу. Порфириялар туралы түсінік.
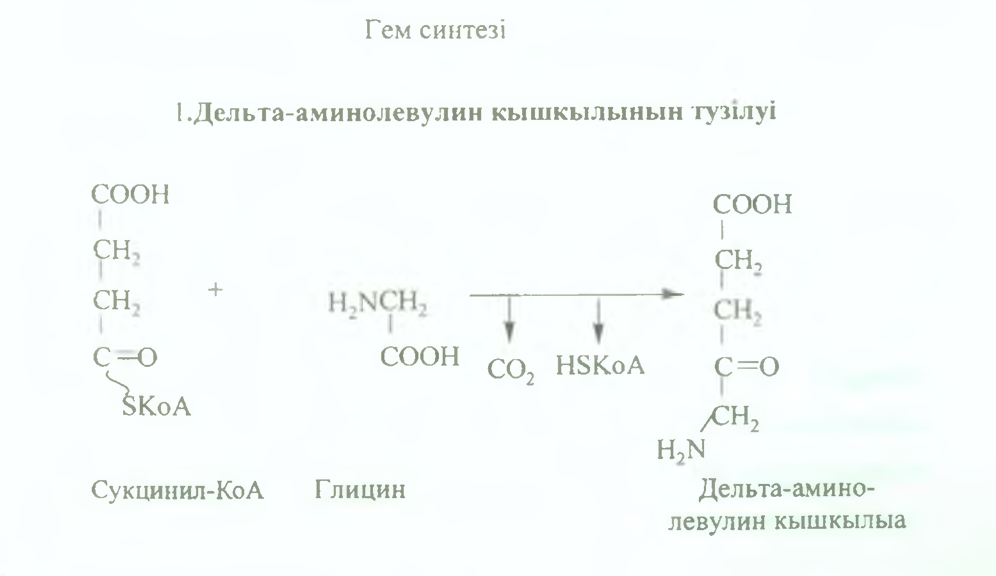
Гемоглобин сүйек кемігінің жасушаларында синтезделеді. Гемоглобин синтезіне қажет барлык құрамдас бөліктері қан арқылы түседі. Гемоглобиннің белоктык бөлігі жай белоктар тәрізді (АҚ матрицалык әдіспен) синтезделеді. Гемнщ синтезі түрлі фер- менттердің әсерінен бірнеше сатыда өтеді: 1. Алдымен дельта-аминолевулин қышкылы түзіледі. Бұл реакция митохондрияда аминолевули- натсинтетаза ферментінің әсерінен сукцинил-КоА мен глициннщ конденсациялануы нәтижесінде жүреді.
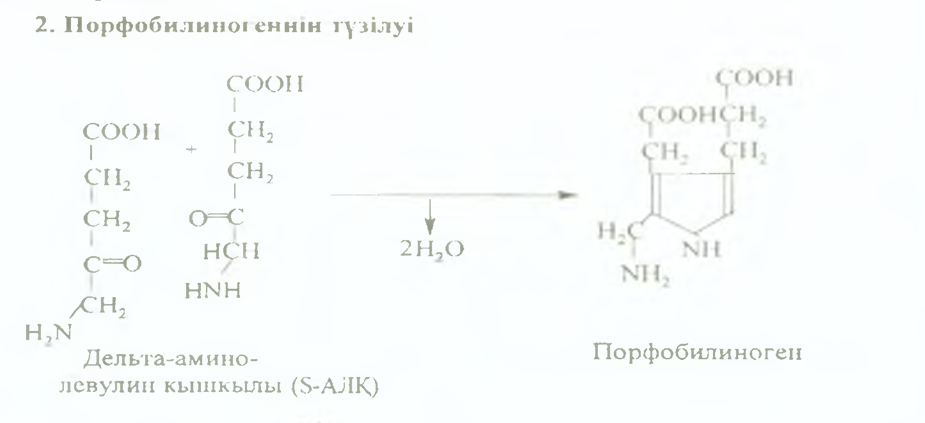
2. Келесі реакция цитоплазмада жүреді. Дельта аминолевулин қышкылының екі молекуласының кон- денсациялану реакциясы нәтижесінде иорфобилино- ген түзіледі.
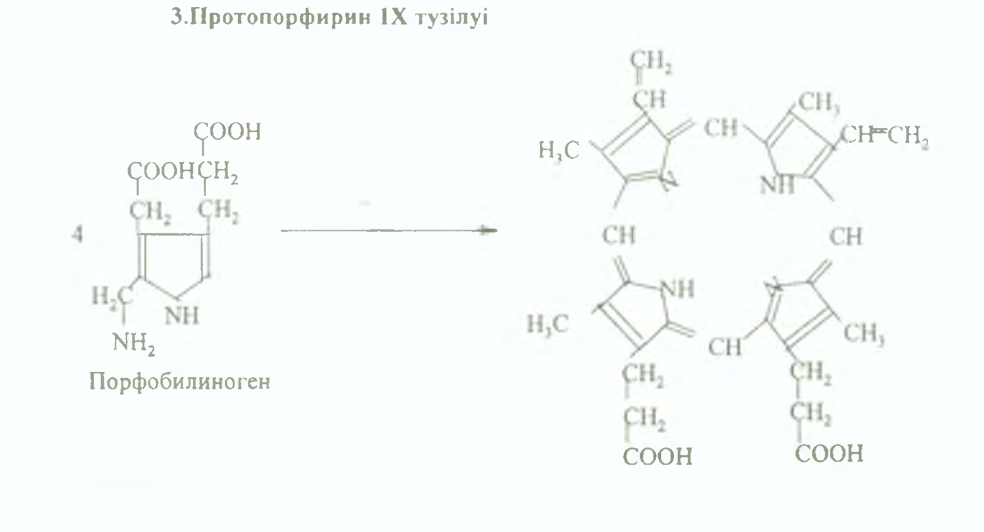
3. Содан кейін көп сатылы реакциялардың нәтижесінде порфобилиногеннің 4 монопирролды молекуласынан протопорфирин IX түзіледі, ол гемнщ тікелей негізін салушы болып табылады.
4. Протопорфирин-ІХ темір ионын (Ғе2 ) қосып алып (гемсинтетаза немес феррохелатаза ферметі әсерінен) гемді түзеді, ол барлық гемопротеидтердің биосинтезіне пайдаланылады. ПБГ синтезіне қа- тысатын екі ферменгтщ де активтілігі реттеледі, олардың активтілігі гем және гемоглобин арқылы тежеледі. Сондықтан гем артық немесе кем түзілмейді. Сондай-ақ гемоглобиннщ белоктық бөліп де белгілі мөлшерде түзіледі, өйткені оның синтезі тек гем болғанда ғана жүреді, түзілген полипептидгік тізбектер бірдей геммен байлаынсады. Гемнщ концентрациясы төмен болғанда, оның синтезі де бүзылып, гемоглобиннш түзілуі де баяулайды. 5. Глобиннщ әрбір полипептидтік тізбегі бір гем- ге байланысып, гемоглобиннш мономерін түзеді, төрт мономер бірігіп, гемоглобин түзеді.
Гемоглобиннщ атқаратын негізгі қызмегі оттекті өкпеден тіндерге, ал көмір қышқыл газын тіндерден өкпеге тасымалдау, қанның рН-ын тұрақты сақтауға қатысу. Гемоглобин өзінщ қызметін тек эритроциттің қүрамында ғана атқара алады. Эритроциттердің өмір сүру үзақтығы 110-120 күн. Содан соңэ ритроциттер гемолизге үшырайды.
81.Гемоглобин ыдырауы. Билирубиннің асказан-iшeк жолдарында өзгеске ұшырауы. Бос және байланысқан билирубин. Билирубиннің касиеттері
Эритроциттердің гемолизі кезінде гемоглобин қанға түседі де, гаптоглобин белогымен байланысып, гемоглобин-гаптоглобин комплексі түрінде макрофагты-моноцитарлық жуйе (ММЖЖ) жасушаларына: бауырдың Купфер жасушаларына, лимфа түйндерінің жасушаларына, көкбауырға, ішектегі пеер түйіндерінің жасушаларына тасымалданады.
Бул урдис гемнин биринши жане екинши пиррол сакиналарынын арасындагы метин копиршесинин тотыгын, узилуинен басталады да, вердоглобин тузиледи. Содан сон вердоглобиннен глобин мен темир болинеди де, сызыктык курылымды биливердин (жасыл тусти ) тузиледи. TeMip (Fe3+) бетта- глобулинмен байланысып, трансферрин туринде бауыр мен кокбауырга тасымалданады, сол жерде ферритин туринде корга жиналады. Глобин жай белоктар сиякты амин кышкылдарына дейин ыдырайды.
Биливердин НАДФН2 есебинен бос, коньюгирленбеген билирубинге тотыксызданады, ол суда еримейтин жане улы косылыс. Тикелей емес билирубин ММЖ жасушаларынан шыгып, альбуминдермен байланыса- ды да, гепатоциттерге тасымалданады.
Онын тикелей емес деп аталу ceбебi Эрлих диазореактивин косканда бирден кызыл тус бермейди, тек кан сарысуына ондагы белоктарды тунбага Tycipy yшин кофеин немесе спирт еритиндисин косканнан кейин кызыл туске боялады. Бауырдын Купфер жасушаларында гемоглобиннин ыдырау npоцeci вердоглобин, содан кейин биливердин тузилуинен басталады. Бауырда тикелей емес билирубин гепатоциттерде глюкурон кышкылынын бip немесе еки молекуласымен конъюгацияланып, моно- жане диглюкуронид билирубин тузеди. Бул билирубин коньюгацияланган, байланыскан,жэне тикелей деп аталады.
Тикелей билирубин суда жаксы ериди , усыз.
Биливердин мен тикелей билирубин от капшыгына жиналады, олар етке жасылдау тус береди, сондыктан ет пигменттери деп аталады. 0т ащы ишекке туседи, ет жолдарында тикелей билирубин глюкурон кышкылдарынан босайды да, кайтадан бос билирубинге айналады. Биливердин озгериссиз нажиспен болинип , нажиске тус береди, нажис пигменти болып табылады. Ишекте бос билирубин мезобилиногенге (уробилиноген) дейин тотыксызданады, онын бираз болиги какпа венасы аркылы бауырга кайта тусип , онда туссиз моно- жзне дипирролдарга дейин ыдырайды, олар буйрек аркылы зармен шыгарылады.
Мезобилиногеннин коп болиги ток iшeккe туседи, онда микроорганизм ферменттеринин асеринен стеркобилиногенге тотыксызданады. Стеркобилиногеннин бираз болиги геморроидальдык вена тамырлары аркылы буйрекке туседи. Зарде жарык пен ауанын асеринен стеркобилиноген стеркобилинге дейин тотыгады. Стеркобилин зарге сары тус береди, демек ол - зэр пигменти. Стеркобилиногеннин калган болиги ток ишекте тотыгады, жарыктын асеринен стеркобилинге дейин тотыгады. Стеркобилин биливердинмен биригип, нажис пигментин тузеди , нажиске коныр-жасыл тус береди. Емшектеги балаларда шириту процеске катысатын бактериялары жок болгандыктан, билирубин стеркобилиногенге айналмайды жане сол калпында сыртка шыгарылады. Соган сайкес нарестелердин нажисинин тусин биливердин мен билирубин (жасыл-сары) калыптастырады.
Балаларда эмбриональдык кезееннин алгашкы уш айында эмбриональдык гемоглобин тузиледи. Содан кейин ол фетальдык гемоглобинге (гемоглобин F), айналады, ол бала туылганга дейин болады. Туылганнан кейин алгашкы айда фетальдык гемоглобин биртиндеп полипептидтик тизбектердин курамы бойынша ерекшеленетин ересектердин гемоглобинине (гемоглобин А) айналады. Эмбриональдык жане фетальдык гемоглобиннин оттекке сайкестиги ересектердин гемоглобинимен салыстырганда жогары болып келеди.
От, зар, нажистин пигменттери.
Гемоглобин ыдыраганда от, нажис, зар пигменттери тузиледи.
От пигменттери биливердин (жасыл тусти), байланыскан билирубин (билирубин глюкурониди сары тусти). Оттин тyci осы пигменттредин аракатынасына байланысты.
Нажис пигменттери биливердин (жасыл тусти), стеркобилин (коныр TycTi).
Зар пигменти: стеркобилин.
Кан сарысуынын Tyci де белгили бip денгейде онда билирубиннин болуына байланысты. Калыпты жагдайда кандагы жалпы билирубин мелшери 8-20 мкмоль/л, ондагы тикелей емес билирубиннин улесине 75-тен 100%-га дейин , ал тикелей билирубинге 0 до 25%-га дейин келеди.
82.Сар ғ ыштану. Т ү рлер і , пайда болу ce б e пт epi, гемолизд і к, меха н икалы қ , паренхиматозды қ ж ә не жа ң а ту ғ ан н ә рестелерд ің сар ғ ыштануы кез ін де гі биохимиялы қ к ө рсет кі штерд ің салыстырмалы си паттамасы.
Кейбир ауруларда жалпы билирубин мелшеринин артуы, тикелей жэне тикелей емес билирубин арасындагы катынас озгеруи мумкин. Сондыктан кандагы жалпы билирубиннин , бос жэне тикелей билирубиннин молшерин аныктаудын диагностикалык манызы бар. Кандагы билирубин мелшеринин артуы онын тиндерде жиналуына,сонын ишинде тepi мен килегей кабаттарында жиналып, тиндердин сары туске боялуын тугызады. Бул куй саргыштану деп аталады, онын артурли себептери бар. Саргыштанудын бирнеше турлери бар:
1. Механикалык (обтурациялык) бауыр асты саргыштану оттин ишекке тусуинин бузылуымен байланысты. 0ттин шыгуын бузатын кез келген урдис (от-тас ауруы, уйкы безинин iciгi, от шыгару жолдарындагы кабыну урдистери жзне т.б.) саргыштану ceбeптepi болып табылады. Осынын натижесинде канда жалпы билирубин тикелей билирубин есебинен артады. Зардин тyci коныр (сыра таризди), ceбебi тикелей билирубин суда жаксы ериди , ал нажистин Tyci пигменттердин жок болуына байланысты аксур болады.
2. Гемолиздик (бауыр ycтi) саргыштану эритроциттердин шамадан тыс гемолизге ушырауынан туындайды. Ол сайкес емес канды куйганда, жукпалы ауруларда, кейбир химиялык заттармен уланганда жане т.б. жагдайларда байкалады. Канда бос билирубин коп тузиледи, оны бауыр залалсыздандырып улгермейди, сондыктан канда тикелей емес билирубиннин есебинен жалпы билирубин мелшери артады. Нажистин Tyci стеркобилиннин артык тузилуине байланысты коныр болады, зардин Tyci озгермейди, ойткени тикелей емес билирубин суда нашар еритиндиктен, зарге туспейди
3. Жана туган балаларда физиологиялык саргыштану байкалады. Онын каркындылыгы калыпты жагдайда екинши тауликте азая бастайды, ал eKi аптадан сон отеди, шала туган балаларда узагырак сакталады. Ойткени оларда фетальдык гемоглобиннин ересектер гемоглобинине алмасуы баяу журеди Бул саргыштану билирубиннин залалсыздандыратын фермент - глюкуронилтрансферазанын жеткиликсиз тузилуине жэне фетальдык гемоглобиннин ыдырауына байланысты.
1. Паренхиматоздык (бауырлык) саргыштану бауыр жасушаларынин вирустармен жане бауыр ушин улы болатын препараттармен закымданганда пайда болады. Бауыр жасушалары закымдангандыктан, бос билирубин заласызданбайды. Бауыр жасушаларынын закымдануы натижесинде канда тикелей билирубиннин де, тикелей емес билирубиннин де мелшери артады. Зардин Tyci зарде тикелей билирубиннин болуына байланысты коныр болады, уробилин болуы да мумкин. Нажис солгын (алсиз боялган) тусти болады, бул бауыр жасушаларынын закымдану дарежесиее тауелди. Бауыр аурулары кезинде зарде уробилин пайда болуы мумкин.
83.Нуклеопротеидтер алмасуы, қорытылуы.
Адам ағзасында нуклеопротеидтердің синтезі жақсы журеді Нуклеин кышкылдары заттар алмасуында маңызды роль аткаратындыктан, олардың синтезі улкен кызгушылык тудырады. Бул урдіске тіндерде жеткілікті молшерде болатын фосфор кышкылы, пентоздык цикл кезінде тузілген пентозалар, амин кышкылдарынан тузілеиін пуриндік жэне пиримидиндік негіздер кажет. Нуклеопротеид тер синтезі коп уакыт алады жэне жогары энергия шыгындарын кажет етеді.
Нуклеопротеидтердің корытылуы ас корыту жолдарында аскорыту сөлдерінің ферменттерінің әсерінен журеді. Белоктық бөліу амин кышкылдарына дейін
ал нуклеин кышкылдары озінің курам бөліктеріне (пуриндік және пиримидиндік негіздер, пентоза, фосфор кышкылы) дейін гидролизденеді.
Пуриндік негіздердің тотыгуы, несеп кышкылынын тузілуі:
Пуриндік және пиримидиндік негіздердің ыдырауы әр турлі жолдармен өтеді. Адам агзасында пуриндік гидролитикалық дезаминдену аркылы несеп кышкылына дейін ыдырап, осы турде зәрмен сыртка шыгарылады. Несеп кышкылынын тузілуі негізінен бауырда журеді.
Несеп кышкылы - пуриндік нуклеотидтер алмасуының соңғы өнімі. Тәулігіне 0,5-1,0 грамм несеп кышкылы тузіледі. Несеп кышкылының туздары (ураттар) - суда нашар еритін косылыстар. Несеп кышкылыныцң тузілуі кушейгенде жэне оның шыгарылуы темендегенде подагра дамуы мумкін . Подагра - бул гиперурикемиямен, артритпен, тіндер мен буындарда несеп кышкылының натрий туздарының
жиналуымен сипатталатын пуриндік алмасудың бузылуы.
Жаңа туган нәрестелердің өмірінің алгашкы аптасында физиологиялык гиперурикемия байкалады. Емшектегі балаларда гиперурикемиялык куй кездеседі- нервті-артриттік диатез, бул жагдайда кетоацидоз
жиі байкалады. Бул несеп кышкылынын диабетогенді әсерімен, көмірсулардың экзогендік жолмен түспеуінен , В1 витаминінің гиповитаминозымен және транскетолазаның активтілігінің томендеуімен байланысты болуы мумкін
84-85 Нейроэндокриндік реттелудің схемасы. Гормондар, түсінік. Химиялық табиғаты бойынша жіктелуі
Гормондар дегеніміз аз молшерде арнайы жасушаларда тузілетн, канга немесе лимфага болініп, нысана- тіндерде зат алмасуды реттейтін, рецепторлар аркылы эсер ететін заттар
Гормондардын химиялык структурасы олардын синтезделетін орнымен аныкталады, оз кезегінде ол кан агысымен тасымалдануын, эсер ету механизмін жэне ыдырау жылдамдыгын аныктайды.
