СКАЗКИ ДАРВИНИЗМА. СТАРЫЕ И НОВЫЕ
СТАРЫЕ ЛЕГЕНДЫ ДАРВИНИЗМА
Как я показал выше, со строгой доказательной базой у дарвинизма - серьезный «швах». Факты ни одной из озвученных выше областей биологии (сравнительной анатомии или эмбриологии, генетики или палеонтологии) не доказывают, что механизмы предполагаемой эволюции были именно дарвиновскими, а не какими-нибудь другими. Доказать реальность дарвиновской эволюции могут только примеры наблюдаемой эволюции, в которых было бы четко установлено, что именно случайные мутации + естественный отбор ответственны за появление тех или иных эволюционных изменений.
Но как мы уже с Вами убедились выше - таких строго установленных примеров (эволюции живых форм под действием естественного отбора) биологам пока зафиксировать не удалось. И это в природе, где с одной стороны, мы наблюдаем миллионы биологических видов, с другой стороны, предполагаем, что это фантастическое разнообразие жизни появилось именно благодаря «единственно верному учению» (случайные мутации+естественный отбор), и с третьей стороны, понимаем, что постулируемая нами эволюция должна продолжаться прямо сейчас. Однако с примерами этой «идущей эволюции» (именно под действием естественного отбора) почему-то совсем туго. Что уже само по себе весьма странно.
А казалось бы, что может быть проще? Сначала рассматриваем какое-нибудь животное. Например, жирафа (Рис. 19):

|
|
|
Рисунок 19. Всем известное животное – жираф (Giraffa camelopardalis).[45]
И видим у него очень длинную шею. Поскольку, согласно нашему единственно верному учению, все (ярко выраженные) признаки организмов - это результат действия естественного отбора более приспособленных, то отсюда следует, что длинная шея жирафа - это тоже итоговый результат выживания более приспособленных. Следовательно, длинная шея повышает приспособленность жирафа.
Теперь проводим специальное исследование этих жирафов. И выясняем, что разница в длине их шеи (между разными экземплярами жирафов), действительно, имеет не нулевую ценность для их выживания. А именно (допустим) длина шеи положительно сказывается на выживаемости жирафа.
И как только мы это выясняем в полевых исследованиях, после этого раскапываем пару десятков скелетов жирафов возрастом 100 тысяч лет. И демонстрируем миру, что длина шеи жирафов 100-тысячелетней давности была, в среднем, короче на 10 см, чем шея современных жирафов... Потом раскапываем еще пару-тройку скелетов жирафов возрастом уже 500 тысяч лет, и демонстрируем миру, что шея жирафа полумиллионолетнего возраста - была уже на 40 см короче, чем шея современного жирафа... И если теперь мы предъявим миру еще и скелет жирафа возрастом три миллиона лет, и покажем, что шея этого жирафа - короче шеи современного жирафа уже на целый метр... то всё, мы имеем очень серьезный пример «наблюдаемой эволюции под действием естественного отбора». Пусть и не самый строгий пример из всех возможных, но всё равно, весьма убедительный. Причем одновременно мы «затыкаем» еще и тех креационистов, которые возмущенно требуют предъявить им (печально знаменитые) «переходные формы». То есть, хотели переходные формы, товарищи? Вот, держите, скелеты жирафов с постепенно удлиняющимися шеями! И всё. Победа «единственно верного учения Дарвина» одержана. А посрамленные креационисты молча расползаются по темным уголкам, чтобы оплакать своё поражение...
|
|
|
Эту красивую сказку про жирафов дедушка Дарвин нам рассказал уже очень давно (еще 150 лет назад). Дедушка Дарвин рассказал нам, что жирафы получились такими удивительными созданиями, потому что из-за конкуренции с другими травоядными за пищу, жирафам необходимо было всё время вытягивать шею, чтобы добраться до еще нетронутой растительной пищи на деревьях. И естественный отбор постепенно отбирал тех жирафов, у которых шеи (или ноги) были немного длиннее, чем у их сородичей. Следовательно, эти жирафы могли дотягиваться туда, куда не дотягивались другие травоядные. В результате, эти жирафы лучше питались, как следствие, лучше выживали, оставляли больше потомства, и постепенно завоевывали популяции. А потом появлялись экземпляры жирафов с еще более длинными шеями, которые тоже использовали это преимущество... и так далее. И в конце концов, получился тот жираф, которого мы знаем сегодня.
|
|
|
Итак, что мешало проверить на протяжении 150 лет с тех пор, как нам эту эволюционную сказку рассказал Чарльз Дарвин, соответствует ли эта сказка реальному положению дел?
Да ничего не мешало. Поэтому пример с жирафами уже давным-давно должен был быть проверен, записан в строгие доказательства теории эволюции, и поставлен на все книжные полки для устрашения креационистов. Чтобы эти коварные враги хорошей науки даже и не думали подымать голову, придавленные столь мощным доказательством эволюции жирафов под действием естественного отбора...
Но что-то как-то не слышно о подобных громких победах дарвинизма.
Если говорить конкретно о жирафах, то никакие скелеты жирафов с постепенно удлиняющимися шеями - миру на обозрение почему-то не представляются. А те скудные свидетельства, которые мы имеем (например, следы жирафа в Лаетоли с предполагаемым возрастом 5 млн. лет) тоже помочь нам не могут. Во всяком случае, след этого 5-миллионолетнего жирафа ничем не отличается от следа современного жирафа (Sodera, 2009).
|
|
|
И самое главное, очевидно, что длина шеи жирафа - вообще не критична для его успешного выживания. Это очевидно просто потому, что например, самки жирафов могут быть гораздо ниже своих самцов (на 1-2 метра), но, тем не менее, почему-то не вымирают от голода. Да и вообще, самки жирафов совсем не стремятся питаться, вытягивая шею строго вверх. Они предпочитают поглощать пищу на средней высоте, держа шею почти горизонтально (на уровне плеч). То есть, реальные жирафы питаются совсем не так, как «завещал им питаться дедушка Дарвин».
И чтобы совсем уже распрощаться с этой легендой, озвучим результаты одного из решающих исследований на эту тему (Mitchell et al., 2010). В данном исследовании было установлено, что жирафы, погибшие во время засушливого сезона (то есть именно тогда, когда в саванне имеется ощутимый недостаток пищи) - имели более длинные шеи. Таким образом, суровая реальность (в очередной раз) не совпала со сказками дедушки Дарвина. Более того, эта реальность не просто не совпала, а оказалась противоположной этим сказкам.
Интересно также, что биологи как-то не слишком торопились проверятьсказки дедушки Дарвина. Первые попытки проверок начали всерьез осуществляться биологами только во второй половине прошлого века. И как только начали проверять, так напряжение и стало нарастать - проверки показывали с каждым разом всё яснее и яснее, что сказка, рассказанная дедушкой Дарвиным - чистая фантастика.
Например, выяснилось, что хотя длинная шея жирафа, действительно, является преимуществом в том случае, если cамые нижние листья деревьев (на уровне 1-2 метров) объедены «короткими» травоядными (Cameron & Du Toit, 2007), но:
1. Жирафы часто питаются на той же высоте, что и другие травоядные (Du Toit, 1990).
2. Предпочитают питаться на уровне собственной холки (Young & Isbell, 1991) большую часть времени (Leuthold & Leuthold, 1972; Pellew, 1984).
3. Даже в сухой сезон питаются в верхушках деревьев, только когда имеются новые побеги (Pellew, 1984).
4. Кроме того, питание на верхних уровнях имеет свою цену - жираф несколько теряет в бдительности (Cameron & Du Toit, 2005).
Впрочем, все эти подробности из биологии жирафов уже не кажутся существенными. Потому что в последнем исследовании, как я уже говорил, обсуждаемая гипотеза Дарвина изучалась самым прямым способом из всех возможных. Исследование поставило в этом вопросе жирную точку, установив, что преимущественно гибнут жирафы с самыми длинными шеями.
Хотя непредвзятым читателям и так было заранее ясно, что сказка, рассказанная дедушкой Дарвиным 150 лет назад, откровенно наивна. Действительно, жирафы могут питаться с высоты 4.5 метра (!) в то время как их предполагаемые конкуренты (другие виды копытных) - с высоты всего 2 метра. Уже из сравнения этих цифр становится совершенно ясно, что «сказка дедушки Дарвина» является сказкой в самом полном смысле этого слова. Действительно, чтобы уйти от конкуренции с другими видами, зачем жирафу надо было перерастать эти виды... на два с половиной метра? А самцы жирафов перерастают этот «порог ухода от конкуренции» уже на 3-4 метра. Неужели «естественный отбор жирафов» - это столь порывистый сказочный персонаж (со столь широкой душой), что разогнавшись, лишканул (с нужным признаком) эдак метра на три?

Рисунок 20. Наглядная разница между высотой жирафа и высотой «конкурирующих травоядных».

Рисунок 21. Еще одна наглядная разница между высотой жирафа и высотой «конкурирующих травоядных».
Правда, остаётся еще теоретическая возможность внутривидовой конкуренции между самими жирафами, как причины удлинения их шей. По принципу «гонки вооружений», где «важен каждый лишний дюйм или два» (по выражению самого Дарвина). Но похоже, эта версия выглядит в глазах полевых биологов (исследующих жирафов в реальной природе) настолько наивной, что они эту версию в своих публикациях не упоминают совсем. Действительно, если самки жирафов - почти на метр ниже самцов, то о каком «соревновании ввысь» между жирафами может идти речь? Ведь именно на самку ложится дополнительная нагрузка по вынашиванию плода (в том числе, и в период засухи) и последующее выкармливание жирафёнка молоком. Поэтому именно в отношении самок естественный отбор должен был сработать в наибольшей степени. И если бы жирафы действительно конкурировали друг с другом за еду, и если бы им действительно было выгодно стать выше «хотя бы на дюйм или два», тогда самки (как продукты естественного отбора) должны быть, по крайней мере, не ниже самцов. Однако мы видим противоположную картину. Что весьма недвусмысленно говорит нам о том, насколько, на самом деле, жирафы «нуждаются» в столь длинных шеях.
И действительно (еще раз). Как показали исследования 2010 года (Mitchell et al., 2010), в период засухи больше всего погибло жирафов с наиболее длинными шеями - высокие взрослые крупные самцы, которые (по предположению авторов работы) были просто не в состоянии обеспечить себя достаточным количеством пищи в этот период (54% всех смертей, см. рисунок ниже).
Из 15 обнаруженных скелетов взрослых жирафов, погибших во время засушливого сезона - 13 скелетов принадлежало самцам и только 2 скелета - самкам. В то же время (согласно данным, представленным в статье) популяции жирафов в дикой природе состоят на 25% из взрослых самцов, и на 31% из взрослых самок. Такое различие между половой структурой живых жирафов в популяции (среди взрослых особей) и числом погибших (в засуху) самцов (87%) относительно самок (13%) - однозначно указывает на то, что причина смертей в этой категории - отнюдь не какие-то посторонние факторы (например, преклонный возраст жирафов), а именно большой рост и масса тела, которые у самцов выше, чем у самок.
На всякий случай (для особо любопытных), в природе жирафы могут доживать до 26 лет, а в неволе (рекорд) вообще до 36 лет. Однако средняя продолжительность жизни жирафов в дикой природе - 6 лет, следовательно, взрослые жирафы в природе гибнут в основном не от старости.
Кроме того, в исследовании погибло некоторое число подростков (1-3-летнего возраста), которые еще не «переросли» другие виды копытных (антилоп куду и канна) - 32% всех смертей. Именно данная категория жирафов (жирафы-подростки) конкурировала со всеми остальными видами копытных. И всё равно смертность в этой категории в засушливый период (32%) оказалась гораздо ниже, чем смертность среди высоких взрослых животных (54%).
А выжили преимущественно самки и молодежь жирафов (см. рисунок 22, категория 2: от 3 до 4 метров высоты):
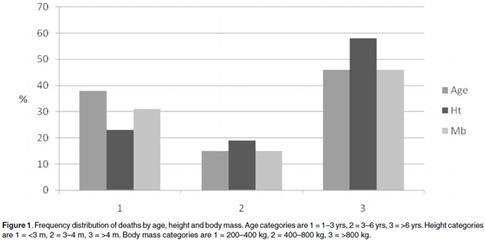
Рисунок 2 2 . Распределение смертности жирафов в период засухи (%) по возрасту, росту и массе тела. Категории по возрасту (серые столбики): 1 – возраст 1-3 года; 2 – возраст 3-6 лет; 3 – возраст более 6 лет. Категории по росту (темные столбики): 1 – рост менее 3 м; 2 – рост 3-4 метра; 3 – рост больше 4 м. Категории по массе тела (светло-серые столбики): 1 – масса тела 200-400 кг; 2 – от 400 до 800 кг; 3 – более 800 кг. Рисунок взят из работы ( Mitchell et al ., 2010).
Из рисунка получается, что самый оптимальный рост для жирафа 3-3.5 метра (в категории выше 4 метров смертность резко возрастает). В связи с этим совершенно непонятно, зачем жирафу надо было становиться ростом 5-6 метров?!

Рисунок 23. И снова «жертвы естественного отбора самых длинных шей» во всей красе.
Можно посчитать и по-другому. И тоже получится очень забавно. Оптимальный размер жирафа мы только что определили в 3-3.5 метра. Одновременно мы знаем, что жирафы легко достигают высоты 3 метра уже в холке! Отсюда получается, что жираф мог бы вообще не иметь длинной шеи, чтобы выживать в природе максимально успешно. Теперь понятно, почему самки предпочитают питаться практически на уровне своих плеч (то есть, держат шею во время питания практически горизонтально). И тем не менее, только одна шея жирафа зачем-то достигает двух метров длины!
Правда, можно предположить, что у жирафа, наоборот, слишком короткая шея. Ведь несмотря на то, что его шея достигает двухметровой длины, ноги жирафа тоже очень высокие. В результате, жираф, даже имея шею рекордной длины, имеет определенные проблемы, связанные с недостаточной длиной его шеи. Например, чтобы просто напиться воды на водопое жираф вынужден несколько «раскорячиться» (Рис. 24):

Рисунок 2 4 . А может быть, у жирафа, наоборот, слишком короткая шея? Потому что ноги слишком длинные и туловище слишком высокое?[46]
То есть, даже такая банальная процедура, как питье, вызывает у жирафа определенные трудности. Естественно, возникает вопрос – ну и зачем же жираф отрастил себе такие высокие ноги и туловище, что даже шея двухметровой длины оказывается для его тела слишком короткой? Особенно учитывая то обстоятельство, которое мы только что выяснили выше - что жираф мог бы прекраснейшим образом иметь рост вообще на 2 метра ниже (в целом).
И это далеко не единственная проблема, с которой вынуждены сталкиваться жирафы, имея столь специфический дизайн внешности. Соответствующие проблемы так часто перечислялись в разных источниках, что мне даже как-то неудобно повторять их еще раз. Но всё же:
1. Из-за огромной высоты тела возникают повышенные требования к эффективности работы кровеносной системы. Особенно это касается кровоснабжения мозга жирафа. Ведь мозг находится «где-то там в вышине» (на высоте 5-6 метров). Поэтому сердце жирафа очень большое, сильное и создаёт очень высокое кровяное давление.
2. Однако высокое кровяное давление, в свою очередь, порождает другую проблему. Если жираф опустит голову, из-за такого высокого давления он мог бы получить немедленное кровоизлияние в мозг. А если не получит, то после резкого поднятия головы, он, наоборот, потерял бы сознание от резкого падения давления крови, снабжающей мозг. Для того чтобы все эти неприятности с жирафом не случались, у него имеется целый комплекс соответствующих приспособлений. Во-первых, кровь жирафа гораздо более густая, чем у человека. Во-вторых, кровеносные сосуды имеют прочные стенки. В-третьих, в кровеносных сосудах имеются особые запирающие клапаны, которые прерывают поток крови, когда надо, и поэтому перепады давления не могут быть слишком сильными.
3. Очень высокий рост предрасполагает к скоплению жидкости в ногах. Кроме того, остановить кровотечение из раны, полученной в нижней части тела, становится проблематично. Однако жирафы имеют какую-то особенную кожу (и фасции), которая предохраняет их нижние конечности как от скопления жидкости и отеков, так и от чрезмерной кровопотери.
4. Особо длинная шея создаёт проблемы и для дыхания жирафа – трахея, проходящая через всю шею, слишком длинна и воздух, находящийся в ней, образует как бы «мертвую зону» (вдыхаемого, но не используемого воздуха). Однако легкие жирафа имеют особо большой размер, что позволяет успешно решать и эту проблему тоже.
5. Наконец, рождение любого жирафёнка, казалось бы, должно быть смертельно рискованным. Ведь роды у жирафов происходят стоя, и поэтому рождающийся жирафёнок должен упасть с высоты 1.5–2 метра, что при его весе и одновременно длинной и хрупкой шее было бы чревато переломом этой самой шеи. Однако и эта трудность тоже преодолена жирафами - за счет специальной позы новорожденного (и его выхода задними ногами вперед) ни особо рискованного падения, ни переломов шеи не происходит.
Всё это, конечно, очень здорово и весьма впечатляет. Действительно, создаётся ощущение качественно проделанной работы, которая была призвана решить (и решила) многочисленные проблемы, порождаемые специфическим дизайном жирафа. С продумыванием даже тех мелочей, которые сходу и в голову то не придут. Однако опять возникает вопрос – а зачем жирафу и его удивительный дизайн, и сопутствующие этому дизайну проблемы (и успешное решение этих проблем), если выяснилось, что лучше выживают такие особи жирафов, которые имеют рост на 1, 2, а то и на 3 метра ниже, чем рост «типичного» жирафа?
В общем, очевидно, что в отношении пресловутой «шеи жирафа» теория Дарвина, как говорится, «отдыхает по полной».
Но если мы предположим, что после такого яркого опровержения одной из самых знаменитых гипотез дарвиновской теории, кто-то из дарвинистов отказался от дарвинизма... то мы глубоко ошибёмся. Дарвинисты сегодня продолжают верить в теорию Дарвина, кажется, еще более рьяно, чем до провала с жирафами. Как же им это удаётся?
А у них есть палочка-выручалочка. Они теперь всё валят на половой отбор. То есть, теперь жирафы, «изо всех сил тянущиеся к самым высоким и вкусным веточкам» - уже не в моде у верующих дарвинистов. Теперь в моде гипотеза полового отбора - что удлинение жирафов произошло исключительно в угоду их дамам. В том числе, и удлинение самих этих дам.
Столь феерическая гипотеза заслуживает того, чтобы разобрать её более подробно. Однако сначала подведем предварительные итоги. Итак:
- Старая сказка, рассказанная когда-то давным-давно самим дедушкой Дарвиным, сегодня рухнула с весьма оглушительным треском.
- Однако никакое, даже самое громкое обрушение гипотезы не действует отрезвляюще на верующих дарвинистов. Новые гипотезы у них вообще никогда не заканчиваются. Дарвинисты просто не в состоянии сказать самим себе:
- Хватит, достаточно. Уже ясно, что дарвиновская исследовательская программа не в состоянии дать объяснение исследуемому биологическому факту.
И начать поиск в рамках других исследовательских программ.
Вместо этого верующие дарвинисты всё продолжают и продолжают выдвигать новые гипотезы в рамках чисто биологической целесообразности, взамен старых и уже рухнувших. Их не смущает, что выдвигаемые гипотезы становятся всё нелепее, доходя уже до откровенных анекдотов.
Это же надо было до такого додуматься – если самцы жирафов лупят друг друга своими головами, используя при этом длинные шеи в качестве «рычага» (наподобие плётки), то значит, именно поэтому жирафы и эволюционировали в столь удивительных созданий. Какую беззаветную веру в дарвинизм надо иметь, чтобы представлять себе, как самцы жирафов миллионы лет неуклонно лупят друг друга именно головами. А не толкаются, например, боками (как они нередко поступают в реальности), или не прижимают к дереву (как они тоже иногда поступают). И вследствие этой неуклонной тяги к драке именно с помощью голов, жирафы и приобрели весь тот потрясающий комплекс признаков, который мы только что перечислили выше.
Считается, что половой отбор должен приводить к половому диморфизму (по соответствующему признаку). В качестве примеров можно привести обильно рогатых самцов оленей и их почти безрогих самок. Или самцов павлинов с потрясающими «хвостами» и вообще бесхвостых пав. Однако самки жирафов почему-то решили приобрести себе точно такие же пропорции (шеи и ног), что и у самцов. Несмотря на то, что подобные пропорции излишни для самих самок (для их прямого выживания) и следовательно, естественный отбор должен был убирать у самок все подобные отклонения. Понятно, что такой половой отбор самцов, который приводит к развитию вредных признаков у самок - должен заканчиваться примерно там же, где и начинался.
В общем, получается - только из-за того, что самцы жирафов упорно предпочитали лупить друг друга именно головами (с помощью шей) и никак иначе, весь этот биологический вид так морфологически «перекосило» в ходе эволюции, что мы сейчас ходим и поражаемся этой удивительной внешности. Причем перекосило не только борющихся самцов, но и совсем не борющихся самок. Возникает вопрос – а не легче ли было естественному отбору просто отбраковать тех самцов, которые упорно лупили друг друга именно головами, заменив их на таких самцов, которые предпочитали разбираться друг с другом более простыми и естественными способами?
И еще непонятно, зачем в ходе «напряженной борьбы шеями» удлинялись ноги? Просто за компанию? А зачем удлинялся язык, который у жирафов тоже очень длинный?
Или, может быть, ноги исходных жирафов уже были такими длинными, удлинившись еще раньше (еще до начала удлинения шеи) под действием каких-то других причин, не связанных с половым отбором? Но тогда цельный дизайн жирафа, весь как бы направленный ввысь (от кончиков длиннющих ног до кончика длинного языка через рекордно длинную шею) – это просто такое случайное эволюционное совпадение, произошедшее под действием разных эволюционных сил, никак не связанных друг с другом?
Интересно, кто-нибудь в здравом уме и трезвой памяти (за исключением, конечно, самих дарвинистов) сможет поверить в такие «случайные совпадения»?
И как в таком случае жираф пил (с еще короткой шеей, но уже при таких ногах)? Неужели вообще ползком?
Верующих дарвинистов не останавливает и то обстоятельство, что представить себе, каким образом самцы жирафов лупили друг друга своими головами с помощью гипотетических исходных (коротких) шей довольно затруднительно. Это были бы весьма слабые удары. Скорее, наверное, щекотание (чем удар). Ведь способ, которым жирафы наносят удары головой, весьма специфичен.[47] Чтобы получился именно удар (а не щекотание) тем способом, каким самцы бьют сейчас, шея самца должна быть уже достаточно длинной. Таким образом, соревнование самцов с помощью специфической «борьбы шеями», скорее всего, является не причиной, а наоборот, следствием того, что у жирафа такая длинная шея. То есть, самцы потому и бьют головой, используя шею наподобие плетки, потому что эта шея достаточно длинна, чтобы ею можно было наносить удары именно таким способом. Интересно, что даже сами авторы гипотезы «борьбы шеями» (Simmons & Scheepers 1996) в итоге признали это обстоятельство.
Помимо самих авторов, эту гипотезу проверяли и другие исследователи. Как я уже говорил выше, в рамках предположения о возникновении длинной шеи в результате борьбы самцов, ожидается, что у жирафов должен наблюдаться половой диморфизм по этим признакам. А именно, шеи самцов должны быть сравнительно более длинными, чем у самок, а головы – сравнительно более крупными и тяжелыми (поскольку именно головами самцы и наносят друг другу удары). Авторы работы (Mitchell et al., 2009) измерили и сравнили соответствующие пропорции самцов и самок жирафов. И выяснили, что пропорции обоих полов не различаются по озвученным признакам. Поэтому авторы работы сделали вывод, что половой отбор не имел отношения к возникновению длинных шей жирафов. Вот конкретная цитата прямо из резюме статьи (Mitchell et al., 2009):
…Мы заключаем, что морфологические различия между самцами и самками – минимальны, что те различия, которые существуют, могут быть обусловлены большей общей массой самцов, и что половой отбор не является причиной происхождения длинной шеи у жирафов.
Наконец, возникает вопрос, а чего это естественный отбор, который, как известно, «не знает никаких границ приличия», ограничился только маленькими и тупыми рожками на голове жирафа? Почему половой отбор, сумев столь сильно изменить предполагаемые исходные пропорции самца (видимо, в результате очень сильного давления отбора?) и даже сумев «исказить» соответствующие пропорции самки (которая тут вообще ни при чем)… в течение всех этих гипотетических миллионов лет не позаботился о том, чтобы снабдить самца жирафа каким-нибудь более эффективным оружием (за самку), чем просто голова? Например, насколько эффективней была бы та же самая голова, но с острыми рожками? Какое преимущество сразу же получил бы жираф, сражающийся такой головой? И почему в ходе столь серьезного накала полового отбора, который сумел столь сильно (гипотетически) «искорежить» исходные пропорции животного, ни один самец жирафа до сих пор не додумался ударить другого самца своим по-настоящему смертельным оружием – передними копытами? Как известно, ударами передних копыт жираф вполне свободно убивает льва. Почему же еще никто не видел, чтобы один самец жирафа пнул другого самца жирафа передними копытами, хотя бы по ногам? Ради чего жираф демонстрирует столько благородства? Ведь своими головами жирафы бьют друг друга весьма энергично, отнюдь не на шутку, а как говорится, «от души». В результате, дело периодически доходит до нокаутов. Причем в нокаут может попасть не только жираф, которого ударили, но и жираф, который сам ударил (неудачно). И это не теоретическое предположение, а реально происходящие курьезы. Ведь удары всё-таки наносятся головой, а голова, как известно, далеко не самая неуязвимая часть тела (во всяком случае, у жирафов). Так почему же жирафы упорно бьют именно головами, но никогда – копытами?
Давайте представим себе, какое немедленное преимущество получил бы некий мутантный самец жирафа, который без зазрения совести начал бы пинать передними копытами всех окружающих самцов? Ведь окружающие самцы – еще не имеют такой мутации, и соответственно, ответить ему тем же – просто не смогут. Наверное, все окрестные самки сразу же оказались бы в распоряжении данного мутанта. Мало того, этот мутантный жираф таким образом решил бы, наверное, еще и проблему конкуренции за пищу (если бы такая проблема вообще была).
Тем не менее, самцы жирафов почему-то пинают кого угодно, но только не друг друга. Таким образом, естественный отбор в очередной раз поражает нас чудесной неисповедимостью своих путей.
Когда-то давно (еще в «1976-мохнатом году») известный биолог-публицист Ричард Докинз попытался решить проблему явно наблюдаемого «благородства животных»,столь удивительного в русле представлений о «беспощадной борьбе за выживание». В своей нашумевшей книге «Эгоистичный ген» Докинз долго рассуждал о теоретических «ястребах» и «голубях», и о тех преимуществах, которые «голуби», неизменно проигрывая «ястребам», тем не менее, получают в конечном итоге. Он объяснил преимущество «голубей» тем, что при увеличении числа «ястребов» в популяции, их выигрыш сменяется проигрышем - «ястребов» становится слишком много, они слишком часто наталкиваются именно друг на друга, получают увечья и гибнут.[48] А вот теоретический трусливый «голубь», ускользнувший из всех поединков с «ястребами» самым простым способом (постыдного отступления), получит преимущество – просто потому, что сохранит себя в целости. В конечном итоге (по Докинзу), в популяции должна победить некая (устойчивая) «эволюционно стабильная стратегия» - популяция автоматически придёт к определенному (устойчивому) соотношению частоты встречаемости разных типов поведения («ястребов» и «голубей»). И это устойчивое соотношение будет далее автоматически поддерживаться естественным отбором.[49]
Понятно, что теоретический «трусливый голубь» Докинза призван теоретически объяснить наблюдаемые в природе факты «благородства животных». Например, почему хищники не охотятся друг на друга, а предпочитают, допустим, разграничивать территорию? Потому что (по Докинзу) риск получения повреждений в ходе такой охоты слишком велик и не окупается возможным выигрышем. Следовательно, такая стратегия была бы эволюционно нестабильной. То есть, особи, склонные к подобному поведению, будут удаляться из популяций естественным отбором.
Или почему самцы многих видов, конкурируя за самку, не забивают друг друга до смерти, а ограничиваются «благородным турниром по строгим правилам»? Или вообще совершают только ритуальные демонстрации и стилизованные представления? А потому что такое поведение всё-таки обеспечивает (тот или иной) выигрыш в выживании и размножении, с учетом риска получения сильных ранений в случае серьезного поединка между самцами. Поэтому естественный отбор и отобрал (автоматически) только таких самцов, которые способны лишь к мирным демонстрациям и стилизованным представлениям.
Эти рассуждения Докинза были бы прекрасны, если бы хоть что-нибудь могли объяснить в реальной природе. Особенно наблюдаемое разнообразие жизни. Ведь в живой природе мы с избытком наблюдаем примеры видов, где имеются сплошные «ястребы» (т.е. все самцы дерутся между собой очень жестоко, вплоть до смерти). И с таким же избытком наблюдаем биологические виды, где господствуют одни лишь «голуби», то есть, все самцы используют в отношении друг друга лишь демонстрации, декорации или сугубо безобидное «турнирное оружие».[50]
Как объяснить, что, например, морские слоны дерутся до увечий или даже до смерти, и каждый морской слон имеет огромный гарем из самок, а самец жирафа никогда не использует действительно смертельных приемов в отношении своего соперника? А у многих видов самцы из-за самок вообще не конфликтуют, предпочитая, например, позировать перед самками (в надежде на их выбор), несмотря на то, что в это время в зоне их прямой видимости находятся другие самцы, тоже пытающиеся завлечь самку.
Случай с морским слоном можно легко объяснить, используя язык самого Докинза. Можно сказать, что конкретный самец морского слона, действительно, подвергается смертельному риску во время поединков. Но и возможный выигрыш (единственного победившего) тоже исключительно высок - это доступ к практически безграничному размножению (гарему из пары десятков самок).
Но точно такой же тип взаимоотношений между самцами мог бы сложиться и в случае жирафов. Ну стали бы самцы жирафов активно бить друг друга копытами. Ну, покалечили бы друг друга. Ну и что? В этом случае изменилось бы только то, что все окружающие самки стали бы приносить детей только от самых-самых приспособленных самцов жирафа. У выживших (редких) талантливых бойцов (или счастливчиков) просто увеличился бы гарем из доступных им самок и, соответственно, число детей. Вот и всё. То есть, из жирафа просто получился бы очередной «морской слон». Который и так уже имеется в живой природе, тем самым автоматически доказывая, что подобный способ существования биологического вида – вполне возможен.
Так почему же ни один самец жирафа, несмотря на мощную немедленную выгоду, тем не менее, до сих пор не догадался пнуть другого самца копытом?
По Докинзу (еще раз) это чревато излишним размножением «ястребов» в таких популяциях, где все самцы станут жестоко драться без правил, и в результате часто гибнуть. Поэтому в долгосрочной перспективе, самцам жирафов невыгодно переходить от благородных турниров по правилам к травмоопасным боям без правил. Но ведь естественный отбор не «мыслит» категориями отсроченной выгоды. Он «мыслит» только категорией немедленной выгоды – что выгодно для размножения прямо сейчас, то и будет отобрано. И потомство оставит именно мутантный «самец-убийца», а не «благородные голуби». Да, это чревато тем, что через сотни поколений все самцы в популяциях жирафов будут представлены именно «самцами-убийцами». И в результате риск погибнуть в жестоких поединках для них многократно возрастёт. Но это случится уже потом, через много поколений. И когда это случится, появление в такой популяции (обратно) «благородного рыцаря»... будет встречено дружным хохотом всех самцов-жирафов, предпочитающих бои без правил. И даже если этот одинокий «благородный рыцарь» всё-таки сумеет передать свои гены следующему поколению (подкравшись к одиноко стоящей самке в то время, пока другие самцы убивают друг друга в смертельной битве), то всё равно, подобная стратегия никогда не сможет полностью завоевать популяцию (обратно). А возможна лишь в виде редкой (эпизодической) «примеси» на фоне общего доминирования «самцов-убийц».
Таким образом, ответить на вопрос «почему мы наблюдаем в природе благородство животных» (у многих биологических видов) - в рамках представлений об «эволюции в ходе борьбы за выживание», на самом деле, вообще нельзя (в принципе). Потому что данная теория постулирует, во-первых, возможность свободных изменений видов практически в любых направлениях, а во-вторых, в русле этой теории имеется только одна реальная ценность – биологическое выживание (любыми эффективными способами). В рамках таких представлений, самцы жирафов уже давным-давно должны были «скатиться» от «благородной борьбы шеями» до смертельного «мордобоя» вообще без каких-либо правил. И это и была бы самая эволюционно стабильная стратегия из всех возможных. А вот любые «благородные демонстрации» - это эволюционно нестабильное поведение. Потому что «благородные рыцари» беспомощны перед мутантными «бойцами без правил». Причем, во-первых, согласно теории, мутантные «бойцы без правил» просто обязаны появляться время от времени. Во-вторых, появившись, они получают немедленное преимущество над «благородными рыцарями». А в-третьих, даже победив всех «благородных рыцарей» и расплодившись в популяции сверх всякой меры, эти «бойцы без правил» не сделают саму популяцию менее приспособленной. Скорее, такая популяция даже повысит свою приспособленность, так как дети станут рождаться только от «самых-самых» самцов. И практическое подтверждение справедливости подобных рассуждений – это вагон и маленькая тележка именно таких биологических видов в реальной природе.[51]
Таким образом, получается, что эволюционно стабильной стратегией здесь, в любом случае, является не «благородное соревнование» самцов, а реальная борьба этих самцов без соблюдения каких-либо правил. Во всяком случае, немедленная готовность перейти от «благородного соревнования» к «борьбе без правил» при малейшей необходимости.[52] Поэтому объяснить существование в природе видов, демонстрирующих исключительно (и только) «благородное соревнование» - современная теория эволюции не может.
Итак, современная теория эволюции способна выдвигать по поводу удивительного дизайна жирафа лишь откровенно нелепые истории. Про то как:
- Cамцы жирафов несколько миллионов лет подряд били друг друга исключительно с помощью шей (а не бодались, не толкались боками, не наносили друг другу удары копытами или рогами)… и в результате у самок жирафов: эволюционно выросла шея, эволюционно выросли ноги, эволюционно увеличилось сердце и легкие, возникли специальные запирающие клапаны в кровеносных сосудах, особо упрочнилась кожа, и дети стали рождаться задними ногами вперед с аккуратно сложенной шеей.
Что и говорить, это просто «прекрасная» гипотеза, представляющая современный дарвинизм во всём его поразительном «великолепии».
А теперь давайте сравним неуклюжие попытки объяснений уникальной внешности жирафа в русле дарвинизма, с объяснениями в рамках недарвиновских исследовательских программ. Например, почему жираф имеет такую внешность в рамках концепции разумного дизайна?
Многие научно-популярные заметки, посвященные жирафу, начинаются примерно такими словами:
Жираф – одно из красивейших животных нашей планеты…
Собственно, всё. Этой фразы уже достаточно, чтобы объяснить уникальный дизайн жирафа в рамках концепции разумного замысла. То есть, жираф имеет такую внешность – для красоты, в качестве одного из вариантов почти бесконечного разнообразия стилей дизайна разных живых существ.
И смысл целого комплекса специальных приспособлений, которые помогают жирафу выживать, несмотря на столь радикальную внешность, тоже становится ясным – этот комплекс предназначен именно для того, чтобы жираф успешно выживал, имея столь замечательную внешность.
И наконец, почему жирафы не пинают друг друга копытами, а борются именно с помощью своих длинных шей? А потому, что это просто программа полового поведения у данного вида заложена такая (а не другая). Поскольку у жирафа почти всё «крутится» вокруг его шеи, то, наверное, не удивительно, что и в программе брачного поведения эта шея тоже задействована.
Как видим, в рамках недарвиновских исследовательских программ, и внешность жирафа, и его поведение объясняется настолько просто и естественно, насколько это вообще возможно.
На этом я, пожалуй, закончу с нашими удивительными жирафами.
И переключу Ваше внимание на другое африканское животное, которое знаменито ничуть не меньше, чем жираф. Речь пойдёт об абсолютном мировом рекордсмене по бегу на короткие дистанции – о знаменитом африканском гепарде.
Дело в том, что совсем недавно гепарды тоже неожиданно подложили «свинью» бедным дарвинистам. Последнее исследование гепардов в дикой природе очень удивило зоологов. Оказалось, что высшая скорость, развиваемая гепардами в погоне за добычей в дикой природе, обычно не превышает 10-18 м/c. В среднем, высшая скорость в охотничьих забегах гепардов составила всего 15 м/c, или 54 км/ч (Wilson et al., 2013). В то же самое время, эти же гепарды показали, что в принципе, они способны бегать в природе с максимальной скоростью 26 м/c (93 км/ч).[53] А в неволе были получены цифры и до 29 м/c (104 км/ч).
Получается, что гепарды в природе преследуют свою добычу… на средней скорости. То есть, не сильно напрягаясь, эдаким «бегом трусцой». Тем не менее, гепарды добивались успеха примерно в одной трети случаев уже при максимальной скорости забега всего 12 м/c (43 км/ч). А при максимальной скорости 15 м/c (54 км/ч) успех сопутствовал им уже примерно в половине случаев погони (Рис. 25). Интересно, что при дальнейшем увеличении максимальной скорости забега от 15 до 20 м/c доля успешных исходов погони практически не возрастала. А при скорости выше 20 м/c, кажется, даже убывала (Рис. 25):
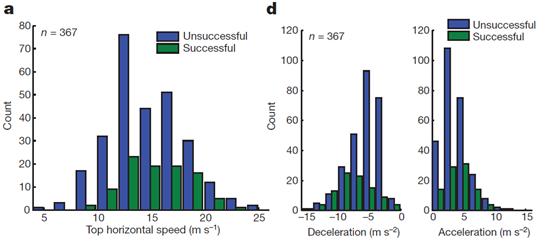
Рисунок 2 5 . Слева (а) график, показывающий число (по вертикали) охотничьих забегов гепардов, проделанных ими с определенной максимальной скоростью (м/c, по горизонтали) из общего числа измеренных 367 забегов. Синие столбики – неуспешные охоты гепардов. Зеленые столбики – успешные охоты гепардов. Справа ( d ) графики, показывающие величины максимального торможения и ускорения, продемонстрированные гепардами в 367 забегах (по вертикали – число забегов, в которых было продемонстрировано ускорение или торможение определенной величины).[54] Рисунки взяты из работы (Wilson et al., 2013).
Очевидно, что получается очередной биологический курьёз – «свидетели Дарвина» многими десятилетиями рассказывали наивным читателям страшные истории о том, как бедные гепарды сурово отбирались естественным отбором в ходе борьбы за существование, где в стремительных забегах за самыми быстроногими антилопами мира выживали только самые скоростные гепарды. И в результате, через многие-многие тысячи лет этого беспощадного отбора и «отобрался» (автоматически) современный гепард, этот шедевр природы, почти каждая черта которого говорит нам о скорости, скорости и еще раз скорости.
Однако результаты последнего исследования радикально изменили ситуацию. Теперь верующим дарвинистам необходимо срочно придумывать какую-то другую сказку (почему гепард стал таким, какой он есть). Ведь теперь возникает закономерный вопрос – а как это гепарды умудрились развить в себе способность достигать скоростей 25-29 м/c, если в природе они вполне успешно «гоняются» за добычей с максимальной скоростью 15 м/c? Кто тот дополнительный тренер (помимо естественного отбора), которому так не понравились скоростные нормативы, предъявляемые гепардам в реальной природе, что он натренировал гепардов бегать со скоростью, в полтора раза большей?
Здесь можно, конечно, что-нибудь придумать. Например, предположить, что гепарды не демонстрируют максимальной скорости забега потому, что убегающая жертва может в любое мгновение повернуть, и гепард должен успеть повернуть за ней тоже. А это, наверное, было бы трудно сделать при слишком больших скоростях. Вот гепарды и осторожничают. Однако такое рассуждение содержит слишком много слов. Если сократить все лишние слова, то получится, что мы предположили следующее - маневренность бега гепарду гораздо важнее, чем высокая скорость этого бега.
Но от такого предположения наш предыдущий вопрос всё равно не снимается. Действительно, если в природе гепарды отбирались, в основном, на маневренность бега (а не наивысшую скорость), тогда каким же образом они так «отобрались», что теперь могут бегать в полтора-два раза быстрее, чем это необходимо?
Если, допустим, у ястребов шел отбор именно на маневренность полёта, то мы и должны ожидать получить «на выходе» ястреба (исключительно маневренную птицу), но никак не стрижа (рекордсмена скорости в горизонтальном полете).
Таким образом, здесь мы имеем случай, полностью аналогичный пресловутой «шее жирафа». Ведь дарвинисты нам тоже рассказывали страшные истории, что жирафы стали такими из-за беспощадной борьбы с голодом, огромной необходимости дотягиваться до самых верхних (оставшихся) веточек акации. И в конце концов жираф так «отобрался естественным отбором», что стал ростом 5-6 метров… А потом оказалось, что жирафы почему-то предпочитают питаться с высоты всего примерно 3 метра.
В общем, получается, что и величавые 5-метровые жирафы, и великолепные гепарды, бегущие со скоростью 100 км/ч (при необходимых 55) – это результат чего-то другого, но только не «естественного отбора более приспособленных».[55]
Ну а сейчас нас ждут совсем свежие легенды дарвинизма. Так сказать, «свежеиспеченные» (или свеже-жареные). Еще не надоевшие так, как древние сказки про «естественный отбор голодающих жирафов» или про «эволюцию гепардов, игравших в догонялки с антилопами».
НОВЫЕ ЛЕГЕНДЫ ДАРВИНИЗМА
Итак, что же делать верующему дарвинисту?
Ведь строгая (эмпирическая) доказательная база дарвинизма - крайне слаба. И это на фоне разрушения старых, известнейших сказок дарвинизма. Только что я рассказал о том, как в начале 21 века была разрушена старая сказка Дарвина – «Почему у жирафа такая шея»? Точно так же, в начале 21 века в ходе эмпирических проверок рухнула и другая, еще более знаменитая сказка дедушки Дарвина – «Почему у павлина такой хвост»? Об этом трагическом крушении я подробно расскажу позже (уже в следующей книге).
Получается, что в начале 21 века, спустя 150 лет после опубликования Дарвиным его теории, дела у дарвинизма в области эмпирических доказательств стали даже хуже, чем были 150 лет назад. Строгих примеров эволюции форм (куда-нибудь за рамки внутривидовой изменчивости) под действием естественного отбора – так и не установлено, в то время как некоторые старые прогнозы этой теории были проверены и уже опровергнуты.
И что же делать дарвинисту в такой ситуации? Некоторые особо верующие дарвинисты нашли выход. Если новых эмпирических доказательств нет, то нужно их придумать. Действительно, не пропадать же бесследно славным традициям Геккеля? Если Геккель, за отсутствием доказательств эволюции в начале истории дарвинизма решил такие доказательства просто нарисовать, то почему бы сегодняшним верующим дарвинистам не придумать новые легенды дарвинизма?
Тем более что это легко сделать. Во-первых, мы все хорошо помним наши «школьные годы чудесные» (С), и благодаря этим «чудесным годам», теперь твердо знаем, что «дарвинизм - есть единственно верное учение, давно доказанное наукой». Во-вторых, современная биология накопила уже много установленных примеров тех или иных изменений организмов во времени. Чаще всего, эти изменения являются колебаниями внутривидовой изменчивости, но не всегда. И если теперь взять те или иные установленные факты изменчивости, и просто переписать их идеологически правильно, утверждая, что это и есть «строго доказанный пример эволюции под действием естественного отбора», и еще расставить восклицательные знаки в нужных местах... то верующие дарвинисты получат тот самый долгожданный пример «наблюдаемой эволюции», который им нужен. Можно еще больше усилить этот пример, сделать его более «говорящим», применив следующие нехитрые приёмы:
1) исказить смысл тех или иных фактических подробностей, хотя бы слегка, но в нужную сторону;
2) о чём-нибудь умолчать, посчитав это «несущественным»;
3) какие-то особо красочные детали вообще добавить лично от себя.
И всё. Теперь можно громко заявлять о «новом установленном примере эволюции под действием естественного отбора». Таким образом, на свет родится свежая легенда, которая так нужна сегодня верующим дарвинистам. Ведь известно, что «коварные креационисты» не дремлют, а покушаются на «хорошую науку» самым наглым образом. Подумать только, креационисты совсем не довольствуются тем, что «единственно верное учение Дарвина» справедливо просто потому, что оно верно. Эти наглецы еще требуют каких-то реальных доказательств справедливости этого учения! Что и говорить, весьма коварные типы. В связи с этим, в 2007 году Парламентская ассамблея Совета Европы произвела на свет свою знаменитую резолюцию про «Опасность креационизма в образовании». И это правильно. Потому что у Европы больше не осталось проблем, кроме как предупреждать народы об опасности креационизма. Теперь осталось только принять еще 23 резолюции об «опасности креационизма». И тогда «единственно верное учение Дарвина» наконец одержит славную победу.
Однако давайте будем честны сами с собой. Резолюция ПАСЕ - это ведь всё-таки не научное подтверждение теории эволюции. А просто некое подобие идеологического кликушества. В священной войне с «опасными креационистами» дарвинистам было нужно более мощное и современное оружие. Поэтому свежая «легенда дарвинизма» уже давно «вопила» о своём создании. Тем более что старые «доказательства эволюции» уже изрядно поистаскались.
Стоит ли удивляться, что в конце концов великое событие свершилось. Верующих дарвинистов, наконец, снабдили свежей научной «легендой дарвинизма». И эту (очередную) «легенду дарвинизма» креационистам теперь опровергать и опровергать. Ибо попробуй опровергнуть то, что уже записано в самой википедии.
Рождение легенды происходит особенно легко в том случае, когда её создателем является какой-нибудь популярный ученый, известный широким массам. Особенно, уже прославившийся в деле священной борьбы с креационизмом. Ведь рядовые верующие дарвинисты склонны читать и слушать только таких ученых (и больше никаких). Даже если этот знаменитый ученый рассуждает о тех областях биологии, которые представляет себе лишь в самых общих чертах. Но кто будет обращать внимание на такие мелочи? Главное, чтобы новая легенда дарвинизма была создана профессиональным дарвинистом. И тогда дарвинисты-любители, вооружившись этой новой легендой, снова смогут смотреть на «этих невежественных креационистов» свысока. Взглядом, преисполненным глубокой мудрости и легкого сожаления.
В принципе, под это описание («создание дарвиновских легенд») подходят почти все примеры «наблюдаемой эволюции», которые я уже разобрал выше. Однако сейчас я хочу специально разобрать еще один конкретный пример рождения эволюционной легенды. Потому что эта эволюционная легенда родилась совсем недавно. И в настоящее время прямо-таки на глазах начинает обрастать дополнительными красочными деталями, которые всё больше превращают её из довольно безобидного биологического факта в убойное доказательство наблюдаемой эволюции под действием естественного отбора.
Главными действующими лицами в этой «классике Геккелевского жанра» выступят:
Создатели легенды:
1. Доктор биологических наук, палеонтолог А.В. Марков, лауреат премии «Просветитель» и член комиссии по борьбе с лженаукой.
2. Доктор биологических наук, биоинформатик и генетик М.С. Гельфанд (тоже активный борец с лженаукой).
Разрушители мифов:
Автор этих строк.
Подобно тому, как «легким движением руки брюки превращаются в шорты» (С), мы сейчас сможем проследить, как достаточно безобидный биологический факт превращается в яркое доказательство эволюции под действием естественного отбора... с помощью всего нескольких небольших неточностей в изложении, излишней лаконичности (приводящей к отсутствию важных подробностей) и одной дополнительной выдуманной детали.
Первым на сцену выходит доктор биологических наук Александр Марков.
В своей заметке «Цепная реакция видообразования» Александр решил рассказать читателям про наблюдаемую эволюцию яблоневых мух (Марков, 2009а). Тех, кто еще не слышал об этой новой «строго установленной легенде дарвинизма» ввожу в курс дела обширной цитатой из самого Александра Маркова. Слово автору.
Резюме заметки:
...Мухи Rhagoletis pomonella, личинки которых живут в плодах боярышника, на глазах у ученых и садоводов за 150 лет «отделили» от себя новый вид, ставший опасным вредителем яблонь. Как выяснилось, это стало стимулом для видообразования у наездников, паразитирующих на этих мухах. Со своей стороны, наездники стимулируют видообразование у мух, заставляя их переходить на новые растения.
Основной текст:
...В последнем номере журнала Science американские биологи сообщили, по-видимому, о первом хорошо документированном случае согласованного симпатрического видообразования у мухи Rhagoletis pomonella и наездника Diachasma alloeum, личинки которого паразитируют в личинках мух.
Мухи Rhagoletis pomonella живут в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Предпосылки для разделения Rhagoletis pomonella на два вида появились в XVII веке, когда европейские колонисты привезли в Новый Свет саженцы яблонь. Первое упоминание об американских яблонях относится к 1647 году. Спустя два столетия, в 1864 году, садоводы впервые заметили нового вредителя — белых червяков, которые вдруг стали пожирать их урожай.
Червяки оказались личинками мухи Rhagoletis pomonella, которая до сих пор откладывала яйца только на плоды боярышника.
За прошедшие с тех пор полтора века боярышниковая и яблоневая формы мух стали так сильно различаться по своему поведению, брачным предпочтениям и генетике, что сегодня их уже вполне можно считать двумя разными видами. Они почти не скрещиваются друг с другом (уровень гибридизации не превышает 4–6%); яблоневый вид спаривается почти исключительно на яблонях, а боярышниковый — на боярышнике. По-видимому, именно эта манера спариваться на «своем» кормовом растении способствовала быстрому видообразованию. У насекомого с такими привычками (а надо сказать, что огромное множество видов растительноядных насекомых предпочитают спариваться на своих кормовых растениях) для видообразования может быть достаточно небольшого случайного генетического изменения, ведущего к смене обонятельных предпочтений. В принципе для зарождения «яблоневого» вида было достаточно одной мутантной самки, которой запах яблок понравился больше, чем запах боярышника, и которая передала эту особенность своему потомству ...
…Позднее энтомологи обнаружили еще два вида Rhagoletis, которые размножаются на чернике и снежноягоднике. Внешне все четыре вида практически неотличимы друг от друга. Генетически они тоже очень близки, хотя некоторые различия всё же имеются.
На мухах Rhagoletis паразитируют наездники Diachasma alloeum, которые откладывают яйца в личинок мух. Личинка наездника развивается в личинке мухи и в конце концов убивает ее...
…Исследователи решили проверить, не привело ли появление нового вида мух также и к появлению нового вида наездников. В анализ были заодно включены и наездники, паразитирующие на черничных и снежноягодниковых мухах.
Проведенные ранее эксперименты показали, что и самцов, и самок всех четырех видов мух привлекает запах плодов «своего» растения, тогда как запахи плодов трех других растений действуют на них как репелленты.
А что же наездники? Оказалось, что у наездников, чьи личинки развивались в каждом из четырех видов мух, наблюдаются точно такие же запаховые предпочтения. Теперь нужно было проверить, нет ли у наездников, как и у их жертв, обычая спариваться возле плодов «своего» растения. Это тоже подтвердилось в ходе полевых наблюдений.
Авторы провели также генетический анализ четырех групп наездников и выявили небольшие различия между ними по частоте встречаемости некоторых аллелей, причем в ряде случаев аллель, часто встречающийся у одной из четырех форм, вовсе не встречался у других. Это свидетельствует об отсутствии значительного обмена генами (гибридизации) между четырьмя разновидностями наездников.
Генетический анализ дал еще один важный результат: он показал, что «яблоневая» разновидность наездников не могла произойти напрямую от «боярышниковой» разновидности. Скорее всего она произошла от черничной разновидности, с небольшой примесью генов от боярышниковой. Иными словами, диверсификация в данном случае не была строго согласованной. Яблоки как новая среда обитания были освоены боярышниковыми мухами, однако яблоневые мухи (тоже как новая среда обитания) были «освоены» не боярышниковыми, а черничными наездниками, возможно лишь с некоторой примесью генов боярышниковых наездников, полученной в результате гибридизации.
Виды мух Rhagoletis отличаются друг от друга, помимо генетических особенностей и брачного поведения, еще и сроками вылупления из куколок. Эти сроки приурочены ко времени созревания соответствующих плодов. Со снежноягодниковым видом исследователи пока не разобрались (не сумели добыть материал в достаточных количествах), а по остальным трем видам картина получилась следующая. Первой на востоке США созревает черника, за ней — яблоки и последним — боярышник. Соответственно, из куколок первыми выходят черничные мухи, потом яблочные и, наконец, боярышниковые.
Авторы обнаружили, что у трех пород наездников Diachasma alloeum наблюдаются точно такие же различия по времени выхода из куколки, как и у их хозяев. Первыми вылупляются черничные наездники, затем яблочные, а боярышниковые появляются на свет последними. Совершенно очевидно, что эти различия являются наследственными. Авторам даже удалось обнаружить корреляцию между некоторыми из изученных ими генетических маркеров (микросателлитов) и сроками вылупления наездников. Иными словами, хотя гены, непосредственно регулирующие срок выхода из куколки, еще не обнаружены, уже приблизительно известно, где их искать: рядом с теми маркерами, аллельное состояние которых коррелирует со временем выхода. Следует также иметь в виду, что различие по срокам выхода из куколки автоматически ведет к снижению вероятности гибридизации между разными породами (зарождающимися видами) наездников, которые во взрослом состоянии живут совсем недолго — около двух недель.
В целом полученные данные убедительно показали, что видообразование у мух Rhagoletis стало стимулом для видообразования у их паразитов — наездников Diachasma. Самое интересное, что связь между видообразованием у мух и наездников может быть взаимной. Есть все основания полагать, что переход жертв на новые кормовые растения стимулируется паразитами. Перейдя на новое растение, жертва во многих случаях частично или полностью избавляется от паразитов, «привыкших» искать ее на старом кормовом растении. Перейдя с боярышника на яблоки, мухи действительно получили преимущество в этом отношении. Хотя наездники вскоре последовали за ними на новое кормовое растение, тем не менее до сих пор уровень зараженности личинками наездника у личинок яблоневых мух на 70% ниже, чем у боярышниковых. Отчасти это связано с тем, что яблоко крупнее плодов боярышника и в нём легче спрятаться от длинного яйцеклада самки наездника. Интересно, что яблоки являются менее благоприятной средой обитания для личинок мух Rhagoletis, чем плоды боярышника. В боярышнике личинки выходят из половины отложенных мухой яиц, а в яблоке — только из четверти.
Таким образом, в переходе с боярышника на яблоки были не только плюсы, но и минусы. Это можно рассматривать как косвенный аргумент в пользу того, что паразиты были важным фактором, способствовавшим видообразованию у мух. Если бы не паразиты, от которых яблоки дают лучшую защиту, естественный отбор не поддержал бы переход мух на менее подходящие для них плоды.
Итак, человеку, который прочитал этот текст Маркова, становится очевидно, что строго установлен по крайней мере один хороший пример образования нового вида буквально «на глазах» - это новый вид яблоневой мухи. Более того, за новым видом мух «последовали» наездники, паразитирующие на этих мухах, и как следствие, также сэволюционировали, возможно, тоже в новый вид.
Понятно, что для верующего дарвиниста столь яркий пример наблюдаемой эволюции, причем сразу до нового биологического вида, да еще и вызвавший эволюцию еще одного (связанного) вида... такой пример для дарвиниста-проповедника является более ценным, чем клад золотых монет времен татаро-монгольского ига.
Поэтому «яблонные мухи» пронеслись по среде верующих дарвинистов подобно урагану. И уже через полгода чуть ли не каждый второй верующий дарвинист, с которым я беседовал о проблемах эволюции, приводил мне этот пример «безусловного видообразования под действием естественного отбора».
Однако следует обратить внимание, что с естественным отбором в данном примере пока совсем не густо. Собственно, его пока нет вообще. Яблонные мухи могли перейти с боярышника на яблоню (по словам А.В. Маркова) буквально из-за «одной мутантной самки, которой запах яблок понравился больше, чем запах боярышника, и которая передала эту особенность своему потомству». Очевидно, что новая эволюционная сказка без естественного отбора выглядит еще логически незавершенной, неполной. Ведь нам нужен пример не абы какой эволюции (пусть даже и с образованием нового вида), а строго установленный факт эволюции именно под действием естественного отбора. То есть, новую эволюционную сказку хорошо было бы логически завершить.
Такое логическое завершение сделал уже второй соавтор легенды, доктор биологических наук А.М. Гельфанд, в телевизионной передаче «Мозговой штурм. Эволюция человека».[56]
В ходе беседы, когда речь опять зашла об установленных примерах эволюционных изменений, А.М. Гельфанд пересказал «историю яблоневой мухи» уже с добавлением новых, весьма красочных подробностей. Привожу полную цитату Гельфанда:
Это такой стандартный запрос учёным: «Покажите нам, как один вид превратился в другой».
Самый красивый пример – это история с яблоневой мухой в северной Америке. Там не было яблонь, их туда завезли в середине XIX века. Соответственно, часть популяции плодовой мухи, которая жила на боярышнике, переселилась на яблони, жила в яблоках.
С точки зрения конкретной мухи, это на самом деле плохо. Те мухи, которые живут на яблонях, у них меньше потомства, личинки хуже выживают. Казалось, это был шаг назад. Но на самом деле произошло это потому, что на мухе паразитирует наездник, который откладывает яйца в личинку мухи. Когда муха живёт в яблоке, наездник яйцекладом не достаёт. Тем самым, мухи хуже живут с точки зрения еды, но у них лучше защита от хищника.
Самая полная вершина этой картинки была, когда оказалось, что образовалась раса наездников с более длинными яйцекладами, которые специально паразитируют на тех мухах, которые живут в яблонях. И всё это случилось за 150 лет.
Таким образом, Гельфанд заострил внимание на том, что яблонные мухи на яблоках хуже выживают, но зато получают преимущество в их борьбе с наездниками. Бедные наездники не достают яйцекладом до личинок мух в яблоках, и поэтому (видимо) естественный отбор позаботился о том, чтобы всего за 150 лет отобрать более длинные яйцеклады новой расе наездников, паразитирующих на новой расе мух. То есть Гельфанд добавил к вышеприведенному рассказу Маркова несколько важных деталей, самой главной из которых явилось эволюционное изменение длины яйцеклада наездников. Таким образом, яблонные мухи (вместе с их наездниками) теперь стали демонстрироватьне только «строго установленный пример видообразования», но еще и «строго установленный пример эволюции под действием естественного отбора». Более того (и это самое главное обстоятельство, на которое я всё время обращал внимание читателя), теперь яблонные мухи (а точнее, их наездники) стали еще и примером морфологических изменений видов под действием естественного отбора, ибояйцеклад наездников стал длиннее в ходе эволюции.
Понятно, что этот рассказ Гельфанда тоже достаточно широко распространился по русскоязычному интернету (в узком кругу любителей эволюции). Дело дошло до того, что теперь каждый второй дарвинист рассказывал мне о «безусловном видообразовании у яблонных мух», а каждый третий - вспоминал при этом еще и об «удлинении яйцеклада наездников под действием естественного отбора». Что и говорить, интернет сегодня - великая образовательная сила.
В какой-то момент мне надоели постоянные отсылки дарвинистов к яблонным мухам, и я решил всё-таки узнать, что же там произошло на самом деле с этими обаятельными насекомыми. Не из научно-популярных ссылок, а из первоисточников, то есть, соответствующих научных статей и монографий. По собственному опыту (изучения научных публикаций на тему естественного отбора), я уже хорошо знал, что эволюционные изменения под действием естественного отбора в живой природе практически не происходят. Исключения из этого общего правила экстремально редки. Соответственно, и установленные примеры эволюции под действием именно естественного (или полового) отбора в научных публикациях тоже крайне редки. Поэтому мне и стало любопытно узнать, что же там такое случилось с яблонными мухами и их наездниками? Это просто очередное «эволюционное недоразумение», то есть, очередная «легенда дарвинизма», не соответствующая реальности? Или же это - то самое экстремально редкое исключение, когда естественный отбор наконец-то (!) вспомнил о своей обязанности быть «двигателем эволюции», и реально там чего-нибудь «сдвинул» у этих мух и наездников?
Изучение первоисточников быстро расставило всё по своим местам. И я сейчас, конечно, напишу, что там случилось на самом деле. Но есть одна трудность. Практически все области биологии серьезно насыщены какими-то своими, чисто биологическими подробностями. И чтобы обрисовать полную картину, мне придется сейчас разобрать много таких деталей (из биологии мух, наездников и яблонь). То есть, в двух абзацах описать ситуацию не удастся. Поэтому если читатель хочет разобраться с этим «примером эволюции», ему придётся набраться немного терпения, следя за теми биологическими подробностями, которые я сейчас озвучу.
Итак, первое, с чего мы начнём - в приведенных выше цитатах А.В. Маркова и М.С. Гельфанда я выделил жирным шрифтом те фразы, которые не соответствуют действительности. То есть, это либо искажение реальной ситуации, либо ошибка или неточность, либо добавление (от себя) фактов, не существующих в действительности.
Давайте кратко пройдемся по каждому пункту. Сначала разберем цитату Маркова:
А.В. Марков (пишет прямо в резюме собственной заметки):
...Мухи Rhagoletis pomonella, личинки которых живут в плодах боярышника, на глазах у ученых и садоводов за 150 лет «отделили» от себя новый вид, ставший опасным вредителем яблонь.
Разрушитель мифов:
На самом деле, никакой «новый вид» мухи Rhagoletis pomonella от себя не «отделили». В исходной статье (которую пересказывает Марков в своей заметке), авторы называют этих мух (перешедших на яблоки) - новой «host race», то есть, новой расой, которая перешла на новое пищевое растение. Ни о каком новом виде и речи быть не может. Не может быть просто потому, что авторы данной работы - это нормальные исследователи. Поэтому они не могут игнорировать заключение своих коллег систематиков (специалистов по данному таксону, роду Phagoletis), которые в ходе исследований установили, что «яблонных мух» следует считать именно новой пищевой расой. Но никак не новым видом. То есть, во всём мире считают, что яблонные мухи - это «новый вид», только два человека. Это палеонтолог Александр Марков и еще (наверное?) генетик и биоинформатик Михаил Гельфанд.
Это самая главная ошибка Маркова в его заметке. Он слишком легкомысленно относится к этому вопросу, потому что далек от этой области, и не понимает, что:
1) Новая раса и новый вид - это далеко не одно и то же, и это не мелочь, а важный вопрос, которому были посвящены многие научные публикации. Столь легко смешивать эти понятия способен только полный дилетант в этой области, которому (в виду его дилетантизма) «что воля, что неволя, всё равно» (С).
2) Конкретно в обсуждаемом случае имеются все основания, чтобы считать новую расу - именно расой, а не «новым видом». И ниже я объясню, почему это так.
Обратим внимание, что эта «небольшая неточность» в корне меняет содержание всей заметки. Из достаточно безобидного биологического факта этот пример сразу же превращается в очень серьезный пример «наблюдаемой эволюции», причем зашедшей так далеко, что образовался новый вид. Следовательно, возможность видообразования теперь «строго доказана». Я же говорю – «легким движением руки брюки превращаются в шорты» (С).
А.В. Марков:
Как выяснилось, это стало стимулом для видообразования у наездников, паразитирующих на этих мухах.
Разрушитель мифов:
На самом деле, ни о каком «видообразовании у наездников» тоже речи быть не может. Авторы исходной статьи (а также авторы других публикаций о «яблонной мухе») пишут о возможно начавшемся процессе видообразования. И никогда - в утвердительном наклонении. В утвердительном наклонