Факты, свидетельствующие о резкой неравномерности темпов «эволюции».
1.1. В первую очередь, это, конечно, знаменитый «кембрийский взрыв», когда в промежутке от 0 до 30 млн. лет[72] на Земле вдруг «решили» одновременно появиться все основные типы билатеральных животных (около двенадцати). То есть, подавляющее большинство высших таксонов современной фауны появилось именно путем «кембрийского взрыва». И с тех пор, вот уже на протяжении полумиллиарда (!) лет, структура высших таксонов почти не изменяется. Причем проблема не только в том, что новые типы больше не появлялись после кембрийского взрыва, а еще и в том, что они не появлялись и до кембрийского взрыва, на протяжении, как минимум, двух миллиардов лет. То есть, сначала было 2 млрд. лет относительного «эволюционного застоя»[73], потом (вдруг) за период от 0 до 30 млн. лет появляются почти все известные типы билатеральных животных. И после этого короткого бурного периода, опять наступает длительное «эволюционное затишье».
Понятно, что такая взрывообразная «эволюция» крупнейших биологических таксонов больше всего напоминает именно акт творения.
А вот чтобы объяснить этот поразительный феномен не актом творения, а результатом естественной эволюции - тут уже надо напрягать воображение. Действительно, трудно понять, почему именно в предполагаемом промежутке истории, да еще и в таком коротком (от 0 до 30 млн. лет) скорость «эволюции» вдруг подскочила в десятки (если не в сотни) раз, дружно породив целый букет основных типов животных, а потом упала обратно. Совершенно ясно, что для объяснения подобного чуда необходимо вводить дополнительные предположения о каких-то особых условиях, которые возникли именно в этот период, а потом куда-то «рассосались». По принципу:
|
|
|
- Карлсон был, но уже улетел (С)
То есть, чтобы объяснить кембрийский взрыв в рамках именно естественной эволюции, необходимо вводить те самые дополнительные сущности (об «особых условиях»), которые так не любил старик Оккам. Причем, что это за «особые условия» конкретно (ускоряющие воображаемую эволюцию сразу в десятки и сотни раз) пока остаётся предметом чисто теоретических рассуждений, не подтвержденных ничем более существенным. Другими словами, гаданием на кофейной гуще. В качестве возможных причин ускорения эволюции, например, называют: 1) повышение содержания кислорода в морской воде, 2) повышение содержания кальция (которое позволило организмам строить скелеты), 3) первое в истории Земли появление хищников, которое резко «подстегнуло» эволюцию (в том числе, на создание тех же скелетов и панцирей) и многие другие предположения.
Эти гадания на кофейной гуще серьезно осложняются тем, что кембрийские таксоны «вываливаются» в палеонтологическую летопись сразу «в готовом виде».
|
|
|
Во-первых, они «вываливаются» как бы из ниоткуда – практически, ни для одного высшего таксона, возникшего в раннем кембрии, нельзя уверенно назвать их докембрийских предков. А для некоторых кембрийских таксонов нельзя назвать даже предполагаемого предка. Потому что те, кто мог хотя бы претендовать на эту роль, в докембрийской палеонтологической летописи полностью отсутствуют.
Во-вторых, раннекембрийские животные «вваливаются» в палеонтологическую летопись с уже готовыми органами. И даже с целыми системами готовых органов. В том числе, самого сложного строения, которое мало чем отличается (или вообще не отличается) от строения этих же органов у современных высокоорганизованных животных, относящихся к этим же таксономическим группам.
Например, раннекембрийские животные «вваливаются» в палеонтологическую летопись с уже готовыми глазами совершенного строения.
До недавнего времени было известно, что некоторые группы древних членистоногих из вымершего класса трилобиты (Trilobita) имели глаза достаточно совершенного строения. Причем устройство глаз трилобитов в некоторых случаях отличалось оригинальностью (отличалось от глаз современных членистоногих).
|
|
|
Однако недавнее открытие отпечатков глаз животного из раннего кембрия (Lee et al., 2011) удивило биологов тем, что у этого «зверя» оказались типичные для современных членистоногих глаза такой степени совершенства, которая ничуть не хуже, чем у сегодня живущих высших членистоногих.[74] Глаза этого животного имели специфическую «зону яркости», и были более совершенны, чем глаза кембрийских трилобитов (Lee et al., 2011).
Чрезвычайно совершенными оказались глаза другого раннекембрийского членистоногого хищника, знаменитогоаномалокариса (Anomalocaris). Глаза этого животного были настолько совершенны, что по остроте зрения они конкурируют с глазами наиболее зорких современных членистоногих (Paterson et al., 2011):
…Их сохранившиеся визуальные поверхности состоят, по меньшей мере, из 16000 шестиугольно упакованных омматидиальных линз (в одном глазу), конкурируя с наиболее зоркими сложными глазами современных членистоногих.
Наконец, недавняя находка еще одного раннекембрийского животного (Cindarella eucalla) опять показала аналогичную картину – строение и степень развития глаз этого древнейшего членистоногого является вполне типичным для современных членистоногих (Zhao et al., 2013):
|
|
|
…Образец представляет собой старейшее микроанатомическое доказательство, подтверждающее возникновение высокоразвитого зрения в раннем кембрии, свыше 2000 омматидиев в каждом глазу.
Таким образом получается, что совершенные фасеточные глаза современного типа были весьма распространены уже у самых первых (из обнаруженных) членистоногих животных.
Более того, примерно то же самое относится и к глазам других типов. В частности, к камерным глазам позвоночных животных. Действительно, глаза были обнаружены уже у древнейшего найденного позвоночного животного Haikouichthys. Которое, в свою очередь… (правильно догадались!) было найдено тоже в раннем кембрии. Получается, что кембрийский взрыв породил сразу и одновременно глаза разных стилей дизайна - фасеточные глаза членистоногих и камерные глаза позвоночных (Zhao et al., 2013).
Точно так же обстоят дела и со сложной нервной системой и мозгом.
Как оказалось после сравнительно недавних ископаемых находок (Ma et al., 2012; Tanaka et al., 2013), строение и степень развития нервной системы и мозга (!) у древнейших животных периода кембрийского взрыва… тоже мало чем отличается от аналогичных органов высокоразвитых современных таксонов членистоногих животных (высших раков и хелицеровых). Причем в раннем кембрии в «готовом виде» появляются (опять) сразу оба основных типа мозга, характерных для современных хелицеровых и жвалоносных членистоногих (Tanaka et al., 2013).
Наконец, дальнейшее изучение одной из только что озвученных ископаемых окаменелостей привело к тому, что у этого раннекембрийского животного (Fuxianhuia protensa) обнаружилось уже и полностью «готовое» сердце, и весьма развитая система кровеносных сосудов (особенно, в районе головного мозга). То есть, обнаружилась вполне «готовая» сердечно-сосудистая система, соответствующая современным членистоногим (Ma et al., 2014).[75]
Понятно, что всё это «богатство» требует наличия сотен (если не тысяч) соответствующих генов, со сложнейшей системой регуляции их совместной работы. Непонятно, откуда такое «эволюционное богатство» вдруг взялось, да еще и сразу в таких объемах.
В общем, давайте повторим еще раз. Конечно, если биология скатится до уровня астрологии, предполагая, что в начале кембрийского периода все эти типы животных дружно возникли потому, что «так сложились звезды» (т.е. создались какие-то «особые условия»), тогда мы, конечно, сумеем «объяснить» кембрийский взрыв в рамках представлений о естественной эволюции. Вообще, предположения о «каких-то особых условиях» способны объяснить что угодно в рамках чего угодно (задним числом).
Но нельзя не признать, что больше всего кембрийский взрыв напоминает именно акт творения. Соответственно, этот факт укладывается в теорию непрерывного творения лучше.
1.2. Удивительно, но точно такая же ситуация складывается не только в отношении кембрийского взрыва… но и в отношении возникновения большей части всех биологических таксонов, которые когда-либо обнаруживались в палеонтологической летописи. Да, именно так. Большинство классов, отрядов, семейств, родов и даже видов (!) живых существ «вваливаются» в палеонтологическую летопись сразу «в готовом виде» будто бы из ниоткуда. То есть, демонстрируют тот же самый сценарий внезапного появления, что и новые типы животных кембрийского периода. Единственным отличием от кембрийского взрыва здесь является то, что в раннем кембрии вдруг дружно «вывалился из сингулярности»[76] сразу десяток типов животных. А в более поздние периоды времени биологические таксоны «вываливались» в палеонтологическую летопись уже не так массово, но так же внезапно.
Затем, после своего внезапного «выхода на сцену», биологические таксоны существуют какое-то время, практически не изменяясь. А потом снова исчезают из палеонтологической летописи (раньше или позже).
Понятно, что подобный сценарий «эволюции» биологического таксона просто «тютелька в тютельку» совпадает с тем сценарием, который можно было бы себе представить в рамках именно теории непрерывного творения.
Например, недавно была опубликована статья о самой древней находке членистоногих в янтаре (Schmidt et al., 2012). Это открытие «удревнило» подобные находки сразу на 100 млн. лет. В окаменевших каплях янтаря возрастом 230 млн. лет было обнаружено два вида древнейших (микроскопических) клещей.[77]
И вот эти «древнейшие из древних» клещей продемонстрировали… вполне современное строение тела. За что и были отнесены к современному надсемейству галловых клещей (Eriophyoidea). Примечательно, что это надсемейство является одной из самых специализированных групп в рамках всего таксона Acariformes.
Более того, один из найденных видов древнейших клещиков (Ampezzoa triassica) строением тела и характером восковых выделений вообще оказался очень похож на современный вид галлового клеща Cymeda zealandica (Schmidt et al., 2012).
Но и это еще не всё. Второй найденный вид галлового клеща (Triasacarus fedelei) морфологически серьезно отличается от первого. А это означает, что клещи обсуждаемой группы были достаточно разнообразны уже в триасовом периоде (Schmidt et al., 2012).
Что и говорить, весьма поучительная история в контексте нашего обсуждения – вот ученые находят самых древних представителей членистоногих в куске янтаря возрастом 230 млн. лет… А эти «древние представители» демонстрируют не только современную морфологию, но еще и принадлежат к одной из самых специализированных групп в рамках своего таксона.
Практически такая же история произошла еще и с находкой древнейших сенокосцев каменноугольного периода. Конкретно, было найдено два вида сенокосцев возрастом более 300 млн. лет, которые… тоже оказались обладателями поразительно современной морфологии (Garwood et al., 2011). За что и были отнесены к современным (ныне существующим) группам сенокосцев. Причем эти группы оказались еще и разными – это ныне существующие подотряды Dyspnoi и Eupnoi. А это означает, что сенокосцы групп Dyspnoi и Eupnoi были такими, какими они выглядят сейчас уже в каменноугольном периоде.
Еще пример. Давайте посмотрим вот на эту фотографию (Рис. 32):

Рисунок 32. Жук из современного семейства мягкотелок (Cantharidae) использует столь же современное «химическое оружие»… но 100 миллионов лет назад (Poinar et al., 2007).[78]
Здесь запечатлен жук из семейства мягкотелок (Cantharidae) как раз в тот момент, когда он защищался от какого-то нападавшего на него хищника. Жук защищался, выбрасывая из брюшка струю специального отпугивающего вещества. В этот момент его и застигла та самая роковая капля смолы, в которую он с тех пор погружен (Poinar et al., 2007).
Возраст этого янтаря около 100 млн. лет. Получается, что мы находим кусочек янтаря возрастом 100 млн. лет… и видим там уже вполне современное насекомое (из современного семейства), которое использует столь же современный способ химической защиты (используемый другими современными жуками из этого же семейства). Если я не ошибаюсь, это самая древняя находка подобного жука. Таким образом, мы имеем сразу (с первого же появления) представителя семейства жуков мягкотелок практически «в готовом виде», да еще и демонстрирующего нам «уже готовые» (полностью современные) способы химической защиты.
Это и есть то самое «выпадение из сингулярности» сразу в «готовом виде», о котором я говорил чуть выше. В данном случае напрашивается предположение, что этот жук был просто создан (тем или иным способом), а не постепенно эволюционировал через целую кучу переходных форм, которых в геологической летописи просто нет.
Именно такой сценарий внезапного появления новых таксонов является (еще раз) самым распространенным в палеонтологии. То есть, имеется множество разных биологических таксонов - не обязательно современных, но с вполне узнаваемыми (характерными) признаками и чертами, которые были свойственны этим группам организмов. Эти биологические группы внезапно появляются (в палеонтологической летописи), существуют какое-то время без существенных изменений (а то и вообще без всяких изменений), а потом вымирают (постепенно и незаметно, или же резко и массово).
Понятно, что эти (разные) линии организмов, если начать их сравнивать друг с другом, будут демонстрировать большую или меньшую степень близости. Это просто неизбежное следствие, которое вытекает из сравнения любых объектов с любыми объектами. Какие-то объекты неизбежно окажутся более похожими друг на друга, чем другие. Например, если мы начнем сравнивать автобусы, троллейбусы, трактора, легковые автомобили, мотоциклы и мотороллеры, то неизбежно придем к выводу, что морфологически, троллейбусы и автобусы ближе друг к другу (а мотоциклы и мотороллеры между собой), по сравнению, например, с тракторами. И вот на степени такого сходства, энтузиасты дарвиновской теории пытаются соединить похожие биологические таксоны пунктирными (воображаемыми) линиями эволюционного родства. По принципу:
- А от кого бы мог произойти этот биологический таксон, если бы дарвиновская эволюция была правдой?
Что из этого получается, я уже писал выше. Повторюсь еще раз. Такое «художественное пририсовывание» эволюционных предков и потомков – по степени спекулятивности ничуть не лучше, чем объединение вышеупомянутых мной куниц, кустарниковых собак и медведей в одну эволюционную цепочку (см. выше).[79]
Однажды, в качестве одного из свидетельств эволюции, мне привели следующие биологические группы из разных геологических эпох:
…Bunodidae и Pseudoniscidae (оба из силура) - сегменты опистосомы соединены свободно. Bellinuridae (средний девон - верхний карбон) - сегменты соединены плотно, но в единый щит ещё не срастаются. Paleolimulidae (карбон - пермь) - сегменты срастаются за исключением последнего, аксис опистосомы имеет отчётливую кольчатость. Limulidae (триас - ныне) - все сегменты опистосомы слиты, кольчатость аксиса видна слабо.
Здесь речь идет о разных группах мечехвостов (начиная с самых древних), которые последовательно сменяли друг друга в ископаемых отложениях. Очевидно, что в данном случае, при желании, можно провести воображаемые линии от предков к потомкам, причем в этой (воображаемо-эволюционной) цепочке будут прослеживаться определенные эволюционные тенденции - например, от свободного соединения сегментов до их полного слияния.
Но ведь точно так же можно провести «предково-потомковые» линии и в цепочке «куница - кустарниковая собака - медведь». И в этой цепочке тоже были бы ясно видны «эволюционные тенденции». Например, в сторону постепенного увеличения размеров тела. Или постепенного уменьшения длины хвоста…
Так переходили ли на самом деле Bunodidae в Bellinuridae (а после этого в Paleolimulidae, и затем в Limulidae)?
Или же это были просто разные группы мечехвостов, каждая из которых имела свои характерные черты. Причем эти характерные признаки были пронесены (каждой из этих групп мечехвостов) через всё время их существования?
Определенной подсказкой, которая могла бы помочь ответить на данный вопрос (конкретно в этом случае), возможно, является тот факт, что представители последней озвученной группы (Limulidae) – практически не изменились со времен триаса вплоть до наших дней (около 200 млн. лет). Поэтому современные мечехвосты являются одним из самых известных сегодня примеров «живых ископаемых».[80]
Возникает вопрос – а чего это мечехвосты, постепенно изменявшиеся (в нашем воображении) с силура по триас (примерно 200 млн. лет), после этого вдруг решили вообще перестать изменяться (тоже примерно 200 млн. лет)?
Может быть потому, что на самом деле, это была не постепенная эволюция (ранних групп мечехвостов в более поздние). А это были просто разные группы мечехвостов, каждая со своими характерными признаками. Которые существовали определенное время (сохраняя свои признаки), а потом исчезали.
Интересно, сегодня вообще есть кто-нибудь на Земле, кто сможет дать твердый ответ, как же здесь обстояло дело на самом деле? [81]
Только в некоторых, редких случаях палеонтология всё-таки находит ряды постепенных изменений организмов, более подробные, чем «куница-собака-медведь». В этих случаях появление биологических таксонов уже не кажется внезапным, а выглядит именно постепенным изменением одного таксона в другой. То есть, именно так, как это и должно было бы выглядеть, если бы дарвиновская эволюция была правдой.
Однако установленные случаи таких (подробных) рядов форм, постепенно переходящих друг в друга, являются весьма редкими. А сами изменяющиеся таксоны при этом весьма мелкие (обычно не выходят за пределы биологических родов). Поэтому возникает вопрос - а точно ли это эволюция? Или же здесь мы наблюдаем просто случайный «дрейф и блуждание» каких-нибудь легко изменяемых признаков внутри диапазона тех значений, которые были характерны для созданного (базового) таксона уже изначально? Если это так, то подобная «эволюция» должна заканчиваться примерно там же, где и начиналась.
Ну а в большинстве случаев мы наблюдаем эволюцию именно по сценарию внезапного появления новых таксонов. Причем чем крупнее ранг новых таксонов, тем «внезапней» их появление, и тем труднее подобрать им каких-нибудь «эволюционных предков», которые выглядели бы достаточно убедительно для того, чтобы против них не протестовала хотя бы некоторая часть специалистов по этому таксону.
Если какой-нибудь дарвинист воспротивится подобному сценарию эволюции, и начнет говорить, что про «внезапный характер появления новых таксонов» - Вы сами всё придумали, то в этом случае просто давите его авторитетом Стивена Гулда и Нильса Элдриджа. Эти палеонтологи в своё время как раз и подчеркнули именно «внезапный» характер эволюции. И даже предложили специальную «теорию прерывистого равновесия», которая была призвана объяснить, почему же новые биологические таксоны обычно появляются в палеонтологической летописи столь внезапно.[82] В рамках данной теории они постулировали, что виды изменяются не равномерно, а скачкообразно. То есть, долгое время биологический вид существует в «стационарном состоянии», почти не изменяясь. А вот изменяется биологический вид бурно – быстро эволюционирует за очень короткий (по геологическим меркам) период времени. Что и создаёт ощущение творения (внезапного появления) нового таксона. Из-за того, что за столь короткий период эволюционных изменений соответствующие переходные формы просто не успевают отложиться среди ископаемых останков в подавляющем большинстве случаев.
Поскольку научный авторитет Гулда и Элдриджа достаточно высок, я думаю, Вам без труда удастся задавить этим авторитетом дарвиниста, заставив его признать внезапный характер появления новых биологических таксонов в большинстве случае.
Понятно, что «теория прерывистого равновесия» и «теория непрерывного творения» являются, по сути, близнецами-братьями. Только теория непрерывного творения полагает, что появление нового таксона в палеонтологической летописи выглядит таким похожим на творение именно потому, что это событие и было творением на самом деле.
Итак (еще раз), столь распространенное в палеонтологии появление нового таксона по «внезапному сценарию» вписывается в концепцию непрерывного творения просто идеально.
С другой стороны, следует признать, что даже очень постепенная эволюция биологического таксона, тем не менее, тоже не будет противоречить теории непрерывного творения. Ведь мы не знаем характер той разумной силы, которая творила разные таксоны. Соответственно, не можем делать и предположения о том, с какой скоростью (и каким способом) эта сила должна была работать. Данное обстоятельство, в общем-то, не слишком красит нашу теорию непрерывного творения. Потому что делает её трудно проверяемой. Практически, эта теория является столь же «всеобъясняющей», как и дарвиновская теория эволюции, которая, как известно, может объяснить задним числом вообще что угодно.
1.3 Наконец, третью наблюдаемую неравномерность темпов эволюции можно художественно назвать «парадоксом слонов». В рамках дарвиновской теории, быстрее эволюционировать должны такие организмы, у которых, во-первых, быстрее сменяются поколения, а во-вторых, которые имеют популяции с большой численностью особей. Но палеонтологические факты показывают, что всё обстоит с точностью до наоборот. То есть, например, хоботные являлись (и являются) одними из самых больших и медленно размножающихся животных Земли. Тем не менее, они эволюционировали очень быстро – разные формы «слонов» сменяли друг друга в течение кайнозойской эры, как в калейдоскопе. А вот мелкие животные с огромной численностью популяций и высокой скоростью смены поколений, наоборот, эволюционировали медленно.
Пожалуй, лучше приведу прямую цитату профессионального палеонтолога (Расницын, 2002):
Согласно синтетической теории эволюционный процесс управляется в основном теми же факторами и механизмами, которыми оперирует генетика популяций. В этом случае и скорость макроэволюции должна подчиняться тем же закономерностям, что скорость изменения частот генов в генетике популяций, т.е. определяться мощностью потока адаптивно компетентных генетических вариаций, протекающих через эволюционирующую совокупность («Основное уравнение эволюции: скорость эволюции в пределах некоей популяции равна числу мутаций, возникающих в единицу времени, умноженному на долю фиксируемых мутаций»; Вилсон, 1985). Следовательно, скорость должна быть тем выше, чем выше частота мутаций и скорость смены поколений, чем больше размер популяций и мобилизационный резерв изменчивости.
Палеонтологический материал не позволяет прямо оценить большинство перечисленных параметров, используемых в генетике популяций. Некоторые из них, коррелирующие с другими признаками (размеры, таксономическая принадлежность), все же могут быть косвенно оценены у ископаемых. В частности, если генетический подход к эволюции справедлив, то ее скорость у млекопитающих, обладающих сравнительно медленной сменой поколений и небольшими популяциями (в связи с относительно крупными размерами особи), должна быть минимальной, особенно у наиболее крупных из них, у одноклеточных - максимальной, а у беспозвоночных - в среднем промежуточной. В действительности же, как показывают многочисленные палеонтологические данные, эти соотношения имеют обратный характер (Расницын, 1987). При этом различия весьма велики даже на уровне вида, таксономической категории, которая считается наиболее сравнимой в разных группах. Так, возраст (в миллионах лет) полусовременной фауны (в которой половина видов современные, половина - вымершие) у крупных млекопитающих (хоботные и копытные) 0.2, у мелких 0.5, у птиц и рыб 0.7, у насекомых 3-7, у моллюсков 3.5-5, у диатомовых водорослей - 15.
Понятно, что озвученные факты находятся в очень серьезном диссонансе с ожиданиями дарвиновской теории эволюции.
И столь же понятно, что в теорию непрерывного творения эти же факты вписываются лучше. Потому что теория непрерывного творения вовсе не требует, чтобы более крупные животные изменялись медленнее мелких.
2. Факты, свидетельствующие о несводимости происхождения биологических таксонов к какой-либо одной группе эволюционных предков.
Это очень важные факты, которые, с одной стороны, вступают в серьезное противоречие с дарвиновской теорией эволюции, но с другой стороны, полностью ожидаемы в рамках концепции непрерывного творения. То есть, эта группа фактов серьезно склоняет чашу весов в пользу именно теории непрерывного творения.
Сюда относятся все те многочисленные филогенетические конфликты, о которых я уже много говорил выше. Действительно, дарвиновская теория эволюции требует, чтобы гены в линии организмов-потомков были унаследованы именно от организмов-предков, а не от какого-нибудь «постороннего дяди» (т.е. от совершенно независимого таксона). Это чуть ли не обязательное требование дарвиновской теории. Потому что в рамках этой теории, у конкретной эволюционной линии организмов может быть только какой-то один эволюционный предок, а не много.
Поэтому обнаружение многочисленных фактов, когда организмы одной эволюционной линии имеют гены, сходные с генами организмов из совершенно другой эволюционной линии (включая самые неожиданные и далекие) – сразу же ставит дарвиновскую теорию в ступор. И заставляет объявлять каждый такой факт очередным филогенетическим конфликтом.
Поскольку таких фактов обнаружено уже много (см. выше)[83], и с каждым годом обнаруживается всё больше и больше, то биологи, конечно, пытаются как-то объяснять филогенетические конфликты в рамках своего любимого учения. Как известно, существует три варианта подобных «объяснений». Это: 1) «генетическая конвергенция», 2) «горизонтальный перенос генов», и 3) «нам требуются будущие исследования».
Однако подобные объяснения являются, как минимум, весьма спекулятивными, а в некоторых особо «тяжких» случаях выглядят уже как откровенные попытки спасти принятую теорию от разгрома любой ценой, даже с помощью самых невероятных допущений.
Действительно, для того чтобы объяснить, например, установленное множественное сходство между генетическим механизмом эхолокации летучих мышей и дельфинов (!) через некую «генетическую конвергенцию»… Для этого нужно как следует изнасиловать теорию вероятностей.
Но я уже достаточно говорил на эту тему выше, поэтому повторяться не буду.
А вот в рамках теории непрерывного творения, все подобные факты отнюдь не являются никакими «конфликтами». Наоборот, такие факты в свете этой теории настолько ожидаемы, насколько вообще можно чего-нибудь ожидать в рамках теоретических рассуждений. Действительно, если биологические таксоны создавала некая разумная сила, то этой разумной силе ничто не мешало использовать «технические разработки», созданные в рамках одного биологического таксона – для какого-нибудь другого таксона. Особенно, если требовалось решать примерно одинаковые задачи. Например, почему бы не использовать сходные механизмы высшей нервной деятельности в двух независимых, но при этом, высокоразвитых таксонах животных - членистоногих и позвоночных (см. выше). Или, почему бы не использовать сходный генетический механизм, обеспечивающий эхолокацию, в тех группах животных, которых было решено наделить этой способностью? Даже если эти организмы совершенно не похожи друг на друга во всех остальных отношениях (летучие мыши и дельфины). И так далее.
Кстати, помните, в главе, посвященной быстрому накоплению молекулярно-генетических фактов, не вписывающихся в дарвиновскую теорию, я написал следующий абзац:[84]
В общем, мы сейчас, похоже, наблюдаем крах аргумента про «генетику, уже доказавшую эволюцию», прямо в режиме реального времени. Если такие милые сюрпризы с массированной «генетической конвергенцией» продолжат стабильно обнаруживаться и в других молекулярно-генетических исследованиях, то разнообразным «народным просветителям», посвятившим долгие годы проповеди дарвинизма среди населения с помощью именно подобных «фактов генетического родства», наверное, придётся просто застрелиться от стыда.
Похоже, эти мои слова оказались в прямом смысле слова, пророческими. Массированная «генетическая конвергенция» действительно продолжает стабильно обнаруживаться. Причем этот процесс пошел так быстро, что я, честно говоря, уже устал добавлять такие факты в эту рукопись. Поэтому официально заявляю – то, что я сейчас озвучу – мои последние добавления на эту тему.
Итак, из самого последнего. Обнаружено, что генетические механизмы образования электрических органов у разных видов электрических рыб… правильно (!) опять cходные. Я даже не буду описывать этот пример своими словами. Просто приведу несколько характерных цитат из соответствующей научно-популярной заметки (Наймарк, 2014):
Цитата №1:
Бесспорно установлено, что электроциты получились из мышечных клеток, которые увеличились в размерах, упорядочились по взаимному расположению и иннервации и избавились от необходимости сокращаться, утеряв ту или иную часть белкового сократительного аппарата. Однако у разных рыб электрические органы настолько разные, настолько различаются по особенностям размещения на теле и строению клеток, по специфике иннервации, что об их общем предке и речи нет.
И тем не менее, цитата №2:
Оказалось, что в электрических органах на первый план вышли несколько определенных генов, сходных у всех исследованных видов; для этих генов единообразно изменился уровень экспрессии (повысился или понизился). Действительно, удивляет, что при резких различиях и самих электрических органов, и электроцитов всё же нашлись общие изменения, контролируемые общими генами — и те, и другие в достаточном числе. Эти гены вовлечены во все те метаболические пути, которые обслуживают параллельно возникшие свойства: суммацию возбуждения, дипольную сущность электроцитов, их крупные размеры и потерю сократительной способности.
И это несмотря на то, что, во-первых, электрические органы формировались независимо, а во-вторых, между некоторыми видами электрических рыб, которые исследовались в этой работе, согласно современным представлениям – не менее 200 млн. лет независимой эволюции, а между другими – 100 млн. лет эволюции (Gallant et al., 2014).
Цитата №3:
Получается, что конвергентно возникшие сложные органы формировались за счет изменений в регуляции одних и тех же генов. Видимо, для построения сложной схемы, такой, как электрический орган, природа пользуется одними и теми же инструментами, в данном случае — генами.
Какая же всё-таки разумная у нас матушка природа! Прямо как мудрый генетик, занимающийся генной модификацией организмов. Наверное, пора начинать писать еще и слово «Природа» с большой буквы, как личное имя.[85]
В общем, с точки зрения дарвиновской теории эволюции, мы опять наблюдаем очередной «удивительный случай». Который в рамках теории непрерывного творения совершенно не удивителен, а наоборот, прямо предсказывается этой теорией – независимое применение сходных «технических» (генетических) решений для создания функционально сходных органов.
И последний недавний пример. Выше я уже говорил о том, что генетические, анатомические и поведенческие особенности, обеспечивающие пение птиц и человеческую речь, оказались неожиданно сходными.[86] Но биологи продолжили исследования в этой области, и недавно выяснили, что даже на уровне экспрессии определенных генов – между разными группами певчих птиц имеется сходство. Несмотря на то, что согласно современным представлениям, способность к пению формировалась независимо в разных группах птиц. Более того, этот же набор специфических «певчих» генов, вовлеченный в организацию пения у птиц… опять оказался сходным с соответствующим набором генов, вовлеченных в организацию речи у людей (Pfenning et al., 2014). Причем экспрессия сходных генов обнаружилась в сходных отделах мозга (птиц и людей).
И это несмотря на то, что между птицами и людьми, согласно современным представлениям, никак не меньше 250 млн. лет совершенно независимой эволюции, от разных групп (очень древних) рептилий, которые явно не отличались речевым изобилием. Да что там говорить, даже у обезьян экспрессия соответствующих «генов речи» ничем не выделяется. А вот у людей и птиц – выделяется. То есть люди в этом отношении находятся генетически ближе к певчим птицам, чем к обезьянам.
В общем, в этом вопросе всё настолько ясно, что еще какие-нибудь комментарии, думаю, уже не требуются. Итак, многочисленные установленные факты использования одних и тех же «технических» (генетических) «наработок» в совершенно разных биологических таксонах – дают теории непрерывного творения решающее преимущество над теорией дарвиновской эволюции в полноте объяснений всего комплекса имеющихся биологических фактов.
Единственное, что еще следует отметить, так это то, что соответствующие (сходные) генетические комплексы практически всегда оказываются именно сходными, но не идентичными. Например, соответствующие гены – лишь сходны («гомологичны»), но не идентичны. Причем гены (часто) сходны тем меньше, чем дальше организмы находятся друг от друга на «эволюционном древе». Всё это можно интерпретировать точно так же, как это делается в рамках дарвиновской теории – что «непрерывное творение организмов» было растянуто на очень значительное (геологическое) время. Поэтому соответствующие гены уже успели измениться (под потоком нейтральных мутаций) настолько, насколько это было вообще возможно (чтобы возложенная на них функция еще могла выполняться).
Или можно предположить, что именно сходство, но не идентичность «технического решения» в разных биологических таксонах была заложена изначально. Подобно тому, как в автомобилях разных марок используются аналогичные, но не идентичные детали. Или сходные, но не идентичные технологии и технические решения. Такое предположение тоже вполне допустимо в рамках теории непрерывного творения. А вот в рамках дарвиновской теории эволюции, даже сходство (особенно множественное) генетических механизмов между очень удаленными таксонами – уже не имеет объяснений. Во всяком случае, выглядит очень странно.
3. Примерно так же обстоят дела и с биогеографическими фактами. Согласно дарвиновской теории эволюции, любой биологический таксон должен был произойти от какого-то общего предка. И поскольку этот «общий предок», в свою очередь, должен был обитать в каком-то конкретном регионе Земли, то и все линии его эволюционных потомков тоже должны быть приурочены примерно к этому же региону. Конечно, с поправками на «подвижность» этого биологического таксона, на прошедшее геологическое время, и еще на знаменитый «дрейф континентов», то есть на постепенное перемещение материков (за сотни миллионов лет).
В целом, биологические таксоны, действительно, обычно приурочены к определенным географическим регионам. Что используется верующими дарвинистами в качестве одного из «доказательств эволюции». Однако при этом дарвинисты «забывают» упомянуть о многочисленных исключениях из этого правила. И таких «исключений» в биогеографии никак не меньше, чем озвученных выше «филогенетических конфликтов» в генетике. А скорее, даже намного больше. Только в биогеографии подобные факты предпочитают называть не «филогенетическими конфликтами», а «биогеографическими загадками».
Итак «биогеографическая загадка» - это такой факт, в котором какой-нибудь отдельный биологический таксон (или таксоны) географически оказываются совсем не там, где это ожидается в рамках прогнозов теории эволюции. Нередко такие противоречия можно снять с помощью знаменитой концепции дрейфа континентов. Но при этом забывается, что у самой концепции дрейфа континентов дела обстоят тоже неважно. Эта теория всё время критикуется целым рядом специалистов-геологов за то, что многие геологические факты никак не хотят укладываться в эту теорию (см., например, Жирнов, 2011; Красилов, 2009; Антонов, 2007 (а,б); Караулов, 2005; Фролов, 2004; Udintsev, 1996; Белоусов, 1973 и др.). Причем критика этой концепции идет настолько резкая, что доходит до характеристик «мыльный пузырь» (Красилов, 2009) или даже до «эпидемии в геологии» (Караулов, 2005).[87]
Но и без учета этих геологических «разборок», число биогеографических загадок всё равно продолжает впечатлять. То есть, даже если верить в блуждания материков по древним океанам, принимая все воображаемые сценарии этого дрейфа за чистую монету, то и в этом случае, всё равно остаются многочисленные биогеографические загадки. Их так много, что я специально сейчас не озвучиваю конкретные примеры. Думаю сделать это в отдельной книге, где посвящу подобным «загадкам» специальный раздел. Сейчас же просто скажу, что в некоторых случаях, если следовать биогеографическим фактам, континенты придётся отправить в (воображаемое) путешествие по древним океанам… в обратную сторону от той, которую предписывает нам теория дрейфа континентов. Или даже пустить эти континенты выписывать зигзаги по древним океанам. Особенно в этом деле потрясает Индия. Чтобы объяснить некоторые недавно открывшиеся биогеографические факты (и совместить их с более ранними данными), необходимо эту Индию отправить в кругосветное путешествие, с остановками: в Австралии, на Мадагаскаре, в Юго-Восточной Азии, в северной Европе (!) и в Центральной и Южной Америке.
Впрочем, давайте всё-таки озвучим пару конкретных примеров «биогеографических загадок», чтобы иметь более точное представление, о чем идет речь. Начнем с глобального примера.
Как известно, повышенная степень своеобразия флоры и фауны некоторых удаленных островов трактуется, как доказательство независимой эволюции этой флоры и фауны именно на этих удаленных островах. Например, флора Гавайских островов имеет много эндемичных (более нигде не встречающихся) видов и родов растений (из примерно 2500 видов растений, встречающихся на Гавайях, более 90% являются эндемиками). Поэтому флора Гавайских островов была выделена в отдельную флористическую область. То есть, в случае флоры Гавайских островов, у нас всё обстоит именно по сценарию, предсказываемому в рамках теории эволюции.
Но дело в том, что на Земле существует такая область, которая отличается никак не меньшим, а наоборот, гораздо большим флористическим разнообразием, чем флора Гавайских островов, хотя эта область… никогда не отрывалась от континентов. То есть, мы имеем ничем не отграниченный (от континента) массив суши, однако флора этого массива суши выглядит… как самый уединенный остров из всех возможных. Этот знаменитый массив суши – Капская область. Она очень маленькая по размеру, но является территорией, наиболее богатой видами растений в мире. По современным данным, здесь насчитывается более 9000 видов растений.
Цитируем биологический энциклопедический словарь 1986 года издания:[88]
Капское флористическое царство занимает южную оконечность Африки. Флора этого самого маленького флористического царства необычайно богата (ок. 7 тыс. видов), в ней насчитывают 7 эндемичных семейств, в т.ч. груббиевые (Grubbiaceae), роридуловые (Roridulaceae), бруниевые (Bruniaceae), и более 210 родов, в основном моно- или олиготипных. Большинство видов представлено ксерофильными и склерофильными кустарниками, реже невысокими деревьями. В сложной и не во всём ясной истории флоры Капского флористического царства интересны её связи с другими флорами Южного полушария, существующие с тех времён, когда Гондвана представляла собой единый материк или только начинала распадаться. Так, некоторые роды семейств рестиевых и протейных, развившиеся в австрало-азиатской части Гондваны, достигли Южной Африки прямой миграцией. Распределение же видов родов ( Cimonia) (1 вид в Южной Африке, 16 - в Новой Каледонии), (Bulbinella) (5 видов в Южной Африке, 6 - в Новой Зеландии), (Tetraria) (38 видов в Южной Африке, 4 - в Австралии, 1 - на острове Калимантан) трудно или даже невозможно объяснить только дрейфом континентов. Для флоры царства характерны также роды, общие с тропической африканской флорой (молочай, алоэ и др.) и флорой Голарктики (эрика - более 450 видов, рута, каркас, падуб, маслина и др.).
То есть, как говорится, «всё смешалось в доме Облонских» (С). Мы имеем в Капском царстве, во-первых, огромное богатство видов растений, резко выделяющее эту область из соседних территорий, во-вторых, очень высокую степень своеобразия (по разным оценкам, от 70 до 90% эндемиков, включая целые эндемичные семейства). И в-третьих, здесь произрастают отдельные виды и роды растений, характерные для частей света, чрезвычайно удаленных от южной Африки.
В общем, такой вот «остров чудес» прямо посреди… суши. При этом никакими особо суровыми географическими барьерами Капская область не отделена. В качестве чего-нибудь особенного, как-то выделяющего Капскую область, можно назвать только климат (более умеренного типа, чем вокруг). Но если считать, что подобные особенности климата способны порождать такое большое разнообразие видов, родов (и даже семейств) - тогда у нас чуть ли не в каждом уголке Земли, хоть сколько-нибудь отличающемся по климату от других закоуголков, должны наблюдаться сотни эндемичных видов и хотя бы несколько эндемичных родов. Поскольку таких уголков (с различающимся климатом) у нас на Земле сотни тысяч, то всё имеющееся разнообразие видов сосудистых растений, наверное, должно было бы перевалить миллионов за сто.
Кроме того, значительная часть своеобразных растений Капской области – это растения, приспособленные к засушливым условиям (разнообразные ксерофиты и суккуленты). В связи с этим, засушливый климат областей, окружающих Капскую область, кажется, не должен был сильно ограничивать распространение этих растений на окружающие территории в ходе их постепенной эволюции. Тем не менее, мы имеем то, что имеем.
Впрочем, для некоторых суккулентов, никакая биогеография – не указ. Давайте оценим, например, ареал распространения знаменитых листовых суккулентов семейства Aizoaceae, в простонародье именуемых «живыми камнями». Центр их разнообразия – именно Капская область (и разнообразие это весьма впечатляет, по разным оценкам от 1500 до 2500 видов), однако отдельные представители этого семейства (относящиеся даже к одному и тому же роду), каким-то образом очутились: в Австралии, на Тасмании, в Южной и Северной Америке. В общем, как говорится, «весь мир в кармане» (С).
А вот 70% (как минимум) других видов Капской области (из 9000) почему-то «тусуются» только в Капской области, и нигде больше. Такая вот изолированная «Вселенная растений». В связи с этим не очень понятно, почему некоторые роды местных растений так «расшвыряло» по всему белому свету, в то время как растения сразу нескольких (эндемичных) семейств боятся ступить хотя бы немного за пределы Капской области. Как будто эти растения окружают многие сотни километров соленой океанской воды.
И вот теперь, после того, как мы познакомились с целым «островом» флоры, который изолированно «эволюционировал» прямо посреди… суши, давайте теперь сравним этот феномен с другим флористическим царством.
Это так называемое субантарктическое флористическое царство. В отличие от крошечного и предельно компактно расположенного Капского царства, это субантарктическое царство, наоборот, «разметано» по всей южной части Мирового океана, занимая, в том числе, изолированные острова. Конкретно, в это флористическое царство включена флора: 1) Новой Зеландии (и прилегающих островов), 2) крайний юг Южной Америки, 3) несколько субантарктических островов.
Казалось бы, что может быть общего между столь удаленными территориями? Но оказывается, очень много (общего). Включая целый ряд общих родов и даже биологических видов растений. Например, есть такой (субантарктический) остров Кергелен, на котором произрастают примерно 30 видов растений. Этот остров весьма уединен. Ближайшая суша от него – это Антарктида (на расстоянии 2000 км), остров Реюньон в Индийском океане (на расстоянии 3400 км), и наконец, Австралия (на расстоянии 4800 км). Однако флора острова Кергелен не хочет иметь ничего общего с озвученными участками суши. Флора острова Кергелен почему-то желает «дружить»… с флорой Огненной Земли. Но Огненная Земля - это самый юг Южной Америки, расположенный почти на другой стороне Земли от острова Кергелен, на расстоянии около 8000 км (проложенных через океан и еще кусок Антарктиды). Тем не менее, дружба между флорой Кергелена и Огненной Земли весьма впечатляет. Из 29 видов сосудистых растений острова Кергелен, 17 видов (!) общие с Огненной Землёй (Тахтаджян, 1978:210). То есть, более половины всех видов.
Следующей по степени близости к флоре Кергелена является флора Новой Зеландии (тоже находящаяся на огромном расстоянии от Кергелена), с которой остров Кергелен имеет 11 общих видов цветковых растений (Тахтаджян, 1978:210).
Такие удивительные вещи сейчас пытаются объяснять (в рамках «единственно верного учения») некими прошлыми связями между этими участками суши. Например, связями через воображаемый южный суперматерик (Гондвану), который существовал в южном полушарии много миллионов лет назад.
Но дело в том, что согласно современным представлениям, в рамках теории «дрейфа континентов» (не забываем, что у самой этой теории – большие проблемы с геологическими фактами)… материк Гондвана начал расползаться (на отдельные куски, которые потом стали дрейфовать вверх по океану)… уже в начале мелового периода. И интересующая нас (будущая) Огненная Земля 150 млн. лет назадуже была достаточно отделена от других массивов суши «расползающейся» Гондваны. В связи с этим получается одно из трех. Либо эволюция на субантарктических островах практически «заморозилась» с тех самых пор (т.е. на все последующие 150 млн. лет). Либо более половины общих видов флоры Кергелена и Огненной Земли получились в результате каких-то чудесных событий. Или же большая часть суши южного полушария погрузилась на дно морское отнюдь не 100 млн. лет назад, а совсем недавно. Поэтому общая флора острова Кергелен и Огненной Земли просто еще не успела никуда сэволюционировать. То есть, это такой своеобразный «привет» от знаменитой младоземельной (потопной) концепции.
Интересно, что самые первые остатки цветковых растений появляются в палеонтологической летописи примерно 140 млн. лет назад. Поэтому как то странно думать, что в это же время на территории (будущего) острова Кергелен (и будущей Огненной Земли) произрастало уже более десятка видов современных цветковых растений, из разных современных семейств.
Очевидно, что здесь необходим сценарий более поздней «телепортации» растений - уже через океанские воды. Причем «телепортации» весьма дружной, так как этот подвиг должна была совершить, по крайней мере, половина видов флоры Кергелена.
Но и в этом случае, если, например, предположить, что флора побережья Антарктиды (тогда еще достаточно теплой) «телепортировалась», с одной стороны, на Огненную Землю, а с другой стороны, на древнее плато Кергелен (которое тогда еще возвышалось над уровнем моря) - то всё равно это «великое переселение» должно было закончиться не позднее 20 млн. лет назад. Потому что считается, что плато Кергелен окончательно затонуло примерно 20 млн. лет назад.[89] Отсюда следует, что флора острова Кергелен и флора Огненной Земли – просто перестали эволюционировать с тех самых пор и до настоящего времени.
То есть, нам все равно необходимо объяснять отсутствие эволюции у флоры, разделенной восемью тысячами километров, и хотя бы 20 миллионами лет времени. Причем делать это придётся в отношении более чем половины видов растений, общих между Кергеленом и Огненной Землей.
Мы должны предложить какие-то причины такой остановки эволюции. Что случилось с эволюцией флоры в районе субантарктических островов? Неужели у растений перестали происходить случайные мутации? Или экологические условия перестали изменяться? Но экологические условия, как раз таки, изменялись весьма сильно. Хотя бы потому, что раньше (будущая) Антарктида была покрыта буйной растительностью. А сейчас там ледовый щит толщиной 2 км.
Итак, что мы имеем в целом?
С одной стороны, мы имеем на одном маленьком участке суши (Капская область) такой мощный эволюционный «взрыв», что результаты этого «взрыва» приходится выделять в целое флористическое царство. Несмотря на то, что это царство сформировалось в условиях практического отсутствия какой-либо серьезной изоляции.
С другой стороны, мы имеем общие роды, и даже общие биологические виды между самыми уединенными островами, какие только можно себе представить на нашей Земле-матушке. Несмотря на то, что последние (воображаемые) связи между этими участками суши должны были оборваться уже много-много миллионов лет назад.
Что и говорить, весёленький контраст демонстрируют нам биогеографические факты.
Понятно, что для объяснения подобных парадоксов какие-то гипотезы предлагаются. Например, Капское царство в целом, предлагается считать «осколком» (остатком) некоей древней флоры, которая сохранилась только на этом пятачке Земли, и больше нигде. Но «осколком» чего (?) является эта флора, со столь удивительным разнообразием растений? Неужели райского сада?
И почему какой-нибудь другой «осколок древней флоры» (например, та же Огненная Земля, которая «дружит» аж с Кергеленом), не демонстрирует подобной степени дробления видов?
В рамках дарвиновской эволюции, такие контрасты трудно объяснимы (если вообще объяснимы). Потому что в рамках современной эволюционной теории, силы, приводившие к эволюции организмов в Капской области, совершенно идентичны эволюционным силам, действовавшим в области субантарктического царства. Эти силы давно известны (постулированы современной теорией эволюции): 1) случайные мутации, 2) дрейф генов, 3) эффект основателя и 4) естественный отбор.
Итак, какой из этих факторов был включен на 500%-ную мощность в Капском флористическом царстве, и одновременно был отключен (до 5%-ной мощности) в субантарктическом флористическом царстве? И самое главное, почему?
В рамках дарвиновской теории ответить на эти вопросы очень сложно. Впрочем, придумать задним числом хотя бы что-нибудь можно в любом случае. Поэтому речь здесь может идти не о «невозможности объяснения», а о «сложности объяснения» в сравнении с «легкостью объяснения» в рамках разных оцениваемых гипотез. Так вот, в рамках дарвиновской теории, феномен Капского флористического царства, «поделенный» на феномен субантарктического флористического царства – может быть объяснен только с помощью весьма длинного списка допущений, некоторые из которых выглядят, как откровенно притянутые за уши. С позиций теории непрерывного творения, в озвученной «биогеографической сумятице» разобраться тоже нелегко. Но всё же легче, чем под руководством «единственно верного учения». Например, Капское царство (с его резко повышенным биологическим разнообразием), действительно, похоже на остаток какого-то древнего «очага производства» биологических видов.
Кроме того, Капская область наглядно демонстрирует нам, насколько разнообразной могла быть (в принципе) флора и фауна Земли в некий исторический период в прошлом – десятки тысяч эндемичных видов на очень небольших географических площадях. Это, наверное, еще одна своеобразная «подсказка» для младоземельной концепции. Потому что одна из основных трудностей младоземельной концепции – суметь «уместить» все эти толпы видов растений и животных (как ископаемых, так и современных) в рамках единого периода времени на нашей маленькой планете. Так вот. Не знаю как животные, но 9000 видов растений Капской области наглядно демонстрируют, что компактно умещаться они умеют.
И наконец, загадка этого флористического царства в какой-то мере обесценивает «биогеографические доказательства эволюции» в целом. Например, почему бы в свете всего озвученного не предположить, что, допустим, флора Гавайских островов – это тоже остаток какой-нибудь древней флоры? А вовсе не результат эволюции в условиях островной изоляции? Например, в отношении значительной части уникальной флоры острова Новая Каледония – так и считается (что это реликты древних флор). Ну и как теперь отличить своеобразие жизни, получившееся в результате эволюции в условиях островной изоляции… от своеобразия жизни, сохранившегося в качестве «остатка райского сада»?
Можно привести и другие примеры необъяснимой остановки эволюции у некоторых биологических видов, эволюция которых почему-то «заморозилась» вообще намертво сразу на огромных территориях (Расницын, 2002):
Не менее показателен длительный (миллионы, десятки и возможно даже сотни миллионов лет) эволюционный стазис на видовом и родовом уровне, когда об обмене наследственной информацией не может быть и речи. Так, популяции четырех или пяти из тридцати австралийских видов бессяжковых насекомых (Protura) на видовом уровне неотличимы от популяций с далеких материков и островов — Калимантан, Япония, Южная Африка, Европа (Tuxen, 1967).
Бессяжковые нестойки к высыханию и не покидают почву, поэтому трансконтинентальные миграции для них практически исключены, и, объясняя их распространение, нам, очевидно, не избежать ссылок на дрейф континентов (тем более что для бессяжковых известен и классический «дрейфовый» ареал вида, охватывающий оба побережья Атлантического океана, у Delamarentulus tristani Silv., распространенного на Коста-Рике и в Западной Африке; Tuxen, 1963). Но из этого автоматически следует возраст вида у бессяжковых, оцениваемый многими десятками миллионов лет.
Следует отметить, что автор этой цитаты немного поскромничал, оценивая возможный возраст озвученных им биологических видов. На самом деле, с позиций пресловутой «теории дрейфа континентов», Австралия потеряла сухопутные связи с Африкой (и тем более, с будущей Европой и Японией) уже 150 миллионов лет назад.
То есть, если говорить прямо, присутствие сразу четырех или пяти видов бессяжковых насекомых там, где они сейчас присутствуют – в рамках дарвиновской теории эволюции объяснить невозможно. Только одна эта биогеографическая загадка (без каких-либо других столь же удивительных биогеографических чудес) – уже опровергает дарвиновскую теорию эволюции.
Еще один пример «биогеографической сумятицы». Как известно, в африканском озере Виктория сегодня плавает более 300 разных видов рыб, относящихся к семейству цихлид. Но дело в том, что согласно геологическим данным, всего примерно 15 тысяч лет назад на месте озера Виктория… располагалась обширная равнина, по которой ветер гнал зеленые «волны» трав, и скакали стада антилоп, буйволов и жирафов, убегая от гепардов, гиен и львов (Nagl et al., 2000; Verheyen et al., 2003). А это значит, что большинство имеющихся сейчас видов цихлид в озере Виктория каким-то образом успели возникнуть за последние 15 тысяч лет. Уже после того, как будущее озеро стало наполняться водой. Предполагается, что эти виды возникли из тех цихлид, которые существовали в окрестной речной сети (Nagl et al., 2000), или в окрестных озерах (Verheyen et al., 2003).
Конкретно, в работе (Nagl et al., 2000), на основании молекулярно-генетического анализа (разных видов цихлид озера Виктория и его окрестностей), был сделан вывод, что сегодняшние виды цихлид (которые сегодня плавают в озере Виктория), произошли не от единственного предка (как нередко считалось раньше), а от разных исходных линий (разных предковых популяций). Тем не менее, большинство видов цихлид озера Виктория, по заключению авторов работы (Nagl et al., 2000), всё-таки должно было образоваться именно «на месте». То есть, уже в самом озере Виктория. А это обстоятельство всё равно приводит нас к цифре – не более 15 тысяч лет для большинства видов цихлид этого озера.
В чуть более поздней (независимой) работе, проведенной на эту же тему, говорится уже не о 300, а о целых 500 видах цихлид озера Виктория (Verheyen et al., 2003). Авторы этой работы дополняют молекулярно-генетический материал, собранный в предыдущем исследовании (Nagl et al., 2000), собственным материалом по другим видам цихлид. И показывают, что основные генетические линии цихлид озера Виктория, возможно, возникли существенно раньше, чем 15000 лет назад - поскольку такие же линии цихлид присутствуют еще и в соседних африканских озерах, особенно, в озере Киву. Откалибровав (по своему) «молекулярно-генетические часы»,[90] авторы получили время разделения основных генетических линий цихлид – примерно 100 тысяч лет. Авторы предположили, что эти (уже разделившиеся) основные генетические линии цихлид сохранялись в озере Киву (и в некоторых других местных озерах). А потом (уже гораздо позже) эти линии проникли в озеро Виктория (которое вновь стало наполняться водой после своего высыхания примерно 15.000 лет назад).
В общем, авторы этого исследования дали основным генетическим линиям цихлид озера Виктория – не более 130 тысяч лет возраста (Verheyen et al., 2003).
Но дело в том, что сегодня в озере Киву обитает всего 15 видов цихлид. В то время как для озера Виктория сами авторы работы (Verheyen et al., 2003) называют цифру в 500 видов цихлид. В связи с этим становится трудно представить себе сценарий массового переселения рыб из озера Киву в озеро Виктория (всех 500 видов), которые (по этому сценарию) потом должны были еще и тотально вымереть в озере Киву (оставив только 15 современных видов). Гораздо проще предположить, что сотни видов цихлид озера Виктория образовались уже всё-таки на месте, в постепенно наполняющемся водой озере, возникнув из нескольких основных предковых линий, которые проникли в это озеро из речной сети, или из того же озера Киву. Во всяком случае, значительная часть сегодняшних 500 видов цихлид озера Виктория должна была эволюционировать на месте.
Но это значит, что нам всё-таки придётся признать фантастическую скорость образования новых видов конкретно в этом озере – сотни разных видов рыб всего за 15 тысяч лет.
Интересно, что авторы исследования (Verheyen et al., 2003) не нашли связи между морфологическим разнообразием цихлид с одной стороны, и генетическим «расстоянием» между этими же видами, с другой. Точно так же выяснилось, что степень генетических различий между разными видами никак не связана еще и с видовым богатством в рамках этих таксонов.[91] Так, оказалось, что сотни морфологически и экологически разнообразных видов цихлид озера Виктория – генетически различаются между собой меньше, чем некоторые из 15 видов цихлид озера Киву между собой. Или 6 видов цихлид озера Альберт между собой (Verheyen et al., 2003). Отсюда следует, что критерий генетического «расстояния» вообще мало информативен для оценки скорости видообразования. С одной стороны, малые генетические изменения могут привести к образованию многих биологических видов, хорошо различающихся как морфологически, так и экологически. А с другой стороны, виды могут оставаться неизменными (и не разделяться на новые виды), несмотря на значительные (накопленные?) генетические изменения.
Но не будем отвлекаться. Давайте запомним темпы видообразования рыб – сотни разных видов за 15 тысяч лет.[92]
А теперь посмотрим на привычную нам обыкновенную щуку (Esox lucius). Эта рыба имеет огромный ареал, простирающийся по всей северной части Евразии и еще по Северной Америке. На протяжении этого ареала разные популяции щуки существуют в весьма разных экологических условиях. Достаточно сказать, что южные границы щучьего ареала проходят (в Евразии) по югу Черного и Каспийского моря и по водоёмам пустынь Средней Азии, а на северных границах своего ареала щучьи популяции располагаются в водоёмах тундровой зоны (Рис. 33):
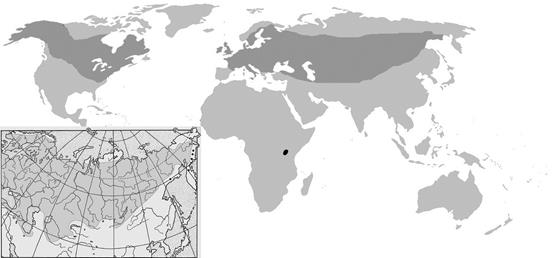
Рисунок 33. Общий (мировой) ареал обыкновенной щуки.[93] В нижнем правом углу – уточненный ареал обитания щуки на территории России и сопредельных территорий. Маленькое пятнышко черного цвета в Африке приведено для сравнения. Это то самое озеро Виктория, которым и ограничен ареал тех трех сотен видов цихлид, которые обитают в этом озере (пояснения в тексте).
Щуки живут и в опресненных частях морей, и в воде с кислой реакцией, и в стоячих озёрах, и в текучих реках (хотя предпочитают стоячие и слабопроточные воды) и т.п.
Но на всем этом ареале… щука представлена одним (!) видом.
То есть, в данном случае мы имеем нулевые темпы эволюции (по сравнению с эволюцией цихлид озера Виктория).
Как можно объяснить столь чудовищное различие в темпах видообразования в этих двух случаях? Ведь согласно дарвиновской теории эволюции, силы, приводящие к изменению биологических таксонов, в обоих случаях одинаковы. Это: 1) случайные мутации, 2) дрейф генов, 3) эффект основателя, 4) естественный отбор. Какие из этих факторов работали с бешеной скоростью в случае африканских цихлид, и фактически стояли на месте в случае щуки? И самое главное, почему?
Скорость появления случайных мутаций – считается примерно постоянной для любых биологических таксонов.
Дрейф генов - тоже не зависит от специфики озвученных случаев, а зависит лишь от размера популяций. Понятно, что размер щучьих популяций (в самых разных озерах, озерцах, реках, речках, речушках, старицах и болотах Евразии и Северной Америки) колеблется никак не меньше, чем размер популяций африканских цихлид в озере Виктория.
Естественный отбор – зависит от разнообразия экологических условий. Понятно, что разнообразие экологических условий, в которых вынуждены существовать разные популяции щуки (на всем огромном ареале),выше, чем у цихлид, дружно «тусующихся» в одном озере (пусть даже и очень большом).
Кстати, щука обитает и в озере Байкал, которое тоже является огромным, да к тому же еще и самым глубоководным озером мира. Тем не менее, никаких 300 видов щук мы в этом озере почему-то не наблюдаем.
Точно так же не наблюдаем мы здесь и 300 видов окуней. А наблюдаем мы на Байкале только один вид окуня. Это всем известный речной окунь (Perca fluviatilis), который распространен по всей Евразии так же широко, как и щука.
Но может быть, в столь чудесной скорости эволюции цихлид виноватпресловутый половой отбор? Может быть, цихлиды – это просто такие рыбы, которым для успешного выживания очень важны чисто эстетические переживания? И поэтому они особо придирчивы к внешности своих половых партнеров, что и заставляет эволюцию двигаться в сторону то одних, то других эстетических вкусов этих рыб?
Нет. Половой отбор не может ускорить темпы самой эволюции (как таковой). Потому что он не ускоряет появление случайных мутаций.
В первую очередь, половой отбор не может ускорить темпы изменения самих половых предпочтений. Ведь в рамках дарвинизма, эти изменения - тоже результат случайных мутаций. Но случайные мутации, приводящие к изменению самих половых предпочтений - не могут «бежать впереди паровоза», то есть, опережать скорость обычного мутационного процесса сразу на несколько порядков. А если всё-таки вообразить, что могут, тогда необходимо озвучить причины подобного галопирования мутаций.
То же самое касается и появления новых признаков. Даже такие признаки, которые предпочитаются в ходе полового отбора, не могут появиться быстрее соответствующих мутаций. Допустим, какая-нибудь молодая самка цихлиды мечтает о самце, отличающемся от всех других особенно красивым крутым лбом. Но как бы эта самка не мечтала о таком красавце-мужчине, её мечтам не суждено будет сбыться до тех пор, пока не возникнет соответствующая случайная мутация. А чтобы после этого самец стал еще красивее (т.е. еще более крутолобым) – должна случиться уже следующая случайная мутация. И так далее. Но соответствующие случайные мутации могут возникать только «в порядке очередности»[94], где очередь «движется» со средней скоростью… обычного мутационного процесса, примерно равного скорости мутационного процесса у тех же щук и окуней.
Наконец, половой отбор не ускоряет даже процесс закрепления любых признаков, крометех, которые прямо предпочитаются в ходе самого полового отбора (в данный момент времени).
Таким образом, единственное, что может сделать половой отбор в плане «ускорения эволюции» – это изолировать какую-то часть популяции рыб от другой части популяции этих же рыб, за счет соответствующей разницы в половых предпочтениях. А дальше можно предположить постепенное накопление различий между этими двумя (изолировавшимися) популяциями… в результате всё тех же: случайных мутаций, дрейфа генов и естественного отбора.
Однако одно озеро в лесотундре, отделенное от другого озера (где-нибудь в тайге) на расстояние в 200 км, наверное, изолирует одну популяцию щуки от другой популяции щуки ничуть не хуже (а скорее всего, гораздо лучше), чем пресловутый половой отбор. Следовательно, эволюция щук в этих изолированных озерах должна идти никак не меньшими темпами, чем эволюция африканских цихлид, воображаемо озабоченных половым отбором в пределах одного и того же озера.[95]
Давайте еще раз вспомним, что щука – это вид, весьма пластичный в экологическом отношении. Совсем маленькие щучки питаются зоопланктоном, потом переходят к более крупной «мелочи», а заканчивают весьма серьёзной добычей, доходящей вплоть до уток. Ну а про всяческое разнообразие: в скорости течения, pH среды, солености и жесткости воды, водного грунта, степени развития водной растительности – я уже говорил выше. Таким образом, казалось бы, щука очень легко могла бы «эволюционно расщепиться» сразу на целую кучу разных видов. Однако не «расщепляется».
Точно так же щука могла бы (теоретически) легко изменить размеры, окраску и пропорции своего тела. Действительно, мы видим, что цихлиды озера Виктория демонстрируют заметное разнообразие в пропорциях тела (или отдельных частей этого тела, например, головы, лба, плавников). Так почему же мы не видим соответствующей эволюции у щук? Почему нет разных видов щук с пропорциями тела, например, более близкими к судаку? Или к окуню? Или к ершу? Да хоть к знаменитой аквариумной рыбке скалярии? Ведь «луноликая» скалярия – это тоже хищник, и кстати, она тоже любит обитать среди зарослей водной растительности (точно так же, как и щука).
Ну так где же разнообразие видов щук? Почему «эволюция щук» и «эволюция цихлид озера Виктория» - настолько различны по результатам?
Дарвиновская теория эволюции не даёт ответа на этот вопрос. Верующим дарвинистам приходится выдумывать дополнительные сущности для объяснения этого парадокса. Например, часто используется такое «объяснение» – цихлиды, как биологический таксон в целом, исходно более экологически пластичны. И поэтому лучше эволюционируют, чем, например, щука. Или, допустим, какой-нибудь чир.[96]
Или тот же окунь. Как известно, ареал обитания речного окуня (Perca fluviatilis) тоже огромен. Разнообразие экологических условий, в которых обитает этот вид - тоже вряд ли меньше, чем у щуки. А по рациону питания окунь даже более пластичен, чем щука. Тут и зоопланктон, и разнообразные донные беспозвоночные (начиная с самой мелочи и кончая речными раками), крупные водные насекомые и их личинки, и конечно, еще и хищничество в отношении разных рыб (и даже лягушек).
Таким образом, все вопросы, которые мы уже задали по поводу отсутствия эволюции у щук, можно с точностью повторить и в отношении окуня.
Понятно, что несмотря на высокую экологическую пластичность обоих видов, у щуки и окуня всё же имеются определенные экологические пределы, которые эти виды уже не переступают. Например, слишком низкие, или наоборот, слишком высокие температуры воды. Интересно, что же мешает окуню или щуке эволюционировать еще и в этом направлении (тоже), породив целую кучу разных близких видов по примеру цихлид озера Виктория?
А ведь помимо щуки и окуня, в самых разных водоемах Земли обитает еще целая кучавидов рыб, которые не демонстрируют вообще никакой эволюции (за 15.000 лет), несмотря на: 1) громадные ареалы своего распространения, 2) с разнообразными экологическими условиями и 3) полной изоляцией между разными популяциями.
Чтобы далеко не ходить, можно отметить близкого родственника речного окуня – обыкновенного ерша (Gymnocephalus cernuus). Который распространен повсеместно на громадной территории. От Британских островов на западе, до бассейна реки Колымы на востоке. И от водоемов Скандинавии и бассейнов рек Северного Ледовитого океана (доходящих почти до самого Ледовитого океана) на севере, до Аральского моря на юге. Ерш обыкновенный - чрезвычайно экологически пластичный вид. Он живёт в стоячих озерах, текучих реках, опресненных заливах морей. Его можно найти как в пресных, так и в солоноватых водоёмах. На глубинах от 25 сантиметров до 85 метров. В равнинных и горных водоёмах. В водах, как бедных биогенными элементами, так и чрезвычайно ими насыщенных. Ерш переносит температуры от 0-2°С до 34.5 °C. Обычно держится в придонных горизонтах прибрежной зоны, или наоборот, на самой глубине, но может подниматься и в зону «открытой» воды. Очень пластичен в выборе корма. Его излюбленной пищей являются личинки комаров (хирономиды) или мелкие ракообразные (гаммариды), но ерш легко переключается и на другие виды корма, включающие любые другие формы донных организмов, зоопланктон или рыбную пищу, которая, в свою очередь, может состоять как из рыбной молоди, так и из рыбьей икры. С возрастом, если позволит размер, ерши могут становиться почти «чистыми» хищниками.
В связи со столь впечатляющим размахом экологической пластичности, казалось бы, мы должны ожидать эволюции, как минимум, десятков видов ершей чуть ли не в каждой новой речке или озере? Но нет. Ерш обыкновенный представлен единственным видом на всем своём громадном ареале. А всего в роду ершей (Gymnocephalus) сегодня насчитывается… 5 видов. Причем в отличие от обыкновенного ерша, остальные четыре вида занимают очень ограниченные ареалы, вплоть до единственного озера.[97] Как бы намекая нам – ну вот же она, начинающаяся «эволюция цихлид». Но нет. Эта «эволюция» заканчивается примерно там же, где и начинается – пятью видами ершей, среди которых ерш обыкновенный заполонил почти всю Евразию, при этом совершенно «позабыв» разделиться на соответствующее число видов. Ни за 15 тысяч лет. Ни даже за 50.
И в заключение нашего короткого обзора рыб, не желающих эволюционировать со скоростью цихлид озера Виктория, давайте упомянем еще одну рыбку. Весьма впечатляющий ареал демонстрирует трехиглая колюшка (Gasterosteus aculeatus). Эта небольшая рыбка обитает в самых разнообразных реках, озёрах и прибрежных морях Европы, Дальнего Востока, а также запада и востока Северной Америки. Такой вот кругосветный ареал. Про экологическую пластичность я уж и не говорю – трехиглая колюшка может жить и в море, и в пресной реке или озере, и в любых водоемах с промежуточной соленостью. Она может распадаться на несколько разных экологических форм (подобно речному окуню). Например, на форму, обитающую у дна, и форму, предпочитающую открытую толщу воды.[98] Тем не менее, этот вид, на всём своём громадном ареале, почему-то не разделился не то что на 300 разных видов (за 15.000 лет), но даже хотя бы на 10.
И это (еще раз) только отдельные факты, выдернутые из целого ряда полностью аналогичных примеров – видов рыб с большими ареалами и множеством прекрасно изолированных популяций, но, тем не менее, не эволюционирующих вообще никак (за 15.000 лет).
Да что там 15.000 лет? В природе есть абсолютный рекордсмен такого рода, который просто издевается над теорией эволюции. Это знаменитый щитень летний (Triops cancriformis)[99], который, как известно, полностью «забил» на какую-либо эволюцию, и существует вот уже… 230 миллионов лет (!) именно в качестве биологического вида Triops cancriformis… несмотря на то, что обитает в крайне изменчивых (эфемерных) местообитаниях - временных лужах (иногда в более долгоживущих водоемах). Да-да, именно так. Как бы невероятно это ни звучало, но панцири щитней и другие части их тел, которые были обнаружены в отложениях триасового периода - неотличимы от современных панцирей (и других частей тел) биологического вида щитень летний (Gore, 1986; Kelber, 1998).
Как известно, один из теоретиков эволюционной биологии, В. Грант, в своей монографии «Эволюционный процесс» написал, что виды, обитающие в непостоянных местообитаниях – просто не могут не изменяться со временем (Грант, 2008):
«…Брадителические, т.е. медленно эволюционирующие, животные встречаются в тех местообитаниях, которые оставались высокостабильными на протяжении геологической истории: в тропических лесах умеренно теплой зоны (опоссум), в больших тропических реках (крокодилы), в морях (Ostrea, Lingula, Litnulus) и в других столь же длительно существующих местообитаниях. И напротив, во временных озерах, на высоких горах, вулканических отложениях, в арктической тундре и тому подобных местообитаниях брадителические группы отсутствуют.
У популяции, обитающей в изменчивой среде, имеются три альтернативы: вымирание in situ, миграция или эволюция. В области, среда которой подвержена изменениям, брадителия невозможна. Для того чтобы оставаться в такой местности, популяция должна эволюционировать».
И правильно написал. Из постулатов современной теории эволюции этот вывод следует неизбежно. Действительно, не может оставаться эволюционно стабильным биологический вид, который: 1) регулярно попадает в разные экологические условия, 2) регулярно проходит через «бутылочные горлышки», и 3) регулярно образует новые популяции, в каких-нибудь удаленных лужах или чуть более долгоживущих водоемах.
Кроме того, следует вспомнить, сколько раз за эти 230 млн. лет экологические условия менялись еще и глобально. Моря отступали и наступали, климат «холодел» и «теплел». Даже континенты (предполагаемо) съезжались и разъезжались.
Однако щитень летний – это совершенно безграмотный товарищ. Он не читал Гранта, поэтому просто не знает, что обязан эволюционировать. Вот он и не эволюционирует вообще (уже 230 млн. лет).
Но вернемся обратно к удивительному объяснению верующих дарвинистов про «исходную пластичность» цихлид. Понятно, что какой-либо особой экологической пластичностью в сравнении со щуками, окунями, ершами или колюшками, цихлиды не обладают. Во всяком случае, эти способности не настолько выше, чтобы скорость их «эволюционного распада» на разные виды различалась на два порядка. Да и непонятно, каким образом эта (воображаемая) «повышенная экологическая пластичность» могла бы повлиять, например, на разнообразие пропорций тела? Неужели (допустим) растительноядность требует от рыбы какой-то особенной, конкретной формы тела, которая отличает эту рыбу от хищника? Нет, это не так. Иначе мы бы не имели нескольких тысяч разнообразных пропорций тела как у тех (растительноядных), так и у других (хищных).
В общем, из всего вышесказанного напрашивается предположение не о каких-то внешних (экологических) причинах, а о некоей внутренней особенности, свойственной таксону цихлид, которая и позволяет им «эволюционировать» гораздо легче и быстрее, чем другие таксоны.
Так может быть, где-нибудь в генах этого таксона просто заложены способности к образованию большого количества разных форм? То есть, здесь надо говорить об исходно запрограммированной способности цихлидобразовывать большое число видов. Но если это так, тогда здесь следует говорить не об эволюции, а о псевдо-эволюции, то есть, о запрограммированной вариативности внутри конкретного семейства цихловых (которая доходит до видового, а может быть, и до родового уровня).
Впрочем, этот же факт можно попытаться объяснить и в рамках естественной эволюции. Можно предположить, что соответствующие мутации у цихлид были всё-таки случайными. Но только происходили гораздо чаще на каком-то определенном участке ДНК.[100] Например, в районе, где находились гены, ответственные именно за окраску. И вот поэтому, мол, и получилось такое разнообразие окрасок цихлид озера Виктория.
Однако эти же цихлиды демонстрируют еще и разнообразие форм и пропорций тела. Хотя и не такое буйное, как разнообразие окрасок, оно всё же весьма впечатляет на фоне того «разнообразия», которое демонстрирует щука обыкновенная (по этому же поводу). Следовательно, нам придётся постулировать еще и другие «горячие участки ДНК» (помимо горячего участка по окраске), которые у цихлид были ответственны теперь уже за разные пропорции разных частей тела. Причем эти «горячие участки» снова (чисто случайно?) имелись только у цихлид, но почему-то напрочь отсутствовали у щук и окуней.
Не покажется ли нам (в конечном итоге), что такое теоретическое постулирование (ускоренной эволюции целой группы «горячих участков» в геноме цихлид, при одновременном отсутствии аналогичных участков в геномах щук и окуней) - выглядит несколько притянутым за уши? А на самом деле, неотличимо от той картины, которая наблюдалась бы в случае разумного дизайна этого биологического таксона? То есть, в случае сознательного внесения специальных генетических изменений (или особенностей) на строго определенных участках генома. В целях получения (в дальнейшем) автоматического биологического разнообразия по желаемому признаку.
То есть, не покажется ли нам, что щуки и окуни – эволюционно изменяться, видимо, не могут, потому что такая возможность в их геномах не запрограммирована. В то время как «эволюция цихлид» выглядит в точности так, будто она была специально запрограммирована в их геноме.
Причем цихлиды демонстрируют эту свою (загадочную) тягу к образованию очень большого разнообразия форм и видов одинаково замечательно как в Новом Свете (очень большое число видов), так и в Старом Свете (тоже очень большое число видов). Что является дополнительным подтверждением в пользу исходно заложенной в этом таксоне способности к подобным вещам. Эдакий «генетический конструктор рыб семейства цихловые», реализованный где-то внутри генофонда этого биологического семейства. Цихлиды вообще – одно из самых богатых видами семейств позвоночных животных.
Понятно, что гипотеза об исходно запрограммированной (внутренней) способности к образованию большого числа видов и форм - находится в русле теории непрерывного творения. Но никак не дарвинизма.
В рамках дарвиновской теории, никаких особенных преимуществ «в деле эволюции» цихлиды перед щуками иметь не должны (а также перед сигами, окунями, ершами и всеми прочими видами с огромными ареалами). Потому что в дарвиновской теории правят бал лишь: случайные мутации, дрейф генов и естественный отбор, отбирающий более приспособленных. А вот про «заранее запрограммированные способности» к образованию множества разных форм, в современном дарвинизме не упоминается.
Зато такая возможность чуть ли не предсказывается в рамках теории непрерывного творения. Где биологические таксоны эволюционируют либо потому, что их прямо изменяют, либо потому, что в новые (создаваемые) крупные таксоны заранее закладывается определенный потенциал для их будущего дробления на более мелкие таксоны. И те биологические таксоны, в которые был исходно заложен высокий потенциал будущей «псевдо-эволюции», соответственно, её и демонстрируют (цихлиды). А вот те таксоны, в которые этот потенциал не был исходно заложен (щуки, окуни и т.п.), соответственно, никакой «эволюции» не демонстрируют.
На всякий случай необходимо отметить теоретическую возможность еще одного, более радикального варианта, но тоже в рамках концепции непрерывного творения. Нельзя исключать, что и сам «распад» цихлид на разные роды или виды, хотя и был заранее предопределен (запрограммирован), но тоже происходил не произвольно, а направлялся соответствующей разумной силой. То есть, процесс формирования окончательного биологического разнообразия тоже мог корректироваться в той или иной мере. Если это действительно так, тогда можно говорить практически о «ручной лепке» (в переносном смысле, конечно) биологических видов в рамках разных таксономических групп.[101]
И в ходе такого (управляемого) процесса вышло так, что какие-то биологические группы получили большее число видов, а какие-то – меньшее. Или даже не «вышло так», а так и было запланировано, чтобы какие-то семейства рыб поражали разнообразием форм, расцветок и поведения. А другие таксоны создавались в более «скромном» варианте. В связи с этим интересно, что цихлиды – это, в подавляющем большинстве, тропические рыбы. А как известно, именно тропики поражают нас особенным изобилием разнообразных красок и форм, причем не только рыб, но и целого ряда других биологических таксонов (например, птиц, бабочек, цветов и пр.). Так что разнообразие цихлид может быть вполне запланированным элементом, вносящим свой (определенный) вклад в общий тропический стиль дизайна (буйства красок и форм).
Итак, феномен образования сотен видов цихлид за 15.000 лет в озере Виктория, «поделённый» на феномен не образования никаких видов щук, окуней, ершей и колюшек на огромной территории Евразии и Северной Америки, может быть объяснен одним из нижеперечисленных способов:
1. Кто-то очень увлекся таксоном цихлид, и наделал (практически «вручную») целую кучу самых разных видов этих очаровательных созданий.
2. В районе озера Виктория примерно 10 тысяч лет назад кто-то потерял работающий прибор «Видообразователь-ПГЦ3495».
3. Весь таксон цихлид был изначально создан с большим (но всё же ограниченным) запасом вариативности, которая теперь и реализуется этим таксоном рыб почти везде, где только эти рыбы способны прижиться (цихлиды, вероятно, исходно создавались, как тропические животные).
Очевидно, что все эти варианты находятся точно в русле теории непрерывного творения.
А теперь давайте проанализируем некоторые биологические факты, установленные как раз в той области, о которой мы сейчас ведем речь, и посмотрим, насколько эти факты согласуются с нашими вариантами теории разумного творения. Сосредоточимся сразу на третьем варианте. То есть, на варианте исходно созданного большого запаса изменчивости («на все случаи жизни»), который был «запрятан» (запрограммирован) где-то в глубинах созданного генофонда семейства цихловых.
Начнём нашу историю с того, что уже хорошо знакомый нам Александр Марков в 2006 году написал очередную заметку с говорящим названием «Для видообразования не нужны барьеры», содержание которой резюмировал следующим образом (Марков, 2006):
«Описан бесспорный случай возникновения нового вида рыб в маленьком озере в Никарагуа. Открытие немецких ученых фактически поставило точку в давнем споре о возможности симпатрического видообразования — разделения одного исходного вида на два без помощи физико-географических барьеров»
В этой заметке Александр Марков рассказал читателям о том, что в сравнительно небольшом озере Апойо (диаметр около 5 км), в Никарагуа (Центральная Америка), возник новый вид цихлид (Amphilophus zaliosus) путем отделения от другого, предкового вида Amphilophus citrinellus. Поcкольку озвученные виды цихлид разделились в пределах всего одного небольшого озера, то получается, что видообразование произошло здесь в отсутствие каких-либо географических барьеров. Такой путь образования новых видов на одной и той же территории называется симпатрическим. И поскольку Александр, видимо, симпатизирует этому пути видообразования, то он искренне порадовался по поводу того, что «описан бесспорный случай» и «истина торжествует».
А вот и наши «герои эволюционного фронта» собственной персоной (Рис. 34):
Рисунок 3 4. Вверху внешний вид двух обсуждаемых видов цихлид. Внизу их рацион питания. Взято из работы ( Barluenga et al., 2006). Пояснения в тексте.
Как видно из рисунка, оба вида цихлид хорошо различаются и морфологически, и экологически - их рационы питания заметно разные. Например, в меню A. citrinellus значительную долю составляет растительная пища, в то время как в меню A. zaliosus она практически отсутствует. Кроме того, A. zaliosus предпочитает находиться в толще воды, в то время как A. citrinellus тяготеет к дну водоема.
В общем, различий хватает.
Интересно, что озвученным видам цихлид для разделения тоже хватило всего примерно 10.000лет (Barluenga et al., 2006). Что весьма близко к обсуждаемым нами 15 тысячам лет эволюции цихлид озера Виктория. Само озеро Апойо образовалось тоже сравнительно недавно - около 20.000 лет назад, на месте жерла потухшего вулкана.
Понятно, что те верующие дарвинисты, которые почитывают научно-популярные заметки, тут же взяли на вооружение этот «явный пример видообразования» (то бишь, эволюции по их мнению). И сейчас используют «цихлид озера Апойо» в словесных баталиях с коварными креационистами едва ли не чаще, чем «эволюцию березовых пядениц», которая, видимо, уже успела надоесть за долгие годы её переписывания из одних литературных «доказательств эволюции» в другие.
Однако с 2006 года наука не стояла на месте. К настоящему времени биологи решили выделить в озере Апойо еще четыре (4) отдельных биологических вида, которых тоже отнесли всё к тому же комплексу Amphilophus s f. citrinellus. То есть, к комплексу видов, очень близких между собой, представляющих некие «вариации на тему» A. citrinellus. Поэтому сегодня можно сказать, что в озере Апойо плавают уже 6 видов из группы A. citrinellus. И таким образом, озеро Апойо в Никарагуа становится некоей «микрокопией» уже известного нам озера Виктория в Африке.
Более того, оказалось, что этот же вид цихлид (Amphilophus citrinellus) с великой готовностью «отпочковывает» от себя разные виды еще и в других никарагуанских озерах. Например, вид Amphilophus xiloaensis, относящийся к этому же комплексу видов (Amphilophus cf. сitrinellus), является уже эндемиком озера Xiloa. И так далее. В общей сложности, сегодня официально признано уже 11 разных видов из этого же комплекса видов. Причем этот список почти наверняка будет пополняться и дальше - некоторые специалисты говорят, что в этом видовом комплексе «скрыты» еще десятки видов, которые ждут своего описания.
Таким образом, что мы наблюдаем? Мы наблюдаем, что некий «крупный вид», то есть, собственно, Amphilophus citrinellus в его наиболее широком понимании, буквально «рассыпается» на разные (более «мелкие») виды, в любых изолированных водоемах, которые хоть сколько-нибудь подходят для его обитания. Складывается ощущение, что внутри его генофонда скрыты десятки разных (возможных) вариантов. И поэтому этот комплекс видов, переполненный буквально «рвущимися» из него «двойниками», как бы двоится, троится, и даже «десятерится» прямо у нас на глазах.
Но почему с Amphilophus citrinellus происходят такие странные вещи? Почему этот вид так надирает всё время «отпочковывать» от себя новые виды? В то время, как например, ерша или щуку – не надирает? Ведь тот же ерш распространен в тысячу раз большем числе водоемов (с самыми разными экологическими условиями), чем обсуждаемая нами цихлида. Тем не менее, ерш обыкновенный – остаётся ершом обыкновенным на всем протяжении своего гигантского ареала. А вот Amphilophus citrinellus «рассыпает» вокруг себя новые виды буквально во все стороны. Это как понимать?
Причем в данном случае, например, половой отбор – уже точно не «катит» в качестве «ускорителя эволюции». Потому что, как мы только что видели выше, разные виды этого комплекса различаются между собой экологическими признаками, которые половой отбор отбирать не может. Различаются эти виды и анатомическими признаками. Понятно, что самка этой рыбы не будет заглядывать самцу в глотку, чтобы оценить строение его глоточных зубов (глотки разных видов этого комплекса цихлид тоже различаются). В общем, к появлению и закреплению подобных признаков половой отбор не может иметь никакого отношения. Как я уже говорил выше, половой отбор (теоретически) мог бы только изолировать разные части популяций, за счет, например, предпочтений к определенной окраске. И дальше, уже изолированные, эти части популяций могли начать эволюционировать независимо друг от друга (то есть, накапливать независимые случайные мутации и подвергаться независимым факторам естественного отбора), постепенно превращаясь в разные виды.
Но как я уже говорил выше, ерш обыкновенный, живущий в водоемах Британских островов, изолирован от популяций ерша обыкновенного, живущего где-нибудь в низовьях реки Колымы – ну никак не меньше, чем цихлиды Amphilophus cf. сitrinellus, живущие в одном озере диаметром 5 км.
Так почему же (еще раз) цихлида Amphilophus сitrinellus буквально «рассыпает» вокруг себя новые виды, а ерш обыкновенный – и не думает этого делать?
Объяснений этому факту в рамках дарвиновской теории – нет.
А вот в рамках приведенного выше объяснения, в русле концепции непрерывного творения, цихлиды просто созданы с таким (исходно заданным) большим запасом изменчивости (т.е. скрытым набором разных вариантов фенотипов). Вот они их и «рассыпают» повсюду. Что в Никарагуа (в Центральной Америке), что в Южной Америке, что в Африке.
Напротив, в генофонде обыкновенной щуки скрытые фенотипы уже не «двоятся» и не «троятся». Поэтому она и ведет себя в этом отношении соответствующим образом – что в Северной Америке, что по всей Евразии - щука представлена одним видом.
Понятно, что и щука, и окунь, и всё тот же ерш - тоже демонстрируют определенную степень полиморфизма. Причем сразу по целому ряду признаков. Было бы очень странно, если бы морфологические признаки этих видов вообще никак не колебались на таких огромных ареалах, и в столь разных экологических условиях. Но всё же размах этого полиморфизма существенно скромнее того разнообразия форм, который демонстрируют нам цихлиды. То есть, генофонды и щуки, и окуня, и ерша тоже обладают определенным (заданным?) запасом внутривидовой изменчивости, но этот запас гораздо меньше, чем у цихлид. И в результате получается то, что получается – один биологический таксон сохраняется в рамках единого биологического вида на территории всего северного полушария, а другой биологический таксон образует сотни видов в одном озере.
Но и это еще не всё. Сегодня установлены и такие факты, которые могут помочь нам сделать еще более точные заключения о том механизме, который приводит к «рассыпанию» на разные виды комплекса Amphilophus cf. сitrinellus.
А именно в исследовании (Elmer et al., 2008) было установлено, что всё тот же Amphilophus cf. сitrinellus «отпочковал» от себя (снова) новую морфо-экологическую форму еще в одном никарагуанском озере - озере Apoyeque (тоже вулканического происхождения). Это совсем молодое озеро. Но еще моложе популяции населяющих его рыб. На основании собственных исследований, авторы пришли к выводу, что популяции рыб населяют это озеро не более 100 лет.
Тем не менее, этих 100 лет вполне хватило на то, чтобы Amphilophus сitrinellus «отпочковал» от себя новую форму, заметно отличающуюся от него как морфологически, так и экологически (Рис. 35):

Рисунок 35. Разные формы Amphilophus сitrinellus, образовавшиеся в озере Apoyeque за последние 100 лет. Взято из оригинального исследования ( Elmer et al ., 2008).
Как видно из рисунка, эти формы рыб неплохо различаются между собой морфологически. Особенно толщиной губ и вытянутой, более узкой головой (у «губастой») формы. Кроме того, имеются и некоторые другие морфологические отличия, которые менее заметны, но были установлены авторами данной работы. Например, некоторые различия в форме тела, форме челюстей, глоточных зубов и т.п.
Возможно, озвученные особенности «губастой» формы имеют определенное экологическое значение. Можно допустить, что «губастая» форма предпочитает собирать корм из щелей скальной породы. И толстые губы смягчают соприкосновение с острыми краями камней. Еще можно допустить, что толстые губы помогают справляться с теми животными, у которых имеется достаточно прочный (или колючий) панцирь. Например, с насекомыми. И действительно, в желудках вскрытых рыбок «губастой» формы обнаружилось существенно больше водных насекомых и их личинок, а также донных ракообразных. Ну а рыбы второй, «тонкогубой» формы, наверное, предпочитают не лазить по щелям в поисках корма. Основным источником пищи для них служат «белые водоросли» (Oomycota). Кроме того, чуть-чуть выше доля улиток и рыбы.
Можно, конечно, верить, что все перечисленные различия успели и появиться,и закрепиться всего за 100 лет, за счет возникновения случайных мутаций (которых, кстати, авторы данной работы практически не обнаружили, из чего и сделали вывод о крайне малом времени попадания этих рыб в это озеро), и еще в результате какого-нибудь давления естественного отбора (неизвестной силы). Однако если допустить подобную скорость эволюции, тогда у нас ерши должны были бы уже давно разделиться на миллион разных видов. По два-три вида в каждом мелком озере, и по сотне – в крупных. Потому что и появление случайных мутаций, и действие естественного отбора – в отношении ершей ничем не отличается от действия этих же эволюционных факторов на цихлид. Отличие только в том, что ерш обыкновенный – это более экологически пластичный вид, чем Amphilophus cf. сitrinellus, и следовательно (в рамках дарвинизма) имеет больше потенциальных возможностей для «дробления» на разные специализированные виды. Но тем не менее, почему-то не дробится. Несмотря на первоклассную географическую изоляцию.
А вот Amphilophus cf. сitrinellus, не успев еще как следует прижиться в озере Apoyeque, уже начал разделяться на две разные формы, приобретя всего за 100 лет целый список морфологических и экологических различий, при практически незаметных генетических различиях. А это значит, что генетические различия сосредоточились на крайне ограниченных участках ДНК, которые по этой причине можно, наверное, назвать просто генетическими переключателями,которые и «переключают» обсуждаемые фенотипы из одной формы в другую.
В общем, больше всего эти факты укладываются в рамки той самой «эволюции по волшебству», о которой я уже писал выше. Когда за крайне короткие периоды времени из какого-нибудь биологического вида вдруг «выпирает» целый комплекс новых черт, о существовании которых (у этого вида) никто даже не подозревал. Давайте еще раз вспомним вышеупомянутых жуков зигограмм, которые все дружно полетели всего за 4 поколения (см. выше).
Вот, похоже, и здесь мы тоже наблюдаем примерно такой же случай. То есть, ту самую скрытую изменчивость, о которой я уже столько говорил выше, и которой геномы цихлид, видимо, «набиты» просто до отказа. А вот геномы щук и ершей – уже не очень («набиты» скрытой изменчивостью).
И действительно, давайте посмотрим, что делают со всё той же цихлидой Amphilophus сitrinellus селекционеры-аквариумисты. Ведь эта цихлида – одна из популярных аквариумных рыбок. Дикую её форму Вы видели на рисунках выше. А вот она же, но уже после того, как побывала в «цепких лапах» селекционеров (Рис. 36):

Рисунок 36. Amphilophus сitrinellus, над которой «поработали» селекционеры-аквариумисты.
Причем красную окраску тела ей вовсе не селекционеры придумали. Формы с такой (ярко-красной) окраской регулярно встречаются прямо в природных популяциях рыб этого вида. А вот «крутой лоб» - это уже так постарались селекционеры. То есть, видимо, нащупали еще одно место скрытой изменчивости (этого вида), с которым тоже можно «побаловаться». Стоит ли теперь говорить, что «крутолобые» цихлиды встречаются достаточно часто и среди других представителей этого семейства? Или и так уже можно догадаться?
Это такой своеобразный «ремейк» знаменитых гомологических рядов Вавилова. То есть, демонстрация заранее заданной вариативности, исходно «прошитой» в геномах конкретных биологических таксонов.
И наконец, последнее (еще немного к вопросу о гомологических рядах). У нашей (уже хорошо знакомой нам) цихлиды Amphilophus сitrinellus есть еще один близкий родственник. Относящийся всё к тому же комплексу близких видов Amphilophus cf. сitrinellus. Это Amphilophus labiatus. Это тоже популярная аквариумная рыбка. И в природе обитает там же, где все перечисленные выше виды этого комплекса.
Amphilophus labiatus тоже весьма похожа на Amphilophus сitrinellus. Но отличается, в первую очередь, пухлыми губами. То есть, именно тем признаком, по которому у нас совсем недавно (чуть выше) пытались разделиться на две новые формы цихлиды озера Apoyeque.
Так вот, еще в одной сравнительно недавней научной работе (Colombo et al., 2013) биологи попытались понять причины многочисленных параллелизмов (схожестей), которые демонстрируют цихлиды водоемов Африки, и цихлиды американских водоемов. Биологи взяли для сравнения именно этот признак – толстые губы. И решили посмотреть, за счет чего же этот признак формируется у африканских цихлид, и за счет чего - у американских цихлид. В качестве биологических объектов для исследования из Америки авторы работы взяли наших знакомых «близнецов-братьев» Amphilophus citrinellus и A. labiatus. А из африканских цихлид они взяли тонкогубую Astatotilapia b urtoni и толстогубого Lobochilotes labiatus из озера Танганьика (Рис. 37):
Рисунок 37. Американские и африканские виды цихлид, исследовавшиеся в работе (Colombo et al., 2013). Рисунок взят из оригинальной работы.
И вот авторы работы посмотрели, какие конкретно гены (работающие в губах рыб), могут быть ответственны за формирование толстых губ (сравнив экспрессию 141 гена в губах толстогубых и тонкогубых видов рыб).
И оказалось, что за формирование толстых губ у африканских и у американских видов цихлид ответственен… один и тот же набор генов. То есть, толстые губы американского Amphilophus labiatus, и толстые губы африканского Lobochilotes labiatus формируются за счет примерно одинаковой работы одной и той же группы генов в количестве 6 штук. Из которых наиболее сильными кандидатами на основную роль в морфогенезе толстых губ оказались 3 конкретных гена (Actb, Cldn7 и Copb). Которые (правильно!) работают одинаково и у африканских, и у американских «губастиков» (Colombo et al., 2013).
Ну и кто бы сомневался? Действительно, в рамках представлений о заранее «прошитой» скрытой изменчивости в генофонде цихлид («генетическом конструкторе») по ряду определенных признаков (для которых в этом таксоне была заранее предусмотрена высокая возможная вариативность) - в рамках подобных представлений - только что озвученное открытие является настолько ожидаемым, что даже банально.
А вот в рамках дарвиновских представлений о развитии живой природы, такие генетические «параллелизмы» всегда вызывают ту или иную степень удивления. Ах, оказывается, какие бывают интересные совпадения! Ведь американские и африканские цихлиды эволюционируют независимо друг от друга вот уже 100 миллионов лет! Тем не менее, когда «эволюции» потребовалось сформировать толстые губы, на роль их «формирователей» были назначены… одни и те же гены. Хотя, если начать фантазировать, то способов, какими можно было бы, в принципе, защитить рот от повреждений панцирями беспозвоночных – можно было бы придумать десятки разных. Вот поэтому верующим дарвинистам и приходится писать, слегка недоумевая (Наймарк, 2012):
…По-видимому, для цихлид существует не так уж много способов сформировать конкретный признак, поэтому если возникает в нём необходимость, то в оборот берутся одни и те же гены.
Может быть и так. Но всё-таки это больше похоже на «генетический конструктор Лего», в котором одни и те же «повороты» конструктора приводят к одинаковым изменениям соответствующих признаков у разных видов.
Впрочем, для «одних и тех же генов» в количестве всего 3 штуки – простоесовпадение в ходе случайной эволюции вполне себе можно теоретически представить. Но когда речь идет уже о 200 (!) совпадениях в многочисленных генах, да еще и в совершенно неродственных биологических таксонах (как, например, в обсуждавшемся выше генетическом механизме эхолокации у летучих мышей и дельфинов)… вот тогда дарвинизм впадает уже в полный ступор.
Итак (еще раз). Подобные «генетические параллелизмы» в рамках теории разумного замысла – крайне ожидаемы и даже предсказываются этой теорией. А вот в рамках дарвинизма – это всегда неожиданность. Начиная от «недоумений легкой степени», и кончая полным ступором.
В общем, давайте больше не будем ходить вокруг да около, а честно скажем – теория непрерывного творения объясняет озвученные факты лучше современного дарвинизма.
При этом и многие другие факты тоже начинают пониматься лучше, стоит только взглянуть на природу в рамках озвученной нами концепции непрерывного творения. Например, сравнительно недавно была выполнена интересная работа по сравнению состава флоры тропических лесов в трех независимых регионах Земли, разделенных между собой многими тысячами километров, причем это расстояние еще и залито водами Мирового Океана (Рис. 38):
Рисунок 38. Регионы, в которых проводилось исследование ( Ricklefs & Renner , 2012). Красными кружками обозначены лесные массивы, подвергнутые анализу.[102]
А именно исследовалось видовое разнообразие тех современных семейств древесных растений, которые сегодня представлены во всех трех исследованных регионах (Ricklefs & Renner, 2012).
Согласно современным представлениям теории «дрейфа континентов», между тремя регионами простираются не только многие тысячи километров географического расстояния, но еще и многие миллионы лет времени. Например, соответствующие регионы Америки и Африки, по представлениям этой теории, «разъехались» друг от друга уже примерно 100 млн. лет назад. А отмеченные участки в Юго-Восточной Азии – вообще непонятно, когда имели последнюю связь с американскими и африканскими тропическими лесами. Во всяком случае, судя по представлениям теории «дрейфа континентов» – это было так давно (не меньше 200 млн. лет назад), что современных семейств цветковых растений в то время еще не было даже в «эскизном проекте» (Рис. 39):
Рисунок 39. Примерно так выглядела Земля 200 млн. лет назад по представлениям теории «дрейфа континентов». Америка и Африка (и еще будущая Австралия с будущей Антарктидой) еще «слиты в неразрывном единстве» в один громадный континент (Гондвану). А вот будущая Азия уже «плавает» весьма далеко от всего этого единства.
Поэтому не очень понятно, каким образом во всех трех озвученных регионах оказалось немало представителей общих семейств растений. И таких «биогеографических загадок» получилось немало – от 27 до 44 семейств оказались общими для обсуждаемых географических регионов.
Но мы пока сделаем вид, что как бы верим, будто эти семейства всё-таки так или иначе сумели осуществить своё практически кругосветное путешествие (столь дружным составом), и прижиться в трех удаленных точках Земного шара. И вот после этого, по представлениям дарвиновской теории, эти семейства деревьев начали свою независимую эволюцию (друг от друга) во всех 3 регионах. В качестве дополнительного свидетельства, что эта эволюция была независимой, озвучивается тот факт, что среди представителей этих общих семейств деревьев – почти нет общих родов. Следовательно, эволюция действительно протекала независимо.
А теперь давайте узнаем, чем же закончилась эта независимая эволюция, для десятков разных семейств растений в трех независимых географических регионах?
Результаты весьма впечатляют. Оказалось, что сегодняшнее разнообразие видов в исследованных семействах оказалось сходным во всех регионах. То есть, если какое-то семейство богато видами, например, в Америке, то оно будет богато видами и в Африке, и в Юго-Восточной Азии. И наоборот, те семейства, которые не могут похвастаться большим видовым разнообразием – показывают такие же скромные результаты и в других регионах. Причем установленная связь оказалась весьма серьезной. Соответствующие коэффициенты корреляции варьируют от 0.4 (умеренная сила связи) до 0.8 (высокая сила связи).[103] Причем эти коэффициенты корреляции почему-то оказались немного более высокими между тропической Америкой и Юго-Восточной Азией, чем между Америкой и Африкой (или между Африкой и Азией).[104] Несмотря на то, что согласно существующим представлениям, Америка и Африка разделились между собой гораздо позже, чем Азия, которая была сильно удалена от будущей Африки и Америки уже тогда, когда еще и самих то цветковых растений практически не существовало.
Даже обилие растений оказалось сходным в разных регионах. То есть, если растения (из того или иного семейства) обильно представлены в растительных сообществах одного региона, то представители этого же семейства с достаточно высокой вероятностью будут обильны и в растительных сообществах других исследованных регионов.
Как можно объяснить этот полученный результат?
Наиболее простым и очевидным объяснением в данном случае является такое:
- В каждое семейство был заложен определенный потенциал видового разнообразия (характерный для этого семейства), который эти семейства и реализовали.
И с этим обстоятельством даже случайные факторы мало что смогли сделать. Несмотря на то, что за прошедшие десятки миллионов лет случайностей разного рода должно было произойти огромное количество, тем не менее, видовое разнообразие трех десятков семейств древесных растений тропического леса реализовалось весьма сходным образом сразу в трех удаленных географических регионах Земли.
Понятно, что такой факт исключительно хорошо ложится в рамки представлений о создании биологических таксонов с заранее запланированным примерным объемом их будущего биологического разнообразия.
Второй вариант объяснения примерно такой же. Возможно, обсуждаемые семейства древесных растений были созданы с заранее запланированным объемом биологического разнообразия, но при этом и сам процесс последующего «распада» этих семейств на более мелкие таксоны – тоже происходил не произвольно, а контролировался (в большей или меньшей степени) определенной силой. В этом случае роль случайности становится еще меньше. В результате и получился сходный стиль дизайна тропического леса. Но одновременно этот дизайн отличается своеобразием на уровне биологических родов и видов. Возможно, для того, чтобы разные географические регионы Земли не выглядели в этом отношении «выполненными по копирку».
И наконец, третий вариант объяснения – самый спекулятивный. Но именно его и озвучили авторы данной работы. Они предположили, что исследованные ими семейства растений находились в тесной экологической связи друг с другом (в растительных сообществах) все эти многие десятки миллионов лет. Поэтому эволюция этих семейств и протекла сходным образом на всех трех независимых территориях. И привела к сходным результатам во всех трех случаях, хотя и шла совершенно независимо (тоже во всех трех случаях).
Понятно, что подобное объяснение является самым слабым из всех озвученных. Но поскольку только его и можно «впихнуть» в рамки естественной теории эволюции, то авторы озвучивают именно его. Честно говоря, у меня отказывает воображение, как такое «совпадение» могло стабильно выдерживать проверку временем (независимой эволюцией) на протяжении сотни миллионов лет. Особенно учитывая тот факт, что в эту (воображаемую) согласованную эволюцию десятков семейств растений неизбежно должны были вмешиваться тоже десятки эндемичных семейств тропических растений. То есть, таких семейств, которые тоже произрастают в этих же растительных сообществах, следовательно, тоже эволюционно формировались здесь же, но являются уникальными для каждого отдельного (озвученного здесь) тропического региона. Естественно, эндемичные растения, которые присутствовали (и присутствуют) в этих же растительных сообществах, тоже должны были вносить свой вклад в воображаемую согласованную эволюцию «трансконтинентальных» семейств, причем вклад этот был каждый раз уникальный (особенный) для каждого изолированного региона.
Поэтому пусть сами авторы объясняют сомневающимся читателям, как такое могло случиться, чтобы десятки «трансконтинентальных» семейств тропических деревьев в ходе независимой эволюции в течение сотни миллионов лет… в итоге заимели сходное число видов во всех исследованных регионах, на фоне постоянного вмешательства в эту историю тоже десятков эндемичных семейств (развивавшихся в этих же растительных сообществах).
А я лишь отмечу, что например, в отношении тех самых цихлид, о которых мы уже говорили выше, аналогичные соотношения не выдерживаются. Цихлиды имеют более 1000 видов в Африке, еще многие сотни видов в Америке… но никак не более десятка видов во всей тропической Юго-Восточной Азии. Вот тебе и «согласованная эволюция».
Итак (подытожим), самая простая версия, объясняющая аналогичность видового разнообразия в десятках «трансконтинентальных» семейств растений трех географически удаленных регионов Земли (Америке, Африке и Юго-Восточной Азии) – исходно заложенный в эти семейства определенный внутренний потенциал их видового разнообразия, который потом реализовался сходным образом в трех независимых случаях естественным (или управляемым) образом.
Дата добавления: 2018-09-20; просмотров: 442; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!