Химиялык табигаты жагынан гормондар 3 топка болшедк
I. Белокты - neптидтi гормондар.
Бул топтагы гормондарды 3 топшага болуге болады;
1. Жай белоктык гормондар: инсулин, паратгормон, осу гормоны, ЛТП
2. Курделі белоктык гормондар: тиреотропты гор мон (ТТГ), фолликула стимулдеуші гормон (ФСГ), лютеиндеуші гормон (ЛГ).
3. Полипептидік гормондар: глюкагон, кальцитонин, окситоцин, вазопрессин, адренокортикотропты гормон (АКТГ).
Олардың ішінде бфазы белсенді емес прогормондар турінде тузілед1 (инсулин, глюкагон).
II. Стероидты гормондар. кортикостероидтар, жыныс гормондары:ерлер және эйелдер гормоны.
III. Амин кышкылдарыныц туындылары болатын гормондар: тироидты гормондар (тироксин-Т4, трийодтиронин-Т3), катехоламиндер (адреналин-А, норадреналин-НА).
Гормондардың бул уш негізгі тобынан баска гормоноидтар бар. Оларга эйкозаноидтар (простаглан- диндер, тромбоксандар, лейкотриендер), ас корыту жолдарының гормондары, т.б. жатады. Олар синтезделген орнында (мысалы, энтерофищп жасушаларда, семіз жасушаларда тузілуші серотонин, гистамин) жергілікті әсер етеді
86. Гормондардың жалпы қасиеттерін атап шығу. Гормондардың рецепторлары, нысана
жасушалары
Құрылысындағы айырмашылыққа қарамастан барлык гормондарда ортак касиеттер болады, муны гормондардың жалпы касиеттері деп атайды.
• Эсер ету арнайылыгының жогары болуы. Гормондардың арнайылыгы гормондардың химиялык курылыстарына, синтезделет орнына, байланысатын рецепторлардын турше, әсер ету механизмше, нысана жасушаларына жэне физиологиялык эффектіне тікелей байланысты. Кез келген гормон букіл метаболизм процесіне эсер етпейді тек белгілі бip заттар алмасуына немесе бip топ реакцияга әсерін тигізеді
• Куаттылыгы. Гормондар канда өте аз молшерде болса да, ( 10'9, 10‘шг/л) оте күшті әсер ете алады.
• Секрециялануы. Гормондар ішкі секреция бездері(эндокринді бездер endo-iinKi, krinen- белемш) деп аталатын ерекше тіндерде тузіледі Бул бездердің ешмдер1 тікелей канга бөлінеді немесе тіндік суйыктыкка, лимфага, одан кейін канга туседі Гормондар узшсаз тузшп отырады, бipaK канга аз молшерде Tycin, кушті биологиялык эффект көрсетіп, ыдырап отырады.
Гормондардың бөлiну ритмі бip-бipiHe сәйкес келмейді Гормондар циркадты ритмде бөлінуі мумкін, мысалы, кортизол, онын кандагы максимальды концентрациясы таңгы сагат 8.00-де, минимальды концентрациясы - тун ортасында байкалады. Гормондар ультрациркадты ритмде бөлінуі мумкін.Гормондар арнайы стимулденуден кейін де (мысалы, бала емізген кезде анада пролактиннің бөлінуі мумкін.
Гормондардың ритмді турде канга түсуі заттар алмасуының биохимиялык реакцияларының координациялылыгын камтамасыз етеді Заттар алмасуын туракты деңгейде сактайды. Коршаган ортаның озгеруі гормондардың синтезделу және канга боліну жылдамдыгының озгеруіне әкеледі осыган байланысты биохимиялык реакциялардың жылдамдыгы мен багыттары озгереді ягни заттар алмасу деңгейінің өзrepyi және организмнің өмip суру ортасына бейімделуі камтамасыз етіледі Гормондардың канга бөлінуінің маусымдык ритмдері белгілі
• Кашыктыктан эсер eтyi (дистанттылыгы). Гормондар тузілген орнынан әр турлі кашыктыкта әсер етеді
• Гормондар канда бос куйінде және белоктар- мен байланыскан куйде болуы мумкін. Бос гормондар биологиялык белсенді ал белоктармен байланысканда белсенділігі уакытша жойылады. Гормондардыц бос жэне байланыскан турлерінің арасында белгілі динамикалык тепе теңдік бар. Бос куйінде гормондар азайганда, белоктармен байланыскан гормондардың Typi диссоциацияланады. Байланыскан гормондар суда жаксы ериді олар гормондардың тасымалдану Typi жэне коры болып саналады.
• Уакытша эсер eTyi. Гормондар кыска уакыт эсер етеді Бул олардың жартылай ыдырау уакытының аз болуына байланысты. Белоктык-пептидті гормондар,
адреналин мен норадреналин 3-10 минут гана өMip суре алады. Буларды стресс гормондары деп атайды. Ал, стероидты жэне тироидты гормондар узагырак эсер етеді - бірнеше сагат, бірнеше тәулік, кейбір жагдайларда апталар бойы өMip суреді Оларды бейімделгіштік /адаптациялык/ гормондары деп атайды.
• Барлык; гормондар оз эсерін рецепторлар аркылы тигізеді Рецепторлармен баска да заттар, мысалы, дәрілік заттар, биогенді аминдер жэне баска да заттар байланысуы мумкін. Рецепторлармен байланысатын заттарды лигандтар деп атайды. Сигналдарды рецепторлар аркылы кабылдау тек кана кеп жасушалы организмдерге тән.
87. Гормондардың әсер ету механизмдері (жалпы түсініктер). Гормондардың әсер етуінің
жасушаішілік екіншілік делдалдары. Протеинкиназалар. Жалпы сипаттама, өкілдері. Олардың активтенуі қалай іске асады
Барлык гормондар жасуша ішілік ферменттердің белсенділігін өзгерту аркылы метаболизмді реттейді
Ферменттер белсендігін өзгерту тәсіліне карай эсер ету механизмі 3 топка болінеді
I. Жасуша ішілік ферменттердің активтілін озтерту жолымен әсер етуші гормондар. Бул гормондар жасуша мембранасының бетінде орналаскан рецепторлармен байланысып, екіншілік мессенджерлер аркылы әсер етеді
а/ мессенджері цАМФ болатын гормондар, олар аденилатциклазаны белсендіру аркылы әсер корсетеді буларга белокты-пептидті гормондар (инсулиннен баска), катехоламиндер жатады;
б/ екіншілік мессенджері цГМФ болатын биологиялык активті заттар: журекшелік натрий-уре- тикальщ фактор, брадикинин, ацетилхолин, гистамин, азот оксиді және журектің ишемиялык ауруларында колданылатын нитропрепараттар, бактериалды эндо- токсиндер;
в/ екіншілік мессенджері кальций немесе фосфа- тидилинозитол болатын гормондар: норадреналин, гонадотропин-рилизинг факгорлары, тиротропин- рилизинг факторлары, вазопрессин, ацетилхолин жэне т.б.
Біріншілік механизм бойынша инсулиннен баска барлык белокты-пептидті гормондар және катехоламиндер эсер етеді Бул гормондар кажет болганда тез тұнып кунт биологиялык эсер көрсеткенде ыдырап кетіп турады.
II. Жасуша ішілік ферменттердің белсенділі белоктар мен ферменттердің синтезделу жылдамдыгын
Өзгерту жолымен эсер етуин гормондар. Бул гормондар жасуша iшілік рецепторлармен: цитозольдік, ядролык немесе органоид рецепторларымен байланысады. Бул топка стероидты жзне тироидты гормондар жатады.
III. Жасуша iшілік ферменттердің белсенділігі плазма мембранасының откізгіштігін өзгерту жолы мен эсер етуші гормондар. Бул гормондар плазма мембранасында орналаскан рецепторлармен байланысып, оз әсерін тирозинкиназа-фосфатаза жуйесі пайдалану аркылы іске асырады. Бул кезде жасуша iшілік ферменттердің белсенділігінің өзгepyi субстрат концентрациясынын артуы, белок-транспортерлер мен иондык каналдардың активтенуі есебінен жузеге асады. Мундай гормондарга инсулин, соматомедин-
дер, пролактин, вазопрессин жатады.
88. Гормондардың ІІ механизм бойынша әсері. Қандай гормондар осы механизм арқылы әсер
етеді?
89. Гормондардың (инсулиннің) ІІІ механизм бойынша әсері (мембраналардың өткізгіштігін
жоғарылату арқылы) Барлык гормондар жасуша iшілік ферменттердің белсенділін озгерту аркылы метаболизмді реттейді
Ферменттер белсенділін езгерту тәсіліне карай әсер ету механизмі 3 топка болінеді
I. Жасушаішілк ферменттердің активтігін озгерту жолымен эсер етуmi гормондар. Бул гормондар жасуша мембранасының бетінде орналаскан рецепторлармен байланысып, екіншілік мессенджерлер аркылы эсер етеді
а/ мессенджер! цАМФ болатын гормондар, олар аденилатциклазаны белсенд1ру аркылы эсер корсетеді буларга белокты-пептиді гормондар (инсулиннен баска), катехоламиндер жатады;
б/ екіншілік мессенджері цГМФ болатын биологиялык активт1 заттар: журекшелік натрий-уретикальщ фактор, брадикинин, ацетилхолин, гистамин, азот оксиді жэне журектің ишемиялык ауруларында колданылатын нитропрепараттар, бактериалды эндотоксиндер;
в/ екіншілік мессенджері кальций немесе фосфатидилинозитол болатын гормондар: норадреналин, гонадотропин-рилизинг факгорлары, тиротропин- рилизинг факторлары, вазопрессин, ацетилхолин жэне т.б.
Біріншілік механизм бойынша инсулиннен баска барлык белокты-пептидтік гормондар жэне катехоламиндер эсер етеді Бул гормондар кажет болганда тез тузіліп, кунт биологиялык эсер көрсеткенде ыдырап кетіп турады.
II. Жасуша ішілік ферменттердің белсендшгш бе локтар мен ферменттердің синтезделу жылдамдыгын
езгерту жолымен эсер етуші гормондар. Бул гормондар жасуша iшілiK рецепторлармен: цитозольдк, ядролык немесе органоид рецепторларымен байланысады. Бул топка стероидты жзне тироидты гормондар жатады.
III. Жасуша iшілiK ферменттердіңбелсендшгш плазма мембранасынның отюзгштпн езгерту жолы мен эсер ететінтгормондар. Бул гормондар плазма мембранасында орналаскан рецепторлармен байла нысып, оз әсерін тирозинкиназа-фосфатаза жуйесш пайдалану аркылы іске асырады. Бул кезде жасуша inrmiK ферменттердщ белсендилігін өзrepyi субстрат концентрациясынын артуы, белок-транспортерлер мен иондык каналдардыц активтенуі есебінен жузеге асады. Мундай гормондарга инсулин, соматомедин-
дер, пролактин, вазопрессин жатады
91. 91. Эпифиздің гормоны (мелатонин), химиялық табиғаты туралы түсінік, нысана- жасушалары, әсер ету механизмі, физиологиялық әсері.
Мелатонин - эпифиздің негізгі гормоны. Эпифиздің белсенділігі жарыктвн тусуше байланысты.
Сонымен бipre, эпифизде химиялык табигаты жагынан пептидтер болатын заттар тузеді, буларды кейб1р галымдар нагыз гормондарга жаткызады.
Мелатонин триптофаннан тузыледы триптофан гидроксилденып (5С орында), 5-окситриптофан тузыледы, ол ары карай декарбоксилденып серотонинге айналады. Серотонин ацилденуы нэтижесынде ацилсеро- тонин тузыледы, ал ацилсеротонин ерекше ферменттын катысуымен метилденып, мелатонин тузшеды Твндеп мелатониннын концентрациясы жэне бул ферменттын белсендыгы эпифиздын функционалдык кушынын корceTKiші болып табылады.
Буган дейш мелатониннын уйкынын жылдам фазасына жауап беретыны тунде тузылетыны(тунп сагат 11-ден тацгы сагат 7-ге дейш тэулпгпк мелатониннщ 70%-ы тузшед1) белгыоы болды. Казыргы кезде мелатонин рилизинг-факторлары тузылетын гипоталамустын орталык томпагынаәж әсер етып мидын гипоталамикалык аймагын баскаратын биологиялык сагат деп саналады. Сонымен, 1) соматотропты гормоннын, инсулин тәрізді осу факторыныц денгейін арттырып, организмнінің ocyiHe эсер ететшып 2) мелатонин гипо
таламус аркылы гонадотропты гормондардын секрециясын тежеп, организмнын жыныстык кызметыне эсер ететыныбелгЫлыболды. Сондай-ак, эпифиздын iciктері кезіндк жыныстык жетылу бузылатыны, закымданган эпифизды алып тастаганда жыныстык дамудын кайта калпына келетыны аныкталды. Мелатониннын антигона - дотропты эсер корсете алатыны жэне урык бездерыне тыкелей эсер етып тестостероннын тузылуын тежейтыны дэлелденген. Мелатониннын адамнын жыныстык функциясын реттеудегіи poлі ер балаларда жыныстык жетылудын алгашында кан сарысуындагы мелатониннщ децгейыные курт темендейт1н1 аныкталган. Эйелдерде мелатониннщ мелшер1 менструация кезынде артады, ал овуляция кезынде азаяды.
3) Мелатонин иммундык жуйенын кызмет: а) оган ткелей стимулдеушы эсер корсету аркылы; б) тимустын инволюциясын тежеу в) Т-лимфоциттердын тузшуые калпына келтыру; г) иммундык жуйеге жанама реттеушы эсер беретынн тиреоидты гормондардыесинтезынын стимулдеуыек жэне мырыштын метаболизмше катысу аркылы реттей/ц;
4) Мелатонин кандагы холестерин денгейын томендетед^ атеросклероз жэне стенокардия дамуын тежейдг Стенокардия кезынде кандагы мелатонинные молшер1 5 есе томендейд1, ал мелатонинные денгейы жогарылаганда стенокардиянын устамалы ауру
сез1мдер1 токталады.
5) Мелатонин кан кысымын томендетеды
6) Мелатонин тез ocin келе жаткан жасушанын
пролиферациясын баяулатады, катерлы iciKTin ocyiH токтатады. Сондыктан оны iciKTin алдын алуга жэне емдеуге колдануга болады.
7) Эндокриндык бездердые кызметын жэне олардын морфологиясын калпына келпру аркылы организмд1 жасартады.
Мелатонин жас адамдарда 50-жастагылармен салыстырганда 4 есе, ал 80-жастагыларга Караганда 8 есе коп тузыледы Мелатонин липидтерддын пероксидті тотыгуын тежей отырып, OMip суру уакытын узартады, картаю процесын тежейд1, карттык катаракта және баска да ауруларын алдын алуга кемектеседы
Организмнын картаюынына жэне кептеген аурулардын пайда болуынын 6ip ce6e6i жасушалар мен молекулалардые липидтердые пероксидты тотыгуы эсерыеке бузылуы болып табылады. Мелатонин оте күшті антиоксидант, ол тотыксызданган глутатион- нан 6 есе кушты. Сол себепт1 ол жасушаларды липидтердынпероксидті тотыгуынан жэне радиация- дан коргайды. Мелатонинные кабылдаганда ДНК-нын радиациялык закымдануы 99%-га темендейд1, ол глутатионпероксидазанын активтылыгые стимулдейды
Мелатонин арнайы рецептор аркылы емес, ол мембрана аркылы (гематоэнцефаликалык тоскауыл аркылы) жасуша ишине тусе алады. Сондыктан мелатонин миды баска заттарга Караганда зиянды эсерлерден жаксы коргайды жэне онын жаска байланысты азаюы ми тындерынын озгергыштыгыне экеп согады. Сонымен, мелатонин мига мынадай себептер ушын кажет:
1. Ми тіндерінде липидтердые пероксидтв тотыгуы каркынды журед1, өйткені ми 02 пайдалануда 6ipiHmi орын алады, сондай-ак онда прооксидант болып табылатын оттектынактивті формаларын тузетын гемс1з тем1р кеп.
2. Ми тындерыне гематоэнцефаликалык барьер неri3ri антиоксиданттардын тусуые шектейтындыктен,мида антиоксидантты жуйенын жетыспеушылыгы бай калады.
3. Мелатонин глутатионпероксидазанын активтылыгын стимулдеу аркылы миды коргайды. Жуйке тынынде бул фермент тотыксызданган глутатионнын тотыгуын тездетед1, осы кезде сутек аскын тотыгы суга айналып зиянды гидроксил радикалдарынын тузшуы токталады, ягни жуйке жасушаларынын де- генерациясын жэне ел1мш тежейдг Супероксидани- онды активаздещцретш супероксиддисмутазанын (СОД) жогарылауынан жас улгайган сайын сутек пе- роксидшщ жиналуы, ал баска Heri3ri ферменттердщ - каталаза мен глутатионпероксидазанын молшершщ жогарыламауы б1здщ антиоксидантты корганыштык жуйем1здщ непзп KeMicTiriHe жатады. Антиокси дантты ферменттердеп бул дисбаланс жаска байла нысты мелатониннщ тузшушщ томендеу1мен 6ipre болады.
Сонымен, мелатонин б1здщ денсаулыгымызды, энергетикалык куцпм1зд1 туракты устап туратын, уйкы, тэбет, журектщ жиырылу ж и т г ш , дене темпе- ратурасын, козгалыстыц координациясын, куану мен жабыркауды, репродукцияны, жыныстык белсендшюл бакылайтын циркадты ыргактыц стимуляторы болып табылады.
Мелатонин рилизинг-факторлардыц тузшуше эсер етш гипоталамус жумысын реттейд1, ал олар аркылы гипофиз жэне iшKi секреция бездерынын жумысынбакылайды. Бул курдел1 жуйенын калыпты жумыс icTeyi ритмдердын б1рмезгшд1 (синхронды) болуымен камтамасыз етыледв
Осыган орай, мелатонин нейромедиатор сиякты нейро-эндокриндж жуйены баскарады, сондай-ак ГАМК-мен 6ipre нейро-эндокринд1 жуйеные ic-эрекетын синхрондау ушин ми кыртысындагы нейрондардын козуын тежеу1де мумкын.
Мелатонинные тапшылыгы ми жасушаларынын шамадан тыс жане санага багынбайтын козуына экелу1 мумкын Мелатонинды емдыкмаксатка колдануга бо лады, ce6e6i узак уакыт мелатониннын артык дозасын пайдалану зиянды эсер корсетпейды
1. уйыктататын зат ретынде уйыктау уакытын тездетед1 жэне уйкыннвнуакытын узартады.
2. уакыттык белдеу1 мен жыл мезгылдерынын озге- pyiHe бейымделуд1 (адаптацияны) женылдету ушын;
3. стресс эсерые темендетеды
4. Паркинсон ауруын, желыкпе-депрессия синдром- дарын емдеу ушые колданады;
5.Ышыктерды (оспе) емдеу ушын.
92. Гипофиздің алдыңғы бөлімінің гормондары, жалпы сипаттама, өкілдері, химиялық табиғаты
туралы түсінік, нысана-жасушалары, әсер ету механизмі, физиологиялық әсері.
Гипофиздың алдынғы бөлімінің гормондары. Химиялык табиғаты, нысана жасушалары, әсер ету механизмі физиологиялык әсерi, секрециясының реттелуы
Соматотропты гормон немесе өсу гормоны (2-Ш1 сабакты караңыз).
Адренокортикотропты гормон (АКТГ)
Химиялык табигаты: Бул 39 аминкышкылынан туратын полипептид. Гормональдык активтілікті 24 аминкышкылынан туратын шеткі бөлігы камтамасыз етеды
Әсер ету механизмі: бірыншілык ц.АМФ аркылы.
Нысана жасушалары: 1) бүйрекүсті безінің қыртыс қабатының шоғырлы аймагы; 2) буйрек усті безынің кыртыс кабатының шумақты аймағы; 3) май Tiні
Физиологиялык әcepi:
1) Глюкокортикоидтардн (ГКС) жэне аз мол шерде минералокортикоид-тардын(МКС) секреция сын стимулдейді 2) глюкокортикоидтардьн және минералокортикоидтардың физиологиялық әсеріне ие; 3) май тінінде липолизды жылдамдатады; 4) терының пигментациясын кушейтеді, ceбeбi меланинді синтезын стимулдейды
Секрециясынын бузылуы - гормонды гипер- продукциясы Иценко-Кушинг синдромы турінде болады. Гинопродукциясы Аддисон ауруы турынде байкалады.
Тиротропты гормон (ТТГ)
Химиялык табигаты: бул 2 суб бірліктен туратын нагыз гликопротеид - оның а-суббірлігі 96 аминкышкылынан (ФСГ мен ЛГ-нщ а-суб бірлігіне ұксас) жэне гормонлы иммунологиялык және биологиялык белсендилігін аныктайтын, кұрамында 112 аминкышкылы бар р-суббірліктктен турады.
Әсер ету механизмаі біріншылік ц.АМФ аркылы.
Нысана-жасушалары: калканша без! жэне май TiHi.
Физиологиялык әсері:
> тироидты гормондардын секрециялануының барлык сатыларына катысады: а) йодидтік насостын кызметын стимулдейді, бул йодтын калканша безының жасушаларына тусуының жогарылауына әкеледі; б) тирозиннын иодтануын камтамасыз етеді; в) тиро- глобулиннын синтезыне әсерін тигызеды г) Т3 пен Т4-тын тузылуыне катысады; д) гормондардының тироглобулинның курамынан босап шыгуын камтамасыз етеді е) олардың канга болынуыне әсерые тигызеды
> нысана-жасушаларында липолиз бен глико- генолизді кушейтеді
> калканша без васкуляризациясын стимулдейді. калканша без фолликулярлык жасушаларының саны мен көлміын арттырады. ТТГ көп мелшері тунде секрецияланады, танга жуык төмендейді
Секрециясының бузылуы - гиперпродукциясы кезынде гипертиреоидизм белгылері болады; гипо- продукциясы кезынде гипотиреоидизм белгілеры байкалады.
Гонадотронты гормондар - фолликуло стимулдеуді гормон (ФСГ) және лютеиндеупи гормон (ЛГ), пролактин (лактотропин - ЛТГ).
ФСГ мен ЛГ 6ipiншілік механизм бойынша цАМФ аркылы әсер етеді ФСГ - нагыз гликопротеид, 2 суббірліктен турады - а (96 аминкышкылының калдыгынан) жэне В (3- (112 аминкышкылының калдыгынан).
Биологиялык арнайылығын (3-суббірлік анытайды.
Нысана жасушалары: жыныс бездеры Физиологиялык әcepi:
ФСГ - әйел адам организмінде аналык бездерді
ӨcyiHe жэне фолликулалардың жетілуіне ыкпал етеді андрогендердын эстрогендерге айналуын камтамасыз етеды Ер адам организмінде сперматогенезді активтендіреді. Сертолли жасушаларымен андроген-байланыстырушы протеинді тузылуыне стимулдейді .
Эюәйелдерде ЛГ аналык жасушалардың жетуі және овуляцияны; прогестеронды синтезын аналык бездерде эстрогеногенез бен андрогеногенезді камтамасыз етеды . Ер адам организмде Лейдиг жасушаларында тестостеронның тузілуын стимулдейді ЛГ сперматогенезді кушейтеді ЛГ-ның тунгы секрециясының жогарылауы - жыныстык жетылуде бырыншы белгысы
Пролактин, немесе лактотроидты гормон -
ГТГ-га жатады. Пролактин - курамында 199 амин- кышкылыны калдыгы бар жай белок.
Әсер ету механизімі: бірыншылык және ушыншылык.
Физиологиялык әcepi: әйел адам организмынде гормон сут бездерыне эсер етеді оларды ©суше жэне суттын тузілуіне ыкпал етеды Пролактин сары денелі жасушаларына әсер етын прогестеронның тузылуін камтамасыз етеді (осы себептен пролактин баска аталуы - лютеотропты гормон); пролактин жыныс бездерде ЛГ- рецепторларының синтезін арттырады.
Ер адамдарда гормон май бездерынын кызметын стимулдейды Пролактин Лейдиг жасушаларына әсер етып , олардагы тестостероннын тузылуын камтамасыз етеды Пролактин буйректе эритропоэтинн тузылyiH стимулдеп эритропоэздж әсер корсетедг Гор мон АКТГ-ныц секрециясын стимулдеп, әйелдердын буйрек ycTi бездерінде андрогендердiң синтезын жогарылауына әкелуы мумкын
Пролактиннің секрециясын гипоталамустан бөліHin шыгатын дофамин тежейді Сонымен 6ipre про- лактолиберин мен пролактостатин да пролактиннің секрециясына әсер етеді
93. Соматотропты гормон, соматомедин С, түзілетін орны, химиялық табиғаты туралы түсінік,
нысана-жасушалары, әсер ету механизмі, организмге әсері, секрециясының реттелуі. СТГ
гипо- және гиперфункциясы.
Химиялык табигаты: 191 амин кышкылының калдыгынан туратын, арнайылығы бар жай белок.
Әсер ету механизмі. Біріншілік және ушіншілік механизм аркылы әсер етед( өзі әсер тиро-
зинкиназды-фосфатазды каскад) ; фосфатидилинозитдж жүйе және соматомединдер (метаболикалык эффект) аркылы өсу гармондарынын әсіресе бауырда соматомединдер тузіледі олар 3 турлі болады - A.B.C. А,С соматомединдер шемірмек, суйек, дәнекер індерше инсулин тәрізді, В соматомединдер-ОЖЖ-га әсер етеді Инсулин тәрізді эффектінің соматомединдердщ нысана жасушаларынын етізпштнт жогарылап, аденил атциклазанын активтілікгі темендеуімен Г А жэне С соматомединдері дэнекер тіндер жасушаларынын тутасып өсу, сүйек жәнене шем1ршек тіндерінде ДНК мен РНК синтезі артады, В-соматомединдер тек жуйке тіндерде эсер етедь
СТГ-нын нысана-жасушалары: агзанын барлык жасушалары.
Физиологиялык acepi: Гормондардың әcepi былай реттеледі
нысана-жасушаларының пролиферациясын стимульдейді СТГ бауырда соматомединдер манальгетиктерде инсулин (инсулин тэр1зд1 эффект) стимульдейдү Осы ксзде АК, уридин, тимидин жэне сульфаттардың саны шеміршек жасушаларына Tycyi артып, белок, нуклеин кышкылдарының синтезі және хондроциттерді және баска да жасушаларының пролиферациясы артады.
- метаболикалык эффект: СТГ комірсулар алмасуына инсулин тәрізді әсер етеді, 6ipaK кыска уакыт
(30-40 мин). Глюкозаның жасушаларга тусуі және тотыгуын арттырып, одан кейн көмірсуларга кон- тринеулярлы әсер көрсетеді глюкозаның май тідері мен булшык еттерде пайдаланылуы томендейді, бауырда глюконеогенез артады. Осы сиякты липидтер алмасуына әсерш типзеді 30-40 минут ішнде инсулин тәргзді әсер ет., липогенезді кушейтеді, одан кейн май тіндерінде липидтерді синтезі төмендейді, бауырда жэне май тідерінде глюкозаның жумсалуы
томендиді, канда БМК молшері артады, липолизде жогарлауынан кетогенезді тугызады.
СТГ уйкы безге әсер ет., глюкагон тузуйне әсер етедг
фосфатидилинозитолды жуйе аркылы әсер eTin, кальцийді кандагы жэне жасуша сыртындагы ортага болу томендегедй
СТГ булшык еттерге инсулин тәрізді эсер етеді (кыска мерз1мді); 6ipaK бул гормон узак уакыт көп мөлшерде әсер етсе, инсулинн салыстырмалы жетіспеушілігі тугызады, ягни диабет ауруын коздыруы мумкін. СТГ бауырдагы глюкоза ешмдері арттырып, тіндерде глюкозаның мөлшерш төмендетеді Сол себепті СТГ-Hi контринсулярлы гормондарга жаткызуга болады. СТГ-нің багыты (инсулин тэр1зд1 жэне контринсулярлы) эсерлері есу мен анаболизмді жогарылатын, катаболизмд1 азайтады.
СТГ тузшуi мынандай жагдайларда артады:
- уйкы;
жаракат алган кезде жзне пирогендер әсер.;
булшык еттерге жасалатын жаттыгулардан ;
- гипогликемияда;
вазопрессин енгізгеннен кейн; наркотикалык анальгетиктерд1 енізген соң.
СТГ-ныц секрециясы соматолиберин, тиролибе- рин, (3-эндорфин, дофамин, серотонин, норадреналин, ацетилхолинмен стимульденедг Соматостатин, адре налин жэне кан плазмасындагы БМК эсершен секрс- циясы тежеледг Тэушгше гормон 4-10 рет импулыгп турде тузшедг Глюкагон жэне эстроген СТГ-ныц бо- сап шыгуын жогарылатса, прогестерон СТГ түзшуш томендетеді
СТГ секрециясының бузылуы: Гормонныц артык OHiMi байкалады: 1) гипофиздің алд. Болигинде ickrep пайда болган кезде; 2) кейб1р жукт1 әйелдерде босану алдында байкалады.
Ересек адамдарда гормонның артык мөлшерде тузшушен «акромегалия дамиды.
СТГ балалык кезде коп молшерде синтезделсе ол «гигантизмге» әкеледі Гормонныц жетюпеуі ергежейлжке әкелед1 (6ipaK гигютиреозбен салыс- тырганда бул ергежейлікте (карлик) адамныц акыл- eci дурыс болады, СТГ жеткеуі балаларда
тек бойы өспейді- «акылды ергежейлж»).
.
95. Гипофиздің гонадотропты гормондары (ФСГ, ЛГ), химиялық табиғаты туралы түсінік,
нысана-жасушалары, әсер ету механизмі, физиологиялық әсері. Секрециясының реттелуі
пролактин (лактотропин - ЛТГ). ФСГ мен ЛГ 6ipiншілік механизм бойынша цАМФ арқылы әсер етеді ФСГ – нағыз гликопротеид, 2 суббірліктен тұрады (96 аминқышкылының қалдығынан) және (3- (112 аминқышкылының қалдығынан).
Биологиялық арнайылығын (3-суббірлігін анықтайды.
Нысана жасушалары: жыныс бездері
Физиологиялык әcepi:
ФСГ - әйел адам организмінде аналық бездерге
eнyiнe және фолликулалардың жетуіне ықпал етеді андрогендердің эстрогендерге айналуын қамтамасыз етеді. Ер адам организмінде сперматогенезді активтендіреді, Сертолли жасушаларымен андроген-байланыстырушы протеиннің түзілуін стимулдейді .
Әйелдерде ЛГ аналық жасушалардың жетілуіне және овуляцияны; прогестеронның синтезі; аналық бездерде эстрогеногенез бен андрогеногенезді қамтамасыз етеді. Ер адам организмінде Лейдиг жасушаларында тестостеронньң түзілуін стимулдейді . ЛГ сперматогенезді күшейтеді ЛГ-ның секрециясының жоғарылауы - жыныстык жетілудің 6iріншілік белгісі
Пролактин, немесе лактотропты гормон -
ГТГ-ға жатады. Пролактин - кұрамында 199 амин- қышқылының қалдығы бар жай белок.
Әсер ету механизмі: біріншілік және үшіншілік.
Физиологиялық әcepi: әйел адам организмінде гормон сүт бездеріде әсер етеді олардың жасушалары және сүттің түзіуіне ықпал етеді. Пролактин сары дененің жасушаларына әсер етеді, прогестеронның түзілуіне қамтамасыз етеді (осы себептен пролактиннің басқа аталуы - лютеотропты гормон); пролактин жыныс бездерінде ЛГ-ныц рецепторларының синтезі арттырады.
Ер адамдарда гормон май бездерінің кызметі стимулдейді Пролактин Лейдиг жасушаларына әсер етеді, олардағы тестостеронның түзілуін қамтамасыз етеді. Пролактин бүйректен эритропоэтиннің түзілуін стимулдеп эритропоэзді әсер көрсетеді .Гормон АКТГ-ның секрециясын стимулдеп, әйелдердің бүйрек үcтi бездерінде андрогендердiң синтезінің жоғарылауына әкелуі мумкін.
Пролактиннің секрециясын гипоталамустан бөлініп шығатын допамин тежейді .Сонымен 6ipгe пролактолиберин мен пролактостатин да пролактиннің секрециясына әсер етеді
96. Гипофиздің артқы бөлімінің гормондары, өкілдері, химиялық табиғаты туралы түсінік,
нысана-жасушалары, әсер ету механизмі, организмге әсері. Қантсыз диабет.
VІ рецепторлармен байланысып, 6ipiншілік механизм бойынша (фосфатиди- линозитидті жуйе аркылы) әсер етед1, ал V,-рецепторлар мен комплекс тузуші, 6ipiншілік механизм бойынша (цАМФ аркылы) әсер корсетеді V - рецепторлар фосфолипаза С-мен байланыскан жэне шеті қан тамырларында орналасады. Вазопрессин осындай рецепторлармен байланысып вазокон-стрикцияны тугызады, бул әcepi гормонның атында корсетілген. У7-рецепторлар аденилатциклазамен байланыскан және буйрек тупшелернің дисталды болімінің эпителиінде орналасады. Вазопрессин осындай рецепторлармен байланысып цАМФ аркылы аквапориндерді – судың белок-транспортерлері активтейді сондьқтан, нефронньң дистальды болімшесінде судьң реабсорбциясы кушейеді, ягни антидиуретикалық әсер керсетеді, гормонның екінші аталуы - АДГ (антидиуретикалык гормон).
Вазопрессин – гепатотропты гормон, біркатар маңызды процестердің активаторы. Аштықта ол бауырдагы глюконеогенезд! стимулдейді, ал карын тоқ болган жагдайда - гликогенолиздгегенде гликогеннің тузушісі тежеп, оның ыдырауын жылдамдатады. Вазопрессиннің әсерінен буйрек ycті безінің гормон дарына тэуелді гипергликемия кушейеді. Вазопрессин БМК-ның бауырга тycyiн жэне тотыгуы мен этерификациясын кушейтеді. Ол глюкагонның кетоген әсеріне кедергі болады. Ол глюкагонның кетогенді әсерінің алдын алады. Вазопрессин инсулиннің секрециясын стимулдейді жэне мәйлінше липогенезді, глюкозаның тycyiн және плазмадагы артык БМК-ның адипоцитке жиналуын стимулдеп, оган инсулин тәрізді әсер етеді. Сондыктан оның әсерінен қан плазмасындагы БМК деңгейі узак уакытта мен молшерде болады.
Вазопресиннің эсерінен АКТГ-ның секрециясы кушейеді жэне ол стресс кезінде ЛГ-ның секрециясының тежелуіне катысады. Вазопрессин VIII фактордың синтезі стимулдейді жэне Вилле- бранд факторының секрециясын жогарылатады, ягни кан уюына катысады.
Вазопрессин ОЖЖ-не әсер етеді - урей сезімін тудырады және ауырсыну сезіміне реакцияны томендетеді, еске сактауды стимулдейді және шолдеу сезімін реттелуінің орталык механизмдері ка тысады. Сондыктан вазопрессиннің ролі тек гана осмореттелумен гана шектелмей, тотенше жагдайлар дын орын алуына организмінің карсы туруын стимулдеу болуы да мумкін.
Вазопрессиннің тузілуі бузылганда кантсыз диабет дамиды. Онымен балалар мен жасоспірімдер де ауы- рады. Бул патология барлык эндокринді аурулардын 0,5%-да кездеседі. Бул кезде диурез (10 л/түзуліне дейн) жогарылайды, гипоизостенурия, никтурия байкалады.
Окситоцин цАМФ аркылы біріншілік механизм бойынша эсер етеді. Бipaқ рецептор стимул денгенде нысана-жасушаларында цАМФ деңгейі төмендейді. Рецепторлар жатыр мен сут бездерінің тeric булшык eттepiнің жасушаларында болады. Окситоцин жатырдың жиырылуын жогарылатады, альвеолалардан суттің бөлінуін камтамасыз етеді. Оны дәрілік препарат босануга жагдай жасау ушін колданылады.
Буйректе окситоцин элаз ангидиуретикалык әсер корсетеді жзне кыртыстың нефрондарында кан айналысы мен сузуді стимулдейді. Май тінінің метаболизмге вазопрессин сиякты әсер етеді, ол вазопрессинмен 6ipre организмді стресс кезінде кетоз дан коргауы мумкін. Окситоцин организмнің ауру сезімге жауабын темендетеді оның босану кезінде физиологиялык маңызы бар. Бул гормон жасушалык иммунитет стимулдеуі мумкін деген мәліметтер бар.
Окситоцин есте сактауды стимулдеп, орталык эффектерді көрсетеді. Ер адамдарда окситоциннің әсерінен жыныс органдарды стимулдеген кезде жыныс гормондарының секрециясы кушейеді жэне эвокуляция кезінде жыныс гормондарыньң концентрациясы жогарылайды.
.
97. Қандағы кальций мен фосфаттардың деңгейін реттейтін гормондар (паратгормон, кальцитонин, кальцитриол), түзілетін орны, химиялық табиғаты туралытүсінік, нысана- жасушалары, әсер ету механизмдері, физиологиялық әсері, секрециясының реттелуі
Кальцитонин - қалқанша безінің парафолликулярлы жасушаларының гормоны.Қалқанша безі кабырғалары куб тәрізді эпителиймен тысталғн фолликулалардан тұрады.Қалқанша безі фолликулаларының коллоидында гормон трииодтиронин мен тетраиодтиронин , ал парафолликулярлы жасушаларынжа тиреокалдитонин ( кальцитонин) гормонын түзеді .
Химиялық табиғаты: кальцитонин 32 амин қышқылының қалдығынан тұратын полипептмд .
Әсер ету механизмі : басқа да белокты пептидтік гормондар сиякты аденилатцилазаны белсендіру арқылы әсер етеді .
Нысана жасушалары : сүйек тін, бүйрек , ішек қабырғасы .
Физиологиялық әсері : жануар организмінде кальциймен фосфор алмасуын реттейді
Сүйек тініне әсері: кальцитонин әсері остеокластардың белсенділігі мен диффреенцировкасы тежеледі .Деминерализацияның тежелу ғана еиес кальцийдін қаңқада фиксациялануы күшейеді .Сонымен гормонның әсері негізінен остеокластардың әсеріе тежеуге , остеобластарды активтеуге бағытталған , сондықтан қандағ кальций мөлшері төмендейді .
Бүйрекке әсері: бүйректен зәрдін шығарылуына кальцийдйң реабсорциясын азайту арқылы оның организмнен шығарылуына әсер етеді .
Натрий зәрмен шығарылады , осы жағдайда диурездің артуы байқалады .
Сүйек қабырғасына әсері: Кальцийдің және фосфаттардың тежелуіне бағытталған .Кальцитонин асқазанда жәе он екі елі ішек түзетін ас қорыту жолдарының гормоны гастрин түзуін тежейтіні анықталған. Бұдан басқа кальитонин трипсин амилазаның , тұз қышқылының түзуін бәсендетіп ішекте кальцийдің, натрийдың хлордың және судың секрециялануын арттырады. Гастрин гипрекальцемия жағдайындағы сияқты кальцитонин түзілуін стимулдейді. Бұдан кальцитонин астың қорытылуы мен сіңірілу қәзірге белпаз роль атқаруы мүмкін деп болжас жасауға болады. Кальцитонин гипоталамус және лимбиялық жүйе деңгейшеде айқын тура әсер көрсетеді. Гормонның секреция қандағы кальциі мөлшерімен реттеледі гиперкальцемия секрециясын арттырса ал гипокальциемия оны тежейді мұндай реттелу глюкагонға да тән құрамыеда кальций мөлшері көп диета ұстағвнда глюкагон серециясы артады .Глбкагон кальцитонин секрециясының активаторы болып табылады .Д3 витаминінің мөлшері қалыпты болғанда паратгармон мен кальцитомин ара - қатынасы қандағы кальций мөлшеріне байланысты болады
Химиялық табиғаты : паратгормон 84 амин қышқылынан тұратын жай белок .Паратгормое молекуласындагы полипептидтіқ тізбектің N шетінен басталатын 34 амин қышқылынан тұратын биологиялық белсендік көрсетеді қалған 50 амие қышқылы молекулаға тұрақтылық береді .Осы белгі арқылы гормон рецептормен байланысады.Қалқанша маңы бездері жасушалары паратгормонға ұқсас пептид өндіреді .Бұл пептид кейбір ісіктін , сүт бездерінің , өкпенің , мидың , қан жасушалары тамырларының плацентаның жане миокардтың жасушаларында түзеді .
Әсер ету механизмі: сүйек тіні, бүйрек , ащы ішектің кілегей қабаты .
Физиологиялық әсері
Сүйек тініне әсері: сүйек тінінде 3 түрлі жасушалар остеобластар, остеоциттер, остеогластар бар .Паратгормонның әсерінен остеокластар мен остеоцидтер активтеніп стимулденеді. Остеокластардың саны артады, остеобластардың активтігі уақытша тежеледі. Остеоциттер мен остеокластарда қышқылдық фосфатазаның коллагеназаның фосфорилазаның активтеп артады .Белоктар соның ішінде коллаген жеке амин қышқылдарына дейіе ыдырайды, бұл кезде қанда оксипролин мөлшері артады , ол бүйрек арқылы зәрмен шығарылады . Зарден окипролин мөлшеріне қарап паратгормонның әсер ету дәрежесін біліге болады .Коллаген мөлшері төмендеуі онымен байланысқан гидроксиатит молекулаларының босап шығуына әкеледі .Паратгормонның әсерінен гликолиз күшейеді соның нәтижесінде сүйекте лактат жиналады. Цитраттың түзу сатысында үш карбон қышқылдары тежеледі гормон изоцитратдегидрогеназаның ингибирлейді бұл цитраттың жиналуына әкеледі жасуша pH қышқылдық жаққа ауысады , минералды заттардың сүйектен шығуы жылдамдайды .Бұдан басқа гидроксиапатитке қарағанда жақсы еритіе лактат пен цитраттан кальций тұздарын түзеді ал бұл кальцийдйң сүйектен шығуына ықпал етеді .сонымен паратгормон әсернен сүйек тіні органикалық негіз ыдырауы ( резорбция ) күшейеді содан кейіе деминерализация да байқалады .Минералды заттар ( гидооксиапатиттер ) Ca2+ және Po43_ ка ыдырайды да олар қаңға түсіа гиперкальцемияны тудырады. Қанда фосфаттың мөлшері артпайды себебі ол зәрмен шығып кетеді .
Бүйрекке әсері : паратгормонның әсерінее бүйрек каналда зәрден қанға кальциймен магнийдйң реабсорбциясы ( кері сіңірілу ) артады .Фосфаттардың калий мен бикарбонаттардың зәрмен шығарылуы артады , ал аммоний протондардың магнийдің эксрекциясы төмендейді.
Ішектің кілегеі қабатына әсері :
Паратгормое Ca 2+ шектен қанға сіңірілу күшейеді Паратгормонның әсерінен бүйректен кальцитриолдың түзуіе тездететін фермен гидроксилаза активтеп артады .Кальцитриол қан ағысымен ішек қабырғасына түсіп гормон сиякты әсер етіп са тасымалдаушы белоктың түзуіне әкеледі .Осы белоктың әсерінен кальцийдін жәе онымен байланысқан фосфордың ішек қабырғасына сіңірілуі артады .Сонымен паратгормонның әсер қандағыикальций мөлшерін арттыруға бағытталған .Паратгормон он инотропты реттецип болып табылады , ол жүректің жиырылуын стимулдейді артериялық қысымлы және бүйректің фильтрациясые жоғарылатады , көмірсулар алмасуына контринсулярлы әсер көрсетеді оның сперматогоненздің бәсендетуі гиперлипопротеинемияны индукциялау тері қышуын болдыртатыны туралы мәліметтер бар
98. Тиреоидты гормондар: трийодтиронин және тетрайодтиронин, олардың түзілуі. Химиялық
табиғаты, нысана-жасушалары, әсер ету механизмі, физиологиялық әсері, секрециясының
реттелуі. Қалқанша безінің гиперфункциясы мен гипофункциясы (ересектер мен
балалардағы) туралы түсінік. Эндемиялық зоб, пайда болу себептері.
Т3 Т4 қалқанша безі фолликулярлы жасушаларының гормондары .
Түзілуі және химиялық табиғаты : тироидты гормондаи тирозиннен түзеді ион түрінде түскен йод сутек асқые тотығының әсерінен қалқанша безде молекулалық йодқа айналады .содан кейін молекулалық йод тирозин йодтауға жұмсалады .Тирозин коллоидта бос күйінде немесе 10 полипептидтік тізбектен тұратыны тироглобулин белогының құрамыеда кездеседі. Тирозин йодтануы 3 немесе 5 орындарда жүреді .Бұл кезде МИТ мен ДИТ түзіледі. Йодталған қосылыстардың екеуі де тироидты гормондардың алғы заты болып табылады , 2 ДИТ конденсацияланғанда Т 4 МИТ және ДИТ конденсацияланғанда Т 3 түзеді .Осы реакциялардың нәтижесінде аланин 35% ы Т3 пен , шамамен 5% ы Т мен 50 % ға жуығы МИТ жәе ДИТ пен байланысатыны анықталған .Кейбір қосылыстар йодтың тотығуын тежеп , Т мен Т 4 түзіне кедергі жасайды .Мұндай заттарғы мочевина, тиоурацил , пропилтиоурацил , метимазол жатады .Бұл қосылыстар гипертиреоидизмді емдеуде антироидты препараттар ретінде қолданылады. Түзген гормондвр тироглобулиннйң құрамыеда болады да жүйке импулсына немесе ТТГ әсернен босап шығады .ТТГ әсерінен тироглобулиннің гидролиз активтеп артады қанға олармен бірге қолданылмаған МИТ пен ДИТ тек қанға дйодиназаның әсерінен йодсызданады .түзілген йод қайтадан пайдаланылады. Тиреоглобулиннің гидролиз иодтың әсерінен тежеледі сондықтан калий йодтын гипертиреоидизмді емдеуге қолданылады .Т 3 екі түрде болады : 3,5,3 - Т3 актива гормон , ал сутектері 3,5,57 орындарда йодидпен алмасқан Т активаз гормонның бұл түрі кері Т ( кері Т3 ) деп белгіленеді. Т 4 80% бауырда дейодтану арқылы Т 3 айналады 2Т4-2Т3+J2
T3 және Т4 қанға тиронин байланыстырушы белокпен және альбуминдермен байланысады. біріншіден Т4 белокпен байланысы Т3 ке қарағандв берік болғандықтан Т4. 9 есе көп түзсе Т 3 актив болады екіншіден Т3 бос түрі(0,5%) бос түрден Т
Бос турден Т4 ке (0,05% аз) караганда кем гиронин байланыстырушы глобулиннен баска тироидты гормондар плазманың липопротеидтермен байланыса алады Т4 ке карагандв белоктарымен байланысады .
Әсер ету механизмі:
Т2 механизм арқылы әсер етеді ал Т4 түгелдей дерлік Т ке айналады, сондықтан Т4 ке әсер ету механтзмі туралы айту ақылға сыймайды .Т3 ядролық рецептормен байланысады Т рецептор комплекс ДНК га әсер ету белоктар мен ферменттердің синтезін стимулдейді .Нысана жасушалары : организмнің көптеген мүшелері мен тіндерінде ( көкбауыр мен ұрық бездерінен басқа ) тироидты гормондардың рецепторлары бар .Әсіресе жүректе бауырда жәе гипофизде рецепторлары көп .
Биохимиялық әсерлері Т3н әсерінен оксидоредуктазалардың синтез күшейеді сондықтан барлық тотығу - тотықсыздану реакциялары жылдамдайды, тіндерде оттегісіне қажеттілігі артады, аэробты тотығу АТФ түзілу ( калоригенді эффект ) артады митохондриялардың саны көбейеді .АТФ фазалардың активтігі және Na + мен K + жасушалық мембраналар арқылы тасымалдаушыларын арттырады. Осы гормондардың әсерінін арқасында организм калория алады АТФ кеп түзеді дене температурасын көтеріп жылуға төзімдік төмендейді .
Көмірсулар алмасуына әсері: ас қорыту жолдарында глюкозаның сіңірілуі артадф , жасушалардың ( бұлшық ет , май тіні) глюкозаны пайдалануы күшейеді , гликолиз , глюконоегенез , гликогенолиз үрдістері күшейеді .
Липидтер алмасуына әсері : липолиз артады , бірақ қандағы БМКконцентрациясы жоғарыламайды себебі оның жасушадағы тотығуы күшейеді .Тироидты гормондар липидтер синтезинде күшеитеді , бірақ бұл гормондардың деңгей артқанда липидтердің ыдырауы басымырақ болып, оның қоры азаяды , адам азады және қандағы ТАГ,ФЛ,ФС концентрациясы төмендейді .
Белоктар алмасуына әсері: гормондардың белоктар алмасуына әсері организмнің өсуі мен дамуы үшін өте маңызды .
.
99. Глюкокортикостероидтар, өкілдері, кортизолдың химиялық табиғаты, түзілетін орны,
нысана-жасушалары, әсер ету механизмі, заттар алмасуына әсері, секрециясының реттелуі.
Бүйрекүсті бездерінің қыртыс қабатының гипо- және гиперфункциясы туралы түсінік.
Глюкокортикостероидтардың қабынуға қарсы, антиаллергиялық және иммуносупрессивті
әсері
99 Глюкокортикостероидтар - буйрек үсті безі қыртыс қабатының шоғырлы аймағында синтезделеді. қaзipгi кезде бүйрек ycтi безі қыртысты қабатынан кортикостероидтар деп аталатын 100-ге жуык холестериннің туындылары табылған. Бұлардың ішінде 10 гормон биологиялық активтілікке ие, ал қалғандары кортикостероидтар алмасуыньң аралык өнімдері болып табылады.
Химиялық табиғаты: Олар холестериннің туындылары. Кортикостероидтардың ортақ ерекшелігі , циклопентанпергидрофосфантреннің сакинасы бар, С4 және С5 арасында қос байланыс бар, С3-те кето тобы, С10 және С13-метил тобы бар. Кортизол мен кортизонда С 17-де (ОН) тобы болғандықтан оларды оксикортикостероидтар-17-(ОКС~17) деп атайды. Кортизол мен кортикостероида С п-де (ОН) тобы бар ,сондықтан олар 11-ОКС деп аталады. Кортизонда С п-де кето-тобы бар. Сонымен, барлық кортикостероидтардың Сп-де әртурлі формадағы оттек болады, ол олардың биологиялық, зат алмасуына тигізетін әсерін анықтайды.Әсер ету механизмі: барлық кортикостероидтар екіншілік механизм арқылы әсер етеді, олардың рецепторлары жасуша ішінде орналасқан. Гормон-рецепторлы комплекс ДНК-ның арнайы белгісімен байланысып, арнайы гендердің транскрипция жылдамдығын арттырып, нәтижесінде белок және фермент синтезі де артады. Сол себепті кортикостероидтардың әcepi 12-24 сағаттан кейін, ал максималды 10-12 күннен кейін әсер етеді (бейімділік гормондары).
Зат алмасуына және түзілетін орнына қарай кортикостероидтар 3 топқа бөлінеді. 6ipiншi қабатта (шумақты аймақта) минералокортикостероидтар (МКС), екіншi қабатта (шоғырлы аймақ)-глюкокортикостероидтар (ГКС), үшінші қабатта (торлы аймақта) андрокортикостероидтар (АКС). Әр топтың гормондары арнайы әсер көрсетеді, басқа топтың гормондарына да әсер етеді, осылай бөлінуі шартты түрде болады.
Бұл олардың құрылыстарының ұксастықтарынан болады. ЛКС ерлердің жыныс гормоны-тестостеронга ұқсас.
ГКС-ныц ішіндегі ең белсенді болып табылатыны кортизол (гидрокортизон) - 70%, кортикостерон (10%), төменгі активтілік көрсететіні кортизон (20%). Адамдарда, маймылдарда жзне иттерде көп мөлшерде кортизол синтезделеді, ал егеукұйрықтар мен қояндарда кортикостерон синтезделеді. Қан плазмасында кортизолдың көп мөлшері арнайы белок- глобулиндер типті транскортин және альбуминдермен байланысады.ГКС-ның плазма белоктарымен байланысуы олардыц eрігіштігін арттырады, органдар мен тіндерге тасымалдануын жеңілдетіп, активтілігі уақытша жойылады, бұзылудан сақтайды.
Нысана-жасушалары: Tepi, көк бауыр, тимустың жасушалары, бауыр, май, лимфоидты, дәнекер және бұлшық ет тіндері
Физиологиялык әcepi: Жалпы әсері катаболикалық, бауырда-анаболикалык реакциялар.
Белоктар алмасуына тигізетін әcepi: ГКС әсерінен бауырдан басқа барлық; нысана-жасушаларында белоктардың ыдырауы жоғарылап, қанда АҚ саны көбейеді. Бауырға түскен 6ipaз АҚ оның трансаминазалары мен оксидазаларының әсерінен тікелей емес дезаминденуге қатысады. АҚ-ның азотсыз қалдықтары глюконеогенезге пайдаланылады, ce6e6i бұл процесске қатысатын ферменттер бауырда көп синтезделеді. Түзілген аммиактан мочевина түзіледі. Бауырда белок биосинтезі артады.
.
100.Минералокортикостероидтар (альдостерон), химиялық табиғаты, нысана-жасушалары, әсер ету механизмі, физиологиялық эффектілері, секрециясының реттелуі. Альдостеронның гипожәне гиперфункциясы.
Минералокортикостероидтар (МКС)
МКС буйрек ycTi безінің туйірлі аймагында тузшеді Оларга альдостерон, дезоксикортикостерон (ДОК) жатады. Альдостеронның активтілігі ДОК-ка Караганда 50 есе артык.Сондыктан альдостерон агзадагы негізгі белсенді минералокортикостероид болып табылады.
Химиялык, табигаты. Холестериннен синтезделеді
Эсер ету мехаиизмі. Альдостеронның эсер ету механизмі баска стероидты гормондардың эсер ету механизміне уксас.Альдостеронның рецепторлары жасуша ішінде орналасады.
Нысана-тіндерге буйрек, ішектің кілегейлі кабаты, сілекей және Tepi бездері, куык эпителий жасушалары, Tepi, миокард, бауыр.
Физиологиялык, әcepi. Натрий, калий, хлоридтер жэне су алмасуына әсер етеді Альдостеронның эсерінен нысана жасушаларда белоктардың синтезі артады. Ол белоктар: 1) буйрек тутікшелерінің дистальды жэне жинагыш бөлімінде натрий каналдарының
белсендігін арттырып, натрий иондарынын реабсорбциясын камтамасыз eryi мумкін; 2) митохондрия ферменттерінің белсенділігін арттырады, Кребс циклінің біpкатар ферменттерінің синтезін арттырып иондар тасымалдануына кажетТі молекуласын тузуі мумкін ; 3) Na+/K+ -ATф-a3a насостарынын жумысын белсендіруі мумюн. Сонымен альдостерон буйрек тупкшелерінің дистальды жзне жинағыш бөлімінде Na+ иондарынын реабсорбциясын, К+, Н+, NH4+ иондарынын экскрециясын жэне натрий каналдарынын санын арттырады.
Тотыгу-тотыксыздану реакцияларының артуынан АТФ синтезі де кебейеді АТФ-тын ыдырауындагы энергия K+,Na+ насостарыныц жумысы үшіH колданылады. Na+ біріншілік peттiK зэрден жэне т.б. (Tepi, сілекей бездері, iшек кілегей кабагы) тіндерден канға сіңеді де мөлшері кебейеді, канның осмостык кысымы жоғарылайды, сондыктан канта жасуша аралык кеңістіктен су туседі Бул жағдай канның көлемін жогарылатып артериалды кысымын көтереді Зәрмен К+ жэне Н+ иондарынын бөлінуі артады, осылайша канның кышкылдык-сілтілік турактылыгын реггейді
Алъдостеронның секрециясының реттелуі.
1. Ренин-ангиотензинді жуйемен реттеледі Буйрек альдостерон секрециясына катысады. Организм кеп кан жогалтканда, гипотензияда (артериялық кысым темендегенде), NaCI концентрациясы темендегенде буйректің юкстатломеруларлы аппаратында ренин
тузіледі Ол кандагы ангиотензиногенге (бауырда тузілетін белок) эсер етіп оны ангиотензин-1-ге айналдырады. Ангиотензин-1 өкпенің ангиотензинге эсер eTyшi ферменттің нэтижесінде ангиотензин ІІ-ге айналады. Ангиотензин-ІІ буйрек ycTi безінің кыртыс кабатынан альдостеронның секрециясын камтамасыз етеді Альдостерон канда натрий мелшерін арттырып, кан көлемінің улгайуына жэне артериялык кысымының жогарлауына экеледі
2. Кан сарысуындагы натрийдің концентрациясы азайып, калийдің конценграциясы кебейгенде альдостеронның секрециясы кушейеді ал натрийдің көбеюі мен калийдің азаюы Kepiсіншe альдостеронның бөлінуін тежейді
3. АКТГ альдостерон секрециясын арттырады.
4. Простагландиндер жэне кинин-калликреин жуйесі
111.Қанның қалдық азоттының заттары, негізгі өкілдерінің қалыпты жағдайдағы мөлшері, түзілуінің маңызы.
Егер плазма белоктарын тұнбаға түсірсе, мөлдір белоксыз ерітіндінің құрамында азотты заттар қалды. Бұл қосылыстардағы белоксыз заттарды қалдық азот деп атайды. Қалдық азот – плазманың жалпы азоты мен белоктың азотының айырымына тең. Қалыпты жағдайда қан плазмасында RN-нің концентрациясы 15-30 ммоль/л тең. Құрамына негізінен белоктар алмасуының аралық және соңғы өнімдері кіреді.Қалдық азоттың көбеюі гиперазотемия деп аталады, ол ретенциондық және продукциондық деп екіге бөлінеді. Ретенциондық гиперазотемия – қан ағымына мөлшерде түскен азот зәрге жеткіліксіз бөлінгенде байқалады. Ол бүйрекке тәуелді және бүйрекке тәуелсіз болады. Бүйректік ретенциондық гиперазотемия бүйректің экскреторлық қызметінің бұзылуы кезінде байқалады. Бұл жағдайда қан плазмасындағы қалдық азоттың мөлшері мочевинаның көп болуына байланысты, яғни ол зәрмен шығарылмай қалғандықтан, RN мөшері де мочевинаның азоты арқылы көбейеді.Продукциондық гиперазотемия – қанға азотты заттардың көп мөлшерде түскенде және тін белоктарының күшті ыдырауында байқалады. Қалдық азоттың қан плазмасында жоғарлауы қан қоюланғанда, ағыл- тегіл іш өткенде, көп мөлшерде тер бөлінгенде болуы мімкін.
112.Қан плазманың азотсыз органикалық құрам бөліктері, қалыпты мөлшердегі, диагностикалық маңызы .
Қанның қанты негізінен глюкоза болып саналады. Қалыпты жағдайдағы мөлшері – 3,4-6,0 ммоль/л. Қан плазмасында және формалық элементтерінде глюкоза мөлшері шамамен бірдей. Қанда глюкозаның көбеюі гипергликемия деп аталады.Физиологиялық гипергликемия жеңіл сіңетін, аз уақытта тез қорытылатын көмірсуларды көп мөлшерде пайдаланғанда байқауға мүмкін. Бұл жағдай тез өтеді және аз уақытты. Патологиялық гипергликемия – қант диабетінде байқалады, бұл кһмірсулар алмасуының бұзылуына алып келіп сроғады, бүйрек үсті безінің гиперфункциясы кезінде, ми ісігінде және тағы басқа жағдайларда байқалады. Қанда глюкозаның мөлшерінің азаюы гипергликемия деп аталады, ол ұзақ ашыққанда, ауыр дене жұмысында, инсулин артық болғанда немесе инсулинді артық мөлшерде ағзаға ендіргенде байқалады. Осы жағдайлар ауыр күйге ұшыратады – бас айналып , аяқта әлсіздік , солғындық, суық тер, ми зақымданады, кома дамиды себебі нерв жасушаларына тек глюкоза ғана энергияның көзі болып табылады. Кома жиі және ұзақ уақытқа созылса, мидың жасушаларында дегенеративтік өзгерістер пайда болады.
114.Ацидоздар:түрері және пайда болу себептері,компенсациялау механизмі
Респираторлы ацидоз - бұл қанда С02-ның жиналуына байланысты болған рН-тың темендеуі. Респираторлы ацидоздың себептері: 1) тыныс алу кызметінің жеткіліксіздігі, тыныс алу орталыгының салдануы (паралич);
2) кан айналымының жеткіліксіздігі , кан ағымының кенет баяулауы нәтижесінде С02-ның қаннан шығарылуы қиындайды.
3) С02-ң жоғары концентрациясымен дем алу. Бұл кезде 6ipнeше теңелтілген механизм қосылады: СО, эритроциттерге кіpiп, сумен көмір қышқылын түзеді (Н2С03). Соңғысы протон (H-) мен гидрокарбонат-анионға (НСО; ) диссоциацияланады. Н+ эритроциттерде гемоглобинде сакталады, ал НСО ' плазмага өтіп, натриймен байланысып, қанның буферлік сыйымдылығын жоғарылататын NaHCO тузеді. Натрийдің көзі болуы мумкін: белоктық буфер; фосфат буфері осы кезде кышкылдык NaH2P04 тұз тұзеді, ол несеппен шығарылады; NaCl, хлоранионы НС03 орнына эритроциттерге түседі.
Теңгерілген реакцияларда буйректе жүректің реакциялардың маңызы зор. Бүйректе НС03' мен натрий катионының реабсорбциясы күшейеді, ал аммиак (NH3) протонды қосып алып (аммонийгенез), NH4+ түзеді. Аммоний катионы қышқылдық өнімдермен байланысып, ағзадан шыгарылады. Н ,С03артығының жоюының негізгі механизміөкпенің гипержелдеткіштігі (гипервентиляция), СО2 жиналып қалганда, гиперкапниятуғызуы мүмкін, ал ол қан айналымының бұзылуына және тыныс алу орталығының қозғыштығының жоғарылауына әкеледі, тыныс алу жиілігі жоғарылайды, С02 бөлініп шығарылады.
Метаболиттік ацидоз - жиі кездесетін ҚНТ-тің бұзылуының ауыр тypi. Қанда қышқылдық өнімдер жиналады. Бұл жағдайдың себептері
1) зат алмасуының бұзылуы. Осының негізінде қышқыл заттар шамадан тыс тузеді, мысалы, кетон денелері;
2) оттек жетіспеген кезде өте тез жэне ауыр турде дамиды. Бұл жағай қан айналымының терең бұзылуында (жүрек тоқтап қалғанда, шок (естен тану), коллапс кезінде) байқалады, бұл кезде лактат, пируват мөлшері кебейеді;
3) бүйрек ауруы кезінде оның қышқыл өнімдерді шығаруы бұзылады (шумақты сүзгілеу төмендейді);
4) ағзадан негіздік эквиваленттердің сілтілік асқорыту сөлімен шығарылуы (iш өткен кезде, ішектің жыланкөзі болғанда);
5) ағзаға көп мөлшерде қышқыл заттардың туcyi (қышқылдармен улану, артық мелшерде парентеральдық жолмен амин қышқылдарды енгізу - аргинин, лизин, гистидин)
Теңелтілу мына жолдарымен жүзеге асырылады:
- жасушааралық кеңістікпен қанға сұйықтық түседі, ол қышқыл өнімдердің концентрациясын сұйылтады;
- реакцияға бикарбонаттық буфер қатысады, ол протонмен байланысады да, Н2С03 тузеді, соның есебінен NaHC03 азаяды, бұл метаболиттік ацидоздың негізгі көpiнici;
113.Қан плазмасының бейорганикалыққұрамдас бөліктерінің маңызы. Канның буферлік жүйелерінің ҚНК сақтауға қатысуы, олардың механизмін бикарбонатты буфер мысалында көрсету
Қан плазмасының бейорганикалыққұрамдас бөліктерінің маңызы.
- Қан плазмасының осмостық қысымын қалыпты деңгейде ұстап тұрады, қалыпты жағдайда осмостық қысым 7,6 атм-қа тең.
- Суда ерімейтін белоктардың (глобулиндердің) еруіне себеп болады.
- Канның бейорганикалық құрамдас бөліктері жасушаларды шайып, жасушаның қалыпты қызмет аткаруына ықпал етеді.
Иондар арасындағы қатынас жүйке жуйесінің қызметіне, жүрек - қантамыр жуйесінің бұлшық еттерінің жиырылуына, тіндік коллагендердің ісіну қабілеттілігіне әсер етеді.
- Канның рН-н тұрақтандырады. Буферлік жуйелердің түзілуіне қатысады:
а) бикарбонаттық буфер жуйесі - H2C03/NaHC03 (1/20) - қан плазмасының ең күшті жүйесінің 6ipi , қанның буферлік сыйымдылығының 10%-н құрайды;
б) фосфаттық буфер - NaH2P04/Na2HP04 (1/4) - аталып кеткен буфер жүйелердің бөлімі көбірек болады, өйткені ағзада қышқылдық өнімдер көбірек түзіледі;
в) белоктық буферН*белок/Na*белок - күшті буферлік жуйе, ce6e6i белоктар құрамында көп мөлшерде қышқылдық және негіздік қасиет көрсететін радикалдары болады;
г) гемоглобиндік буфер (Н*Нb/К*Нb) және оксигемоглобиндік буфер (Н*НbO2/К*НbO2) - ең күшті буфер жүейсі, бикарбонаттық буфер жуйесінен 9 есе жоғары. Бұл буфердің жұмысы оттек пен көмірқышқыл газын тасымалдау рөлімен байланысты.
119. Қалыпты жағдайдағы зәрдің физикалық қасиеттері (мөлшері, меншікті салмағы, рНы, түсі, иісі, мөлдірлігі) және патология кезінде өзгеруі. Зәрдің қалыпты пигменттері.
Мөлшері: Тәулігіне бөлінген зәрдің көлемін диурез деп атайды, ол шамамен 2 л-ге тен. Ер адамдарда Зәр көбірек бөлінеді.
Диурездің жогарылауы — полиурия деп аталады, физиологиялык, (көп мелшерде су ішкенде, суйык тамак ішкенде, кыста - ce6e6i тер аз бөлінеді және патологиялық (кантты жэне кантсыз диабет, созылмалы нефрит, пиелонефрит, iciнy кайтканда) деп екіге бeлiнeдi.
Олигоурия – бөлінген зәр мөлшерінің төмендеуі. (тәулігіне 500 мл-ден аз). Олигоурия физиологиялык және патологиялык деп бөлінеді. Физиологиялык олигоурия- шөлдегенде, сұйықтықты аз қабылдағанда, ауыр кара жумыста, жазда терінің көп бөлінуі арқылы жүреді. Патологиялык олигоурия- калшылдау, диарея, токсикоз, iciнy, жедел бүйрек жетіспеушілігі жағдайында болады.
Анурия – зәрдің бөлінуінің тоқтауы. Анурия: а) пререналдык – соққы кезінде, күйікте, сұйықтықты жоғалтканда, журек жетіспеушілігінде, гипотензияда;б) реналды - буйрек ауруларында; в) постреналдык –зәр шығару жүйесінің аурулары кезінде байқалады.
Никтурия –зәрдің күндізге қарағанда түнде көбірек бөлінуі, журек жумысының бузылуында, жедел нефритте байқалады.
Меншікті салмағы немесе зәрдің салыстырмалы тығыздығы зәрдің калыпты жағдайдағы меншікті салмағы- 1,010-1,025. Зәрдің меншікті салмағы диурезге кepi пропорционал. Полиурияда меншікті салмағы темендейді, олигоурияда – керісінше, жогарылайды. Патология кезінде бул аракатынас өзгереді. Егер зәрдің меншікті салмағы олигоурия кезінде төмендесе, буйрек кызметінің ауыр бузылысы және оның қатерлі белгісі болып табылады.
Tүci: калыпты жагдайда Tүсі: акшыл-сарыдан кою сарыга дейін озгереді. Зәрдің түсін құрамындағы пигменттер қалыптастырады: урохром, урозеин, уроэритрин, стеркобилин. Патология кезінде зәрде қан болса кызыл тус пайда болуы мүмкін. Зәрде жасыл түстің пайда болатынының себебі - зәрде өт пигменттері көп мөлшерде болғанда, метгемоглабин зәрге түскенде коңыр туске боялады. Зәрдің құрамында көп мөлшерде фосфаттар, липидтер және ipiң болса зәр ақ тусті болады.
Алкаптонурия, гомогентизин қышкылының және оның метаболиттерінің бөлінуі кезінде зәр қара туске боялады.
Мөлдірлік : жаңадан шығарылған зәр мөлдір, ал тұрып қалған кезде жеңіл лай сиякты бұлыңғырт болады.
Иісі - Қалыпты жағдайда зэрдің арнайы жаңадан кайнаткан еттің иісіне ұқсайтын иісі болады. Жаман жағымсыз иіс сарымсақты, ақжелкекті (хрен) жегенде пайда болады. Жаңа шыққан зәрде аммиактың иісі цистит, пиелит аурулары кезінде пайда болады. Жеміс-жидектің иісі қант диабеті кезінде пайда болады. Нәжістік иіс өт –қабы-ішек аралық тесігі кезінде пайда болады.
Зәрдің PH-ы- 5-7 арасында әлсіз қышқылдан бейтарап ортаға дейін болады. Осы Ph натрий фосфаты мен( NaH2PO4) калий фосфаты ( KH2PO4) болуымен қамтамасыз етіледі. Ет тағамын және дәндерді жегенде дәндердің қышқылдығы жоғарлау болады
Сау адамдарда зәрдің химиялық құрамы:
Несеп –судан және құрғақ қалдықтан ( 42-62 г/тәул) тұрады
Құрғақ қалдық -органикалық заттардан ( 34-45 г/таул) және бейорганикалық заттардан(15-25 г/ таул) тұрады.
Органикалық заттар- азотты азотсыз болып бөлінеді.
Азотты заттарға белок алмасуының соңғы өнімдері жатады. Зәрдің азотының 90-93% мочевина құрайды.
Зәрдің органикалық құрамдас бөліктері
Мочевина: Тәулігіне 25-30 гр бөлініп тұрады. Зәрде мочевинаның мөлшері тағамдағы белоктың мөлшеріне байланысты. Зәрмен бөлініп шығатын, азотты заттардың мөлшері белокты тағамды көп қабылдағанда немесе тіндердің ыдырауында жоғарлайды, ал шығарылуы бұзылғанда төмендейді.
Креатинин: тәулігіне 1,5-2,4 гр бөлінеді. Креатининнің бөлінуі бұлшық еттің массасымен бүйректің бөлінуі массасымен анықталады.
Несеп қышқылы- пуриндік негіздердің алмасуы соңғы өніміт Тәулігіне 1,2 гр бөлінеді. Сүтте, жұмыртқада, бауырда ,етте, қызанақта- көп болады.
Азотсыз заттар- зәрде аз болады. Оксолоацетат, Цитрат, лактат, май , валерьян, янтарь қышқылдар осы топты құрайды.
Бейорганикалық заттар-негізділері: хлоридтер (10-15 г/таул), сульфаттар ( шамамен 3 г/таул) фосфаттар (1-5 гр/таул)
120. Дені сау ересек адам зәрінің химиялық құрамы, зәрдің қалыпты компоненттерінің арту және төмендеу себептері.
Қалыпты зәрде бұл аз мөлшерде кездеседі. Сапалық реакциялармен анықталмайды.
Протеинурия –несептен қалыпты мөлшерден (50мг/тәу.) артық ақуыздың бөлінуі. Бүйрек зақымдануының ең жиі белгісі, бірақ кейде сау адамдарда да бола береді. Ересек адамға тәулігіне 30-50мг ақуыз бөлінуі физиологиялық жағдай болып саналады. Айқын лейкоцитурия немесе гематурия болған жағдайда ақуызға оң реакция несептің ұзақ тұрып қалуында формалық элементтердің ыдырауы нәтижесінде пайда болады, мүндай жағдайда ақуыз мөлшері тәулігіне 0,3г. асса, паталогиялық болып табылады. Егер несепте иодты контрастты заттар, көп мөлшерде пенициллин немесе цефазолин аналогтары, сульфаниламидтер метаболиттері анықталса, ақуыздық тұнбалық сынамалар жалған оң нәтиже береді. Тәулігіне 3 г. асатын протеинурия нефротикалық синдромның дамуына әкеледі.
Гематурия- зәрде қанның пайда болуы. Глобинурия және эритроцитурия болуы мүмкін. Бұл жағдайларда тұнбасын микроскопия әдісімен ажыратуға болады. Гематоурия бүйрекпен несеп шығару жолдары зақымдалғаанда, бүйректе және несеп шығаратын жолдарда ісіктердің ыдырауы, зәр- тас ауруларында, бүйректе операция жасалғаннан кейін, гломерулонефрит кезінде байқалады.
Гемаглобинурия-ауыр гемолитикалық анемияда, қатты уланғанда, сепсис күйік кезінде пайда болады.
Глюкозаурия- зәрде глюкозаның болуы. Ол бүйрекке тәуелді және бүйрекке тәуелсіз болып келеді. Бүйрекке тәуелді глюкозаурия кейбір нефрит түрлерінде, бүйректе глюкозаның раебсорбциясы жүргізетін ферменттің кемшілігінде байқалады. Бүйрекке тәуелсіз глюкозаурия қанда глюкозаның деңгейі бүйрек межесінен асып кеткенде байқалады.
Кетаноурия-Қант диабеті кезінде, ашығуда, тиреотоксикозда, церебральді кома кезінде,ауыр қалтырау кезінде, алкагольді интоксикация кезінде гиперкатехолемияда байқалады.
Креатинурия- Креатиннің зәрмен көп шығуын айтамыз. Креатинурия - миопатиялар, бұлшықеттің прогрессияланған дистрофиясы кезінде байқалады. зәрде креатиннің пайда болуы гипертиреоз кезінде, аддисон ауруы кезінде, қантты диабет кезінде, инфек аурулар, күйіп қалу, белоктық ашығу, қызыл волчанка және Е витамині жетіспеген кезінде байқалады.
Өт пигементтері: Тікелей билирубин- зәрде паренхиматозды және механикалық сарғыштанудан пайда болады.
Зәрдің диастазасы: асқорыту сөлінде (  және липаза) Гидролазалар- ферменттерді қосатын термин. Панкреатитте және паротитте зәрде анықталады. Диастазаға несеп талдауы дамудың ерте кезеңдерінде панкреатикалық ауруларды анықтауға мүмкіндік беретін зерттеудің көрсеткіш әдісі болып табылады. Негізінен емшарадан өту пациентке ұйқы безі қызметінің бұзылғанын куәландыратын клиникалық көріністер аясында тағайындалады. Қалыпты мәндер Сонымен, зәрдің диастазасы сәл жоғары болды. Енді осы талдаудың қабылданған нормаларын қарастыру керек, өйткені олардың төмендеуі немесе ұлғаюы ұйқы безінің қызметінің бұзылуының тікелей көрсеткіші болып табылады.
және липаза) Гидролазалар- ферменттерді қосатын термин. Панкреатитте және паротитте зәрде анықталады. Диастазаға несеп талдауы дамудың ерте кезеңдерінде панкреатикалық ауруларды анықтауға мүмкіндік беретін зерттеудің көрсеткіш әдісі болып табылады. Негізінен емшарадан өту пациентке ұйқы безі қызметінің бұзылғанын куәландыратын клиникалық көріністер аясында тағайындалады. Қалыпты мәндер Сонымен, зәрдің диастазасы сәл жоғары болды. Енді осы талдаудың қабылданған нормаларын қарастыру керек, өйткені олардың төмендеуі немесе ұлғаюы ұйқы безінің қызметінің бұзылуының тікелей көрсеткіші болып табылады.
Диастаза нормасының нұсқалары науқастың жыныстық тиістілігіне, сондай-ақ оның жасына байланысты бірнеше түрленуі мүмкін. Ферментативті зат деңгейінің көрсеткіші ретінде мккат/л, катал, кат немесе шартты бірлік мәні болады. Бұл не? Берілген терминдер диастаза белсенділігінің бір бірлігін белгілеу үшін қолданылады. Тәжірибеде жиі катал мәні қолданылады.
Орташа алғанда, диастазаның мөлшерлері дені сау адамның зәрінде 10-нан 160 бірлікке дейін ауытқуы мүмкін. Олар кіші жастағы балаларда болуы керек. Қарт адамдарда неғұрлым жоғары мән байқалады
Диастазды көтеретін және түсіретін жағдайлар:
1. Бүйрек жетіспеушілік. Бүйрек жұмысының бұзылуы
2. Панкреатит.
3. Холецистит.
4. Спирттік сусындардың әсерінен асқазан сөлінің, бауырдың функциясының бұзылуы
5. Жедел аппендицит, перитонит.
6. Ауыз қусының жедел асқыну процесстері асқана ас қорыту жүйесіне әсер ету салдарынан
7. Паротит.
8. Қант диабеті.
121.Зәрдің патологиялық компоненттері. Олардың зәрде пайда болу себептері. Глюкозаныңбүйрек межесі. Зәрдің патологиялық компоненттерін ашатын сапалық реакцияларды атапшығу. Зәр диастазасы, түсінік, себептері.
Зәрдің патологиялық құрамдас бөліктері
Қалыпты зәрде бұл заттар болар-болмас мөлшерде кездеседі және сапалық реакциялармен анықталмайды. Оларға қан (гемоглобин мен эритроциттер), өт пигменттері, глюкоза, кетон денелері жатады.
Протеинурия - зәрде белоктың болуы. Нағыз (бүйрекке тәуелді) және жалған (бүйрекке тәуелсіз) болып бөлінеді. Нағыз протеинурия бүйрек шумақтарының қызметі бұзылғанда байқалады.
Гематурия - зәрде қанның пайда болуы (гемоглобинурия және эритроцитурия болуы мүмкін, бұл жағдайларды тек зәрдің тұнбасын микроскопия әдісімен ғана ажыратуға болады). Гематурия бүйрек пен несеп шығару жолдары закымданғанда, бүйректе және несеп шығаратын жолдарда ісіктердің ыдырауы кезінде, зәр-тас ауруында, бүйрекке операция жасағаннан кейін, гломерулонефрит кезінде, нефрит кезінде байқалады. Нәрестелерде физиологиялық гематурия – бүйректің зәрқышқылдық инфарктісі болған кезде байқалады және өмірдің 10-шы күнінде
аяқталады.
Гемоглобинурия - ауыр гемолитикалық анемияда, қатты уланғанда, сепсис, күйік кезінде болады.
Глюкозурия - зәрде глюкозаның болуы. Ол бүйрекке тәуелді және бүйрекке тәуелсіз болып бөлінеді. Бүйрекке тәуелді глюкозурия кейбір нефрит түрлерінде, бүйректе глюкозаның реабсорбциясын жүргізетін ферменттің (туа біткен нефрит)
кемшілігінде байқалады. Бүйрекке тәуелсіз глюкозурия қанда глюкозаның деңгейі бүйрек межесінен асып кеткенде байқалады (8,8 ммоль/л). Көбінесе глюкозаның
қанда көп болуы қант диабеті мен басқа эндокриндік ауруларда байқалады.
Кетонурия - қант диабет кезінде, ашығуда, тиреотоксикозда, церебралды кома кезінде, ауыр қалтырау кезінде, алкогольді интоксикация кезінде, гиперинсулинизм кезінде, гиперкатехолемияда, операциядан кейін байқалады.
Өт пигменттері - тікелей билирубин - зәрде паренхиматозды және механикалық сарғыштануда пайдаболады. Бауыр патологиясы кезінде зәрде уробилин пайда болуы мүмкін.
Креатинурия - қалыпты жағдайда ересек адамдардың зәрінде креатин болмайды. Креатинурияның физиологиялық түpi (нәрестелік және қарт адамдарда кездесетін креатинурия) бар. Нәрестелердің ағзаларында креатин түзіледі, бipaқ олардың бұлшық eттepi дамымағандықтан, креатинді толығымен пайдаланбайды. Сондықтан қандағы креатиннің мөлшері бүйрек межесінен артып, зәрде пайда болады. Қарттарда креатин әлі түзіледі бipaқ бұлшық eттepi атрофияланып (сему) креатинді толығымен пайдаланбайды. Осындай креатинурия көмірсуларсыз диетада, тағаммен шикі еттi көп мөлшерде пайдаланғанда байқалады. Патологиялық креатинурия белокты ашығуда, үлкен сынықтың бітуінде, бірқатар операциялық әрекеттерде, бұлшық ет дистрофияларында. Сонымен қатар креатинурия Е витаминнің авитаминозында, қант диабеті кезінде, тіндер көп ыдырағанда. Креатиннің көп мөлшерде бөлінуі гепатит кезінде және кастраттарда байқалады.
Амин қышқылдарының мөлшері бүйрек түтікшелері ауруларында артпайды, өйткені олар реабсорбцияланбайды.
Фенил кетонурия - тұқым қуалайтын ауру, бауырда гидроксилазалар болмағанда зәрде фенилаланин пайда болады.
«Зәрдің диастазасы» - асқорыту сөліндегі (а-амилаза, липаза) гидролазалар-ферменттерді қосатын термин. Панкреатитте (амилаза және липаза) және паротитте (амилаза) зәрде анықталады.
Диастазды жоғарылату
Кейбір дәрі-дәрмектерді ұзақ уақыт бойы қабылдау, күшті болу.
Ішкі ішімдік ішу арқылы ішу және ішке зиян келтіру, алкоголь бар.
Қант диабеті кезінде ұйқы безі де әсер етеді, бұл диастаздың бірдей көрсеткіштері бойынша жиі кездеседі, және екінші ауру.
Ішкі ағзалардың басқа қабыну аурулары, ас қорыту процесіне тікелей қатысады.
Қабыну процесінің болуы фермент деңгейінің салыстырмалы түрде аз мөлшерде өсуіне әкелуі мүмкін, сілекей безінің ауруы. Алайда, бұл патологияның қатысуымен сұйықтықтың түсі, ереже бойынша, теріс бағытта өзгермейді.
Бүйрек жеткіліксіздігі диастаздың жоғарылауымен сипатталады, зәрдің түсі жиі өзгереді, сұйықтық көбірек турбулентті және қараңғы болады.
Диастазды азайту
Зәрдің құрамындағы диастаздың деңгейінің төмендеуі панкреатиттің болуын да көрсете алады. Бұл фактке байланысты, бұл аурудың ұзын бағдары аясында ұйқы безінің қызметі айтарлықтай азаяды, бұл оның ферментті қалыптастыру функциясының төмендеуіне әкеледі.
122.Дәнекер тінінің химиялық құрамының схемасын сызу. Дәнекер тінінің атқаратынқызметтерін атап шығу.
Дәнекер тінінің құрамы
-Жасушалардан
- Жасушааралық заттан(матрикс) Матрикс саны жасушаларға қарағанда өте көп болады. Оның құрамы өте күрделі: фибриллярлық құрылымы және негізгі заты бар. Фибриллярлық құрылымға коллагенді, эластинді, ретикулярлы талшықтар жатады. Негізгі заты-матрикстің аморфты компоненті, бұл компоненттің арасына жасушалар мен талшықтар орналасқан.
Дәнекер тінінің атқаратын қызметтері:
Тіректік қызметі - бұл дәнекер тінінің маңызды қызметі, ceбeбi осы тін басқа тіндерге қарағанда тығыз болып келеді. Соның арқасында ол ағзаны тек байланыстырып қана қоймай, сонымен қатар ағзаға тірек
болады, бұлшық ет жүйесінің қозғалтқыш кабілетін қамтамасыз етеді.
Метаболиттік қызметісаналуан. Дәнекер тіні ағзаның iшкi ортасы болады, қан мен лимфа тамырлары арқылы барлық тіндерді қоректік заттармен, метаболизм өнімдерімен қамтамасыз eтiп (трофикалық кызметі), зат алмасуына белсенді катысады. Бұл қызметін көбінесе дәнекер тінінің жасушалары мен протеогликандар атқарады.
Қорғаныш (тосқауыл) қызметі. Дәнекер тіні тepi қабаттарын (сыртқы жабынды) түзеді, олар ағзаны сыртқы ортаның әсерінен және түрлі зиянды заттар мен микробтардың енуінен қорғайды. Дәл осы қызметін iшкi мүшелердің қаптары мен серозды қабықтары да атқарады.Жасушалық деңгейде бұл қызмет макрофагтар, фибробластар, қан тамырларының эндотелий жасушалары арқылы жүзеге асырылады. Фагоциттік активтілік арқылы олар бөгде және эндогендік заттарды сіңіреді.
Структура түзушi (құрылымдық) қызметi. Бұл қызметі эмбрионалдық кезеңде өте қарқынды жүреді Ол тек дәнекертіндік қана емес, сонымен бipгe бұлшық еттердің жэне эпителий жасушаларының көбеюіне бағытталған коллаген мен гликозамингликандардың реттеуші әсepi арқылы жүзеге асырылады. Бұл қызметі бала туылғаннан кейін де жүре береді.
Penapamuвтік қызметі - тіндерде экзо- немесе эндогенді факторлардың: жарақат немесе жұқпалы аурулардың әсерінен пайда болған тіннің кемістігін жою және тіндерді қалпына келтіру. Дәнекер тінінің бұл кызметі өте ерекше. Жетілген ағзадағы көптеген
дифференциацияланған тіндердің пролиферацияға(жасушаның көбею қабілеті) түсу қабілеті шектелген. Дәнекер тіні ересек адамдарда да қайта қалпына келу (регенерация) кабілетін сақтайды. Сондықтан ол
тек дцнекер тінініңкемістігінің орнын толтырып қана қоймай, сонымен қатар паренхиматоздық мүшелердің кемістігінің де орнын толтырады (орынбасушылық регенерация).
123.Дәнекер тініндегі зат алмасу ерекшеліктері. Дәнекер тінінің жасқа байланысты өзгерістері.Дәнекер тініндегі метаболизмнің реттелуі. Коллагеноз жағдайында дәнекер тініндегі негізгі өзгерістер.
Дәнекер тінінің алмасу қызметі өте көп түрлі және күрделі
Дәнекер тініндегі зат алмасуының ерекшеліктері:
- дәнекер тінінде басқа тіндерге қарағанда зат алмасу қарқындылығы біршама төмен;
- дәнекер тініндегі әржақты кызметтік қоры бар. Патологиялық жағдайда (мысалы жарақат кезінде) дәнекер тінінің жасушаларында жасушааралық заты түзілетін макромолекулалардың, талшықтардың ерекше синтезі күрт күшейеді.
- дәнекер тінінде осы тінге тән арнайы белоктар: коллагендер, эластиндер, протеогликандар мен ГАГтар түзіледі
- дәнекер тінінде липидтердің олардың алмасуына қатысатын ферменттердің де саны аз;
көмірсулар, амин қышқылдары, яғни төмен молекулалық метаболиттер энергия көзі болып табылады. Гликогеннің ыдырауы амилолитикалық жолмен жүзеге асады;
-дәнекер тінінде арнайы реакциялар жүреді, оларға гликозилдену, гидроксилдену, сульфаттану және т.б. реакциялар жатады;
- дәнекер тінінде протеогликандардың негізгі компоненттерінің синтезі - урон қышқылдары мен аминқанттар түзіледі. Аталған заттар глюкозадан екі жол арқылы түзіледі. бірінші жолы - глюкозо-1-фосфаттан УТФ-ң қатысуымен урон қышқылы
түзіледі. Екінші жолы - фруктозо-6-фосфаттан глутамин мен УТФ-ң қатысуы арқылы аминоқанттар түзіледі. Сульфотрансферазаның әсерінен және фосфоаденозилсульфаттың қатысуымен урон қышқылдары мен гексозаминдерге сульфотоптар енгізіледі.
Дәнекер тінінің жасқа байланысты өзгерістері.
Балалардың дэнекер тінінде матрикске қарағанда жасушалар көп болады, сондықтан оларда қайта қалпына келу кабілеті жоғары болады. Жалпы зат алмасу жылдамдығы да балаларда жоғары. Балаларда лизин мен пролиндігидроксилдендіретін ферменттердің активтілігіересек адаммен салыстырғанда 8-10
есе жоғары, сондықтан коллаген тез түзіледі. Бала ағзасында ГАГ-тармен байланысқан су көп болады. Жас ұлғая келе дәнекер тінінің барлық тіндеріне
тән жалпы өзгерістерге су мөлшерінің азаюы және негізгізат/талшыктар арасындағы арақатынастың өзгepyi жатады. Коллаген мөлшерінің көбеюі және
ГАГ-тардың санының кемуімен бұл коэффициент азаяды. Алдымен гиалурон кышқылының мөлшері азаяды, бұл байланысқан судың мөлшері де төмендейді, бұл жағдай ГАГ-тардың өзара қатынасын өзгертуге әкеледі. Сол кезде молекулалық елеуштің кеуектілігі бұзылады. Коллагеннің физико-химиялық қасиеттері
өзгереді: оныңсерпімділігі мен iciнyгe қабілеттілігі төмендейді, коллаген талшықтарының құрылымдық тұрақтылығы жоғарылайды, коллагеназаға резистенттілігі пайда болады, коллагеннің суда еритін фракцияларының мөлшері азаяды.
Дәнекер тінінің метаболизміне витаминдер мен гормондар әсер етеді. Мысалы, тироксин гиалурон қышқылының деполимеризациялануын жүргізеді, ал
глюкокортикостероидтар фибробластардың коллаген мен ГАГ-тардың синтезін тежейді. Сонымен қатар фибробласттардың биосинтетикалық активтилігі тежеліп қоймай, коллагеннің ыдырау үрдістері артады. Сондықтан глюкокортикостероидтар коллагеноздарды емдеу үшін қолданылады. Минералокортикостероидтар, кepiciншe, фибробластардың пролиферациясын стимулдеп, негізгі затының биосинтезін күшейтеді. С витамині де осындай әсер көрсетеді және пролин мен лизиннің гидроксилдену реакциясына себепші болып, коллагеннің синтезін арттырады.
Дәнекер тінінің ауруларының бipi – коллагеннің алмасуының бұзылуы, бұл кезде коллагеноздар дамиды. Бұл ауруға дәнекер тінінің диффуздық зақымдануы, макрофагтардың миграциясы, аурудың асқынуы және аутоантиденелердің пайда болуы тән. Осындай ауруларға ревматизм (кұяңжел), склеродермия, жүйелі
қызыл жегі, ревматоидты артрит және тағы басқа аурулар жатады.
124.Сүйек тінінің химиялық құрамының схемасын сызу. Негізгі өкілдерін атап шығу. Сүйектінінің атқаратын қызметтерін атап шығу. Гидроксиапатиттің кеңістіктік құрылысы, апатиттің басқа түрлерінің құрылысы туралы түсінік. Минералдану мен минералсыздануына ықпал ететін факторлар. Сүйек тінінің гомеостазын реттеудегі витамин мен гормондардыңролі
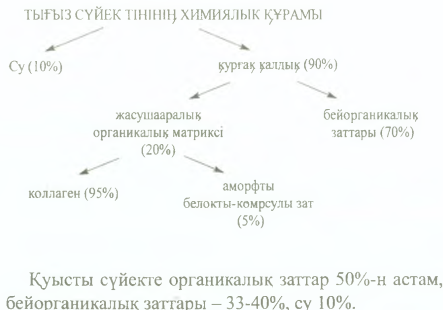
Сүйектінінің атқаратын қызметтері
Қорғаныштық - сүйек организмінің негізгі қаңқасы болып саналады, оған бұлшық ет пен iшкі мүшелер байланысады.
Сүйек бұлшық ет, буын және байламдармен бірігіп тipeк-қимыл кызметін атқарады.
Метаболиттік - сүйек тіні заттар алмасуына, әcipece минералды заттардың алмасуына қатысады.
Минералды заттардың қоры - сүйек тінінде кальцийдің 99%, фосфордың 87%, магнийдің 50%, натрийдің 46% және т.б. заттар қорға жиналады.
Реттеуіш – ішкі ортаның гомеостазын (иондық құрамын, кышқыл-сілтілік тепе-тендікті (қанның рН-ын)) сақтайды. 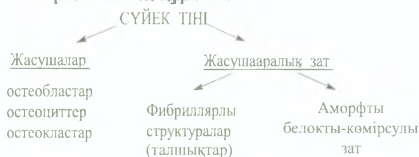 Сүйек тінінің минералдық бөлігін гидроксилапатит кристаллдары - Са10(РО4)6(ОН)2 мен аморфты кальций фосфаты - Са3(РО4)2 құрайды. Гидроксилапатиткристаллдарыныңпішіні пластина немесе таяқшатәрізді болады. Бұл қосылыстың структурасыөзін
Сүйек тінінің минералдық бөлігін гидроксилапатит кристаллдары - Са10(РО4)6(ОН)2 мен аморфты кальций фосфаты - Са3(РО4)2 құрайды. Гидроксилапатиткристаллдарыныңпішіні пластина немесе таяқшатәрізді болады. Бұл қосылыстың структурасыөзін
қоршаған тіндік сұйықтықтарға иондарды оңай бepiп,оларды өзіне сіңіре алуға бейімделіпқұрылған. Аморфты кальций фосфаты Са2+ және фосфат иондарының лабильдіқоры деп есептеледі. Аморфты кальцийфосфатының мөлшері нәрестелерде көп, ал жетілгенсүйекте кристалдық гидроксилапатит басым болады.
Егер сүйекте радиоактивті стронций жиналса, ісіктің пайда болуына әкеледі. Стронций аппатиттері Ca9Sr(P04)6(0H )2 сүйекті сынғыш етеді.
Егер фосфор қышқылының бip қалдығы бикарбонат анионына алмасса, онда бикарбонатты апатит Са10(РО4)5НСО3(ОН)2 түзіледі.
Минералдану мен минералсыздануынаықпал ететін факторлар
-тек коллаген талшықтарыбар кезде ғана мүмкін болады.
-€-амин топтары, ковалентті байланысқан фосфаттар, лизосомалық протеиназалар, остеобласттардың минорлы коллаген емес кальций байланыстырушы белоктары қатысады
Сүйек тініндегі кальций мен фосфаттардың реттелуіне паратгормон, кальцитонин, Д, А, С витаминдері катысады. Сүйек тінінің өcyi мен дамуына А витамині әсер етеді. А витаминінің жеткіліксіздігі кезінде хондроитинсульфаттың синтезі бузылатыны анықталган. С витамині, гидроксилазалардың коферментi ролін атқара отырып, коллаген синтезі үшiн қажет оксипролин мен оксилизиннің түзуіне қатысады.С витаминің жетіспеушілігі коллагеннің құрамы мен беріктігінің өзгеруіне және сүйек тініндегі ізбестену үрдітерінің бұзылуына әкеледі.
СТГ (өсу гормоны), пролактин, инсулин, жыныс гормондары сүйек тінінің түзілуін арттырады. Глюкокортикостероидтар сүйек жасушаларының бөлінуі мен қызметін, коллаген синтезін, ішекте кальцийдің сіңірілуін, бүйректе кальцийдің реабсорбциясын бәсеңдетеді, остеопороздың дамуына және сүйек тіні массасының азаюына әкеледі.
125.Сүйек тініндегі гомеостазды реттеудегі витаминдер мен гормондардың рөлі
Сүйек тініндегі кальций мен фосфаттардың реттелуіне паратгормон, кальцитонин, Д, А, С витаминдері катысады.
Сүйек тінінің өcyi мен дамуына А витамині әсер етеді. А витаминінің жеткіліксіздігі кезінде хондроитинсульфаттың синтезі бузылатыны анықталган.
С витамині, гидроксилазалардың коферментi ролін атқара отырып, коллаген синтезі үшiн қажет оксипролин мен оксилизиннің түзуіне қатысады.С витаминің жетіспеушілігі коллагеннің құрамы мен беріктігінің өзгеруіне және сүйек тініндегі ізбестену үрдітерінің бұзылуына әкеледі.
СТГ (өсу гормоны), пролактин, инсулин, жыныс гормондары сүйек тінінің түзілуін арттырады. Глюкокортикостероидтар сүйек жасушаларының бөлінуі мен қызметін, коллаген синтезін, ішекте кальцийдің сіңірілуін, бүйректе кальцийдің реабсорбциясын бәсеңдетеді, остеопороздың дамуына және сүйек тіні массасының азаюына әкеледі.
126.Эритроциттердің химиялық құрамының схемасын сызыңыз. Газдардың (оттектің жәнекөмірқышқыл газының) тасымалдануы. Гемоглобиндік және оксигемоглобиндік буферлікжүйелер. Гемоглобиннің туындылары. Олардың жұтылу спектрлері.
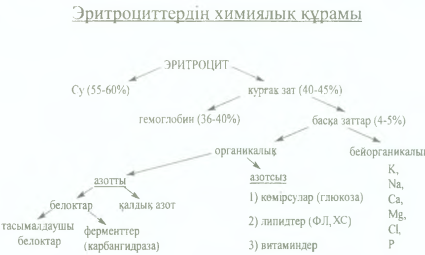
Гемоглобиннің оттекті тасымалдаудағы ролі
Оттектің гемоглобинге косылуы үшін келесі шарттар орындалуы қажет:
1. Оттектің парциалдық қысымы жоғары болуы керек (сынап бағанасы бойынша 100 мм-ден төмен болмауы керек).
2. Көмірқышқыл газының парциалдық қысымы төмен болуы керек.
3. Орта аздап қышқыл болу керек.
4. Температура төменірек болуы керек.
5. 2,3-дифосфоглицераттың концентрациясы төменірек болуы керек.
Осы жағдайлардың барлығы өкпеде болады және гемоглобиннің оксигенденуіне себепші болады (НЬ +О2 -> НЬО2). Оксигендену кезінде глобиннің В-тізбектерінің жақындауы нәтижесінде гемоглобин жиырылады, ал оксигемоглобин ыдырағанда - созылады (кеңейеді). Биохимик Перутц осы қасиеттеріне сүйене отырып, гемоглобиннің молекулаларын «дем алушы» деп атады.
Гемоглобиннің комірқышқыл газын тасымалдаудағы ролі
Зат алмасу процестері нәтижесінде түзілген Көмірқышқыл газы жасушалардан вена қанының плазмасына өтеді. Аз бөлігі қан плазмасында ipiгeнкүйде қалады. Heгiзгi бөлігі эритроциттерге өтіп, сондағыкарбангидраза ферментінің әсерінен көмір қышқылын түзеді. СО2+Н2О->Н2СО3
Kөмip қышқылының 5-10%-ы диссоциацияланып, протон мен гидрокарбонат анионын түзеді. Гидрокарбонат ионы қан плазмасына кайта отеді. НСО3-
белок буферімен әрекеттесіп, NaHCО3– бикарбонат буфері жүйесінің бөлімін түзеді. Қалған көмірқышқылы (90-95%) өкпеге тасымалданып, тыныс арқылы сыртқа шығарылады. Kөміp қышқылының 10-15% карбгемоглобин түзілу арқылы тікелей жолмен тасымалданады. Карбгемоглобиннің түзілуі және ыдырау шарттары оксигемоглобиннің түзілуі мен ыдырауының шарттарына қарама-қарсы болады. Kөмip қышқылының нeгiзгi бөлігі (80% шамасында) гемоглобиндік буфер жүйесі арқылы тасымалданады (тікелей емес жол). Вена канының эритроциттерінде гемоглобиндік буфері Н*НЬ/К*НЬ болады, ол өзінің бөлімімен көмip қышқылын байланыстырып, оны нейтралдандырады. Бұл реакцияның жүруі мүмкін, ceбeбiН2С03, гемоглобинге қарағанда күштірек қышқылжәне KHb-нен K+ ығыстырады.
Гемоглобин өкпеге түскеннен кейін оксигемоглобин буферiн түзеді ННЬО2/КНЬО2. Оксигемоглобин қышқылы көмip қышқылына қарағанда күштірек, сондықтан КНСО3-пен әрекеттесіп, құрамынан көмip қышқылын ығыстырады. Kөмip қышқылы ыдырап, көмip қышқыл газы тыныс жолы арқылы сыртқа шығарылады
Гемоглобиннің туындылары
Оксигемоглобин (НЬО) - артериялық қанның құрамында болып, оның ашық қызыл түсін камтамасыз етеді. Оксигемоглобиннің жұтылу спектрі: спектрдің сарғыш-жасыл бөлігінің Д және Е Фраунгофер сызықтары арасындағы екі жіңішке күңгірт жолақ.
2. Карбгемоглобин (НЬС02) - көк тамыр қанында түзіледі және қанға күрең-кызыл түс береді. Оның өзіне тән жұтылу cпeктpi жоқ, өйткені СО2 гемге емес, глобинге қосылады. Көк тамырлық қанды спектроскопиялау әдісімен зерттегенде, спектрдің
сарғыш-жасыл бөлігінде бip қалың күңгірт жолақ көрінеді, ол НЬ-ның жұтылу cпeктpi.
Гемоглобиннің патологиялық туындылары
1. Карбоксигемоглобин (НЬСО) – гемоглобиннің тicтi газбен қосылуы нәтижесінде түзіледі. Карбоксигемоглобин - бұл әлсіз диссоциацияланатын, оттекпенбайланыспайтын, бepiкзат. Карбоксигемоглобиннің жұтылу cпeктpi де оксигемоглобин сияқты спектрдің сарғыш-жасыл бөлігінде eкi жіңішке күңгірт жолақ.Бірақолар шамамен қызыл жаққа қарай ығысқан.
2. Метгемоглобин (НЬОН) - күшті тотықтырғыштардыңәсерінен түзіледі. Соның нәтижесінде гемоглобиннің құрамындағы тeмip үш-валенттіге дейін тотығып және гидроксил тобымен бepiкбайланысады. Метгемоглобиннің жұтылу cneктpi – үш жолақ - сарғыш-жасыл бөлігінде eкi жіңішке және қызыл бөлікте бip жолақ.
3. Гликозилденген гемоглобин - қанда глюкозаның деңгейі жоғарлауы гликозилденген белоктардың жәнеолардың гликозилденген туындыларының жиналуына әкеледі. Глюкоза көбейген кезде кез келген АҚ-ның
NH2-Toбынa ферментсіз қосылып, Шифф нeгiздepiн (альдимин) түзеді. Бұл негіздер тұрақсыз болады. Альдимин кейін өзгерістерге ұшырап, тұрақты кетоаминдерге айналады. Түзілген кетоамин белокпен байланысқан күйінде өмірбойы қалады. Ол белоктыңҚacиeттepi мен сапасын өзгертеді.
Қанның, көз жанарының, бүйректің, жүйке тінінің,қан тамырларының белоктары гликозилдену реакциясына ұшырайды.
127.Аралас сілекейдің химиялық құрамы, физико-химиялық қасиеті, биологиялық рөлі
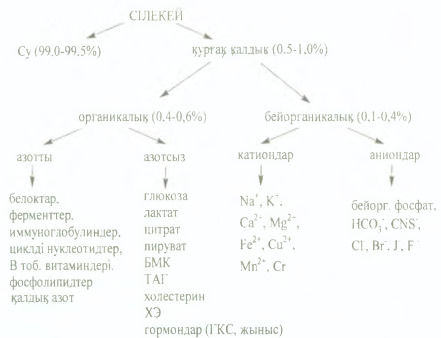 Ересек адамда тәулігіне 1-2 л сілекей түзіледі, оның Meншiктi салмағы 1,001-1,007; ал pH 6,8-7,4 тең.
Ересек адамда тәулігіне 1-2 л сілекей түзіледі, оның Meншiктi салмағы 1,001-1,007; ал pH 6,8-7,4 тең.
Астың қорытылуына қатысады. Қатты тағамды шылауға, жібітуге, тағамды ұсақ бөлшектерге майдалауға қатысып, оның жұтылуын жеңілдетеді.
Ауыз қуысында сілекей альфа-амилазасының әсерінен
полисахаридтердің (гликоген, крахмал) корытылуы
басталады.
Қорғаныштық функция. Ауыз қуысының кілегейлі қабаты мен тістерді кeyiп кетуден, химиялық және физикалық факторлардың әсерінен, бактериялар мен олардың метаболиттік өнімдерінен қорғайды. Ticтi астың қалдықтарынан тазалайды. Сілекейдің құрамында қорғаныштық қызмет атқаратын лизоцим, пропердин, иммуноглобулиндер бар.
Ауыз қуысы тіндері мен сұйықтығының гомеостазын сақтауға қатысады
Tiс эмалінің минерализация және реминерализация үрдістеріне қатысады.
Сөйлеу қызметінқуаттайды
Экскреторлык функциясы - сілекейге заттар алмасуының соңғы өнімдері және дәрілік заттардың кейбір метаболиттік өнімдері түседі.
Тағам температурасын реттейді
128.Сілекейдің буферлік жүйелері. Ауыз қуысындағы қышқыл-негіздік тепе-теңдіктіңреттелуі
Сілекейде буферлік жүйелер (бикарбонаттық, фосфаттық, белоктық) бар. Бикарбонатты буферлік жүйе сілекейдегі бүкіл буферлік сыйымдылықтың 80%-н құрайды. Сілекей қышқылданғанда алымы(Н2С03) жоғарылап, бөлімі (NaHC03) төмендейді. Ары қарай бикарбонатты буферлік жүйе фосфаттымен әрекеттеседі.
Н2СО3 + Na2HPO4+ NaHCO3 + NaН2PO4
Белокты буфердің айтарлықтай маңызы жоқ.
129.Сілекейдің тәуліктік көлемі жәнефизикалық-химиялық параметрлері, олардың кариес кезіндегі өзгерістері. Қызылиек сұйықтығы, химиялық құрамы, биологиялық рөлі.
Ересек адамда тәулігіне 1-2 л сілекей түзіледі
Қызыл иек сұйықтығы қызыл иек жүлгесін толтырып, тicтi қоршап тұрған тіндер мен аралас сілекей арасындағы ортаны қалыптастырады. Қызыл иек сұйықтығы қан транссудаты мен сілекейдің қоспасы болып табылады. Қызыл иек сұйықтығының қалыпты жағдайдағы тәуліктік мөлшері 0,5-2,5 мл, рН-ы 7,9-8,3 тең. Қызыл иек сұйықтығында иммуноглобулиндер, ферменттер, лейкоциттер, лизоцим, цитокиндер, лактоферрин және қорғаныштық қызмет атқаратын басқа да заттар бар.
130.Аралас сілекейдің минералды компоненті, атқаратын қызметі "сілекей шунты". Аралассілекейдің Органикалық заттардың негізгі топтарының құрамы мен сипаттамасы
Астың қорытылуына қатысады. Ауыз қуысында сілекей альфа-амилазасының әсерінен полисахаридтердің (гликоген, крахмал) корытылуы басталады.
Қорғаныштық функция. Ауыз қуысының кілегейлі қабаты мен тістерді кeyiп кетуден, химиялық және физикалық факторлардың әсерінен, бактериялар мен олардың метаболиттік өнімдерінен қорғайды. Сілекейдің құрамында қорғаныштық қызмет атқаратын лизоцим, пропердин, иммуноглобулиндер бар.
Ауыз қуысы тіндері мен сұйықтығының гомеостазын сақтауға қатысады
Tiс эмалінің минерализация және реминерализация үрдістеріне қатысады.
Сөйлеу қызметінқуаттайды
Экскреторлык функциясы - сілекейге заттар алмасуының соңғы өнімдері және дәрілік заттардың кейбір метаболиттік өнімдері түседі.
Тағам температурасын реттейді
Минералды құрамы: Кальций 2,1-2,3 мМоль/л,,,,Калий 19-23 мМоль/л
Натрий 6,5-22 мМоль/л ,,,, Магний 0,4-0,9 мМоль/л
Фосфор 2,1-6,5 мМоль/л,,, Хлор 20-40 мМоль/л
Сілекей шунты- сілекей ас қорыту жолына түскесін, құрамындағы кальций, фосфат және басқа да төменмолекулярлы компонент иондары қанға сіңіріліп қайтадан қаннан сілекей құрамына өтетін жері.(кругооборот)
Органикалық заттары: Су-994г/л
Белоктар 0,8-4 г/л 90%(гликопротеидтер,ферменттер,А иммуноглобулин)
Муцин 0,9-6,0г/л, Холестерин 0,02-0,50г/л, Гистатин
Ферменттер: Альфа-амилаза; лизоцим; сілтілі және қышқылды фосфотаз; қан ұюына және фибринолизге қатысатын ферменттер (плазмин, плазминоген активаторы, фибриназа); нуклеаза; кининаза
131.Сілекей ақуыздары, олардың шығу тегі, патологиядағы өзгерістер. Сілекей және қызыл иек сұйықтығыныңферменттері: шығу тегі, құрылымы, әртүрлі аурулар кезіндегі белсенділігінің өзгеруі.
Қалыпты жағдайдағы мөлшері - 0,8-4 г/л.
Белоктардың 90%-ы (гликопротеидтер, ферменттер, А иммуноглобулині) сілекей бездерінде түзіледі, ал белоктардың 10%-ы қан сарысуынан, бактериялардан және ауыз қуысының кілегейлі қабықтарының жасушаларынан түседі. Қаннан альбуминдер, глобулиндер, липо және гликопротеидтер, G және М иммуноглобулиндер, трансферрин, церулоплазмин түседі.
Муцин – нағыз гликопротеид, оның құрамындағы амин қышқылдарының 1/3 бөлігін серии мен треонин құрайды. Бұл амин қышқылдарының гидроксил топтарына көмірсулардың туындылары (сиал қышқылдары, ацетилгалактозамин, галактоза, фрукоза) кіреді.
Гистатин - құрамында көп мөлшерде гистидинбар белок. Олгидроксиапатит кристаллдарының өcyiн тежейді, микробқа қарсы, вирусқа қарсы жәнесаңырауқұлаққа қарсы әсер көрсетеді, тістің кейін
пайда болған пелликуласының түзілуінеқатысады.
Сілекей цистатиндері.Бұл белоктар тістің кейін пайда болған пелликуласын қалыптастырады, цистеиндік протеиназалардың ингибиторы болғандықтан сілекей белоктарын протеолизден қорғайды.
Statherin-белоктар. Бұл белоктардың құрамында тирозин көп. Tic беткейіндегі фосфорлы-кальцийлі тұздардың преципитациясын ингибирлейді, яғни минерализация үрдістерін реттейді.
Сілекей ферменттері
Альфа-амилаза. Бұл сілекейдің негізгі ферменті полисахаридтердің олигосахаридтерге дейін гидролизденуіне қатысады. Әр түрлі патологиялық жағдайларда (инфекциялық гепатит, асқазан жарасы, гастрит, қантты диабет, эпидемиялық паротит және т.б.) оның активтілігі өзгереді.
Лизоцим. Бұл - мукополисахаридаза, бактериялық жасуша қабырғасыныңқұрамына кіретін a-N-ацетилмурамин қышқылын гидролиздейді. Бактерицидтікәсер көрсетеді,ауыз қуысындағы жарақаттың жазылуы мен тіннің регенерациясына септігінтигізеді.
Cілтілі және қышқыл фосфатазалар. Бұл ферменттер органикалық қосылыстардан бейорганикалық фосфатты бөліп алады. Қышқылдық фосфатазаны
сілекей бездері, бактериялар, лейкоциттер, эпителийжасушалары түзсе, ал сілітілі фосфатазаны (оптимальды pH 9,1-10,5) сілекей бездерінің түтікшелеріндегі эпителий жасушалары синтездейді. Ауыз қуысының жұмсақ тіндеріқабынғандажәне кариес кезіндеолардың активтілігіартады. Созылмалы гастритте,асқазан жарасында, пародонтитте олардың активтілігі төмендейді. Сілтілі фосфатаза бейорганикалық фосфаттың мөлшерін арттырып, эмальдің минерализациясын күшейтеді.
Қан ұюына және фибринолизге қатысатын ферменттер. Бұларға плазмин, плазминоген активаторы, фибриназа, фибринолиздің ингибиторы жатады.Олар фибриннің түзілуіне және ауыз қуысындағыжарақаттың регенерациясына қатысады.
Нуклеазалар. Вирустардың нуклеин қышқылдарын (ДНК, РНК,) ыдыратады.
Катеисиндер. Булар кышкылдык гидролазалар,сілекейге ауыз қуысының зақымданған тіндеріненжәне лейкоциттерден түседі.
Кининазалар. Сілекейде брадикининді ыдырататын кининазалар бар. Олардың шығу Teгi: сілекейбездері, микроорганизмдер, эпителий жасушалары.Пародонтоз кезінде ферменттердің активтілігі күрттөмендейді.
Протеииазалардын ингибиторы (трипсин ингибиторы). Созылмалытонзиллитте олардың мөлшері артады. Жануарлардың сілекей бездерінен алынған
протеиназалардың ингибиторын дәрілік препараттарретінде («трасилол», «контрикал», «гордокс») қолданады.
Сілекейде гликолиз ферменттері (альдолаза, гексокиназа), фосфоролиза, УКЦ ферменттері, липаза,аминотрансферазалар, ЛДГ, гликозаминогликандарды
ыдырататын ферменттер табылған.
132.Сілекей құрамының жас ерекшеліктері. Сілекейдің химиялық құрамына әсер ететін факторлар.
Сәби туғаннан бастап сілекей бездері жұмыс істейді, бірақ олардың өндіретін сілекейінің мөлшері алғашқы айларда өте аз болады. 3 айдан аса сілекейдің мөлшері көбейеді, бірақ бұл кезде баланың сілекейді жұту қабілеті нашар болғандықтан сілекейі шұбырып, өте көп өндірілетін сияқты сезім тудырады. Шын мәніндк оның айтарлықтай көбейетін мезгілі 9 бен 12 айда және 9-11 жаста. Мектепке дейінгі балалардың сілекейінің жалпы тәуліктік мөлшері 850-1000мл шамасында болады. Жаңа туған сәбидің шықшыт безінің салмағы 1,5-2г, жақ асты безі 0,72-1,00г, тіласты безі 0,2-0,6 г. Бұл бездер 3 айлық балада шамамен 2 есе, 6 айлықта 3 есе, 1 жаста 3-4 есе, 2 жаста 5 есе өседі. 5 жастағы балалардың сілекей бездерінің құрылысы ересектерге ұқсайды, бірақ олардың өсуі 13-15 жасқа дейін жалғасады.
Жаңа туған сәбидің сілекейіндегі амилаза мен лизоцим ферменттері жоқтың қасы, өйткені ана сүтінің құрамында бұл ферменттер бар. Амилаза ферментінің мөлшері 1 жаста ересек кісілердікіндей болады. Сәбидің сілекейінде аз шамада муцин, ал суы көп болады. Олар ауыз қуысының, тілдің құрғап қалмауын қамтамасыз етеді. Жаңа туған сәбидің сілекейінің реакциясы айтарлықтай тұрақты: әлсіз-сілтілі реакциядан сәл ғана қышқыл реакцияға дейін өзгереді, ал ересектерде күні бойы рН 5,6дан рН 8,3 ке дейін, яғни қышқылды реакциядан сілтілі реакцияға дейін өзгеріп отырады.
133.Диагностика үшін сілекей компоненттерін анықтаудың мәні. Пайда болған пелликуланың негізгі компоненттері,оның биологиялық рөлі
Муциндер, кальций, пептидтер, белоктар жұқа кілегейлі қабық - пелликула түзеді, ол эпителий жасушаларының мембраналарымен әрекеттеседі. Пелликула тосқауылдық және қорғаныштық қызмет атқарады, кілегейлі қабаттарды кeyiп кетуден сақтайды, өткізгіштігін реттейді.
134.Тістің құрылысы-эмаль, дентин, цемент
Tic тіндері
Tic оның қатты бөлігін құрайтын эмальдан, дентиннен, цементтен тұрады. Тістің қуысы бос дәнекер тiнi - ұлпамен (пульпа) толтырылған. Ticтiң түбipi дәнекер тінімен - периодонтпен қоршалып, альвеоларға бөлінеді.
Цемент, химиялыққұрамы, органикалық және бейорганикалық загтары.
Цемент тic түбipi мен мойнын қоршайды. Цементтің құрамында су шамамен 8%, органикалық заттар 25%және бейорганикалық заттар 68% құрайды. Бейорганикалық заттарының ішінде кальций фосфаты мен карбонаты көп. Органикалық заттарынан коллаген негізгі орынды алады. Цементтің eкi түpiн ажыратады: жасушасыз, біріншілік цемент (тic түбірінің мойын және ортаңғы бөлігінде орналасқан) және екіншілік - жасушалық (тic түбірінің жоғарғы жағында орналасқан). Жасушасыз цементте цементоциттер жоқ және ол коллаген талшықтары мен олардың арасына орналасқан аморфты жабыстырғыш заттан
тұрады. Жасушалық цемент құрамында көп мөлшерде цементоциттер болады.
Эмаль, химиялық құрамы, органикалық және бейорганикалық заттары.
Эмаль тіс сауытын жауып тұрады. Организмдегі ең тығыз тін, оның тығыздығы 2,9-3,05 г/см3 тең. Tic эмалі эпителийлік минерализацияланған тін. Ол механикалық және химиялық әсерлерге өте төзімді болып келеді. Эмальдың құрамында су шамамен 4%; бос су - 3,8% және апатит кристаллдарындағы гидраттық қабаты (3-3,3%). Органикалық заттары - 1,2%, оларға белоктар, липидтер, көмірсулар жатады.
Дентин, химиялық құрамы, органикалық және бейорганикалық заттары
Дентин тістің негізгі массасын құрайды. Дентин негізгі заттан және дентиннің трофикасын қамтамасыз ететінтүтікшелерден тұрады. Түтікшелерде одонтобласттың өсінділері, жүйке талшықтарының ұштары орналасады. Дентиннің сауыттық бөлігі эмальмен, ал түбip бөлігі цементпен қапталған. Дентиннің құрамында 72% бейорганикалық заттар, 21%-ға жуық органикалык заттар, шамамен10%) су болады. Бейорганикалық заттарының көп бөлігі кальций - 32%, бейорганикалық фосфор 16%-17%, сонымен қатар магний, натрий және т.б
микроэлементтер құрайды. Органикалық заттары дентиннің органикалық матриксін (сүйек тініне ұқсас) түзеді. Органикалық заттарына белоктарколлагендер, гликопротеидтер, ГАГтар жатады, онтогенез кезіндегі белок мөлшері тұракты 20-25%), дентинде ерекше белок фосфорин бар. Көмірсулардан энергия көзі ретінде гликоген бар. Липидтер жэне цитрат кездеседі.
135.Тістің пульпасы-бос дәнекер тіннің нұсқасы ретінде; пульпаның химиялық құрамы. Тістіңминералданған тіндері алмасуындағы пульпаның метаболикалық ерекшеліктері мен рөлі.
Ұлпа (пульпа), құрамы мен қызметтері туралы түсініктер
Ұлпа тістің сауытты қуысында және түбірлік каналдарда орналасады. Ол борпылдақ талшықты дәнекер тіннен тұрады, оның химиялық құрамы сүйек кемігіне ұқсас болып келеді. Tic ұлпасында қан тамырлары (васкуляризация) мен жүйке тамырларының көп таралуына байланысхы (иннервация) ол тамақтану мен тicтeгi заттар алмасуында белгілі орын алады. Heгізгi затының құрамына гликопротеидтер (хондроитинсульфаттар, гиалурон қышқылы, гексозаминдер) кіреді. Ұлпаның негізгі заты коректік заттардың қаннан ұлпаның жасушалық элементтеріне түceтiн, ал метаболизм өнімдері осы орта арқылы веноздық қан тамырлар жүйесіне түceтiн орха болып табылады. Ұлпа одондобласт Өсінділері арқылы дентин мен тic түбipiнің қоректенуін қамтамасыз етеді, ceбeбi тістің қатты тіндерінде қан тамырлары болмайды.
136.Тістің минералданған тіндеріндегі судың, органикалық және минералды заттардың арақатынасы.
137.Эмальдың минералды және органикалық компоненттерінің сипаттамасы. Тіс эмальының апатиттеріндегі ионалмасуының физика-химиялық сипаты; эмальдың өткізгіштігі, оның жетілуі. Эмальды минералдау процесініңөзіндік сипаты; эмальдің қалыптасу кезеңдері.
Эмаль, химиялық құрамы, органикалық және бейорганикалық заттары.
Эмаль тіс сауытын жауып тұрады. Организмдегі ең тығыз тін, оның тығыздығы 2,9-3,05 г/см3 тең. Tic эмалі эпителийлік минерализацияланған тін. Ол механикалық және химиялық әсерлерге өте төзімді болып келеді. Эмальдың құрамында су шамамен 4%; бос су - 3,8% және апатит кристаллдарындағы гидраттық қабаты (3-3,3%). Органикалық заттары - 1,2%, оларға белоктар, липидтер, көмірсулар жатады.Эмальдің белоктары қышқылдарда ерімейтін және
еритін болып бөлінеді. Эмальдің құрамына фосфопротеид типті белоктар - амилогениндер, энамелиндер, Е3 және Е4 фосфопротеиндер кіреді. Көмірсулары - негізінен гликоген және гликопротеидтердің көмірсулы бөліктері (хондроитинсульфат, галактоза, глюкоза, фруктоза, манноза, фукоза, ксилоза). Липидтерден негізінен фосфолипидтер. Цитрат (0,1%) кальцийдің еритін тұзын түзіп , кальцийді тасымалдау ролін атқарады. Осы тұздың құрамында сүйекке түскен кальций цитрат тотыққан кезде одан бөлініп кетіп, сүйекке сіңіріледі.Лимон қышқылының көмегімен кальций сүйектен қанға да тасымалданады. Эмальдің минералды негізін апатит кристаллдары құрайды. Гидроксилапатит кристаллдары (75%) цементке, дентинге және сүйекке қарағандаэмальде ірірек болады. Олар өте берік бір-бірінетығыз жанасқан, құрамында кальцийдің 12 атомы бар. Эмальде сонымен қатар бикарбонатапатит (19%) бар: бикарбонат анионы кристалдық тордағы ортофосфатпен немесе гидроксилмен алмасқан. Егер ол көп мөлшерде түзілсе, онда эмаль кариеске жеңіл ушырайды (көмірсулы тағамды көп пайдаланғанда). Шамамен 4,4% хлорапатит бар. Гидроксил тобын фторға алмастырғанда фторапатит түзіледі. Фторапатит кристалдық тордың
тығыздығын арттырып, эмальдің кариеске төзімділігін күшейтеді. Егер ауыз суының құрамында фтор көп болса, дақты эмаль, аз болса - кариес дамиды.
Стронций апатиттері өте осал болады. Стронций радионуклидінің жартылай ыдырау уақыты 20 жыл. Ол тағамға түссе, стронций рахиті (кашинбек ауруынемесе уров ауруы) дамиды. Эмальдің беткейіндегі катиондардың сапалық және сандық құрамы ауыз сұйықтытының құрамына тәуелді.
138.Дентин және дентин сұйықтығының химиялық құрамы. Плащ және интертубулярлы дентин минералдануы.Дентин түзілуінің өмірлік сипаты; екінші дентин. Дентин метаболизмінің ерекшеліктері.
Дентин, химиялық құрамы, органикалық және бейорганикалық заттары
Дентин тістің негізгі массасын құрайды. Дентин негізгі заттан және дентиннің трофикасын қамтамасыз ететінтүтікшелерден тұрады. Түтікшелерде одонтобласттың өсінділері, жүйке талшықтарының ұштары орналасады. Дентиннің сауыттық бөлігі эмальмен, ал түбip бөлігі цементпен қапталған. Дентиннің құрамында 72% бейорганикалық заттар, 21%-ға жуық органикалык заттар, шамамен 10%) су болады. Бейорганикалық заттарының көп бөлігі кальций - 32%, бейорганикалық фосфор 16%-17%, сонымен қатар магний, натрий және т.б
микроэлементтер құрайды. Органикалық заттары дентиннің органикалық матриксін (сүйек тініне ұқсас) түзеді. Органикалық заттарына белоктар коллагендер, гликопротеидтер, ГАГтар жатады, онтогенез кезіндегі белок мөлшері тұракты 20-25%), дентинде ерекше белок фосфорин бар. Көмірсулардан энергия көзі ретінде гликоген бар. Липидтер жэне цитрат кездеседі.
139.Цементтің химиялық құрамы; цемент пен сүйек тінінің арасындағы ұқсастықтар мен айырмашылықтар.Цементтегі зат алмасу. Периодонттың химиялық құрамы, метаболизм ерекшеліктері.
Цемент, химиялыққұрамы, органикалық және бейорганикалық загтары.
Цемент тic түбipi мен мойнын қоршайды. Цементтің құрамында су шамамен 8%, органикалық заттар 25%және бейорганикалық заттар 68% құрайды. Бейорганикалық заттарының ішінде кальций фосфаты мен карбонаты көп. Органикалық заттарынан коллаген негізгі орынды алады. Цементтің eкi түpiн ажыратады: жасушасыз, біріншілік цемент (тic түбірінің мойын және ортаңғы бөлігінде орналасқан) және екіншілік - жасушалық (тic түбірінің жоғарғы жағында орналасқан). Жасушасыз цементте цементоциттер жоқ және ол коллаген талшықтары мен олардың арасына орналасқан аморфты жабыстырғыш заттан
тұрады. Жасушалық цемент құрамында көп мөлшерде цементоциттер болады. Құрылысы мен құрамы жағынан жуан талшықты сүйекке ұқсас, бірақ Гаверс өзекшелері жоқ. Цементтің сүйек тінінен өзгешелігі-құрамында қантамырлары жоқ. Екіншілік цемент жасушалары пластикалық қызмет атқарады. Әртүрлі тітіркендіруші әсерлерден жаңа цемент қабаты құрылып, периодонт саңылауы тарылуы мүмкін. Бұл үрдіс «гиперцементоз» немесе цементтің шамадан тыс құрылуы деп аталады.
Периодонт-периодонт саңылауын (тіс ұяшығының компакті табақшасы мен тіс түбірі цементінің арасында орналасқан) толтырып тұрған тығыз дәнекер тін болып табылады. Периодонт құрамына байланыстырушы аппарат, жасушалар тобы, қан және лимфа тамырлары, нерв талшықтары, жасушааралық зат кіреді.Периодонттың қалыңдығы әр түрлі, адамның жасына, тұрған орныны, оған түсіп тұрған күшке байланысты болады. Астыңғы жақ сүйегінде периодонттың қалыңдығы: 0,15-0,22мм, үстіңгі жақ сүйегінде 0,7-0,8мм. Периодонт қанмен жақсы қамтамасыз етілген. Негізінен альвеола сүйегі арқылы қан тамырларының көмегімен таралады.
140.Кариес. Қышқылдың пайда болу теориясы. Тіс жегісіндегі биохимиялық өзгерістер.
Тісжегі –кариес- тістерді шыққаннан кейін жарақаттайтын, қатты тіндерінің минералды заттарының еруі, органикалық құрамының ыдырауы нәтижесінде тістің бүтіндігінің бұзылып, ақаулардың немесе қуыстардың пайда болуымен сипатталатын патологиялық үрдіс. Ауыз қуысында көптеген бактериялар кездеседі, бірақ тіс қағы пайда болуына және кіреукенің деминерализациялануына көбінесе қышқыл түзетін стрептококктар мен лактобактериялар қатысады. Тамақтанған соң бірнеше минутта рН-ң 6 дан 4 ке төмендеп кетуі байқалады. Тіс қағында көмірсу ыдырауынан пайда болған сүт қышқылынан басқа-құмырсқа, май, пропион және т.б. қышқылдар кездеседі.Эмальдің ең жиі кездесетін зақымдануы - кариес. Эмальдің ақауларының пайда болуына әкелетін басты үрдіс микроорганизмдер өндіретін қышқылдар мен лейкоциттерден түсетін иротеазалар мен тic қағындагы бос оттекті галогендік радикалдар. Қышқылдың әсерінен эмаль деминерализацияға ұшырап кариестің дамуына әкеледі.
141.Тістердегі беттік түзілімдер: кутикула, пелликула, тіс қабғы, тіс тасы, муцин пленкасы
Кіреуке бетінде қаратласа орналасқан өсінділер бар екені мәлім. Кутикула кіреуке ағзасының редуцияланған эпителиі болып табылады. Тіс шыққаннан кейін кутикула тамақ шайнау кезіндегі үйкеліс әсері нәтижесінде жойылады және кіреукенің беткейасты қабатында жартылай сақталады. Тістің қызмет атқаратын бет бұдан кейін сілекей әсерінен дамитын ақуыздыкөмірсулы кешеннен тұратын пелликуламен (жүре пайда болған кутикула) жабылады.
Пелликула кіреуке бетімен оның беткі қабатына ене тығыз байланысқан. Пелликула бетіне қалыптасқан тіс қағы,кіреуке бетіндегі жұмсақ шөгінділер келесі беттік өсінді болып келеді. Бұл субстанцияларды белгілеу үшін келесі «тіс тоғасы» «биоқабық» терминдерін қолданады.
Тіс қағының қалыптасуының маңызды кезеңі іртүрлі типтес микроорганизмдердің матрица бетіне шоғырлануы. Бұл микроорганизмдердің бір-бірімен және организммен қарым-қатынасы тіс қағында айқын микробтық гомеостазбен қамтамасыз етеді, ал бірақ тіс және пародонт тіні бүтіндігі бұзылмайды. Бұл тепе-теңдіктің сыртқы және ішкі қолайсыз факторлардың әсерінен бұзылуы патологиялық үрдістің дамуына, мысалы тісжегінің пайда болуына алып келеді. Тіс қағындағы әртүрлі микроорганизмдердің ішінде шынайы тісжегі туындатушы ретінде қышқылтүзеуші микроорганизмдердің маңыздылығын қарастыру керек.
Тіс қақтарының пайда болуы, құрамы, қызметі, қасиеті ауыз қуыс жағдайымен және организммен тығыз байланысты. Тіс қақтарының тісжегі тұындатушы потенциалы тамақта көмірсуларын көп қолданғанда, ауыз суында фтордың жетіспеушілігінде, ауыз қуыс гигиенасының нашар болуы кезінде жүзеге асады.
Тіс қағының құрамы және касиеті сілекеймен тығыз байланыста. Тістердің тіс жегіге бейімділігі немесе беріктілігі сілекейдің мына параметріне қарай анықталады: бөліну жылдамдығы, буферлік сыйымдылығы, сутекті иондардың концентрациясы (рН), органикалық және минералдық компоненттердің болуы. Тістердің сілекеймен шайылуы үрдісі кезінде тіс қағы және тіс тіндерінің клиренсі жүзеге асады. Тіс кіреукесі және сілекей арасында кальций және фосфат иондарының алмасуы жүруі нәтижесінде кіреуке қабаты бетінде, тіс қағында және сілекейде тепе-теңдік орнайды. Бұл сілекейдің кальций және фосфор ионымен қанығуына әсер етеді.
Тістерді тісжегіден қорғауда сілекейдің буферлік сыйымдылығы үлкен роль атқарады және қышқылдардың, сілтілердің бейтараптануын қамтамасыз етеді. Сілекейдің буферлік қабілетінің негізін карбонаттар, фосфаттар және ақуыздар құрайды.
Дата добавления: 2019-07-15; просмотров: 3210; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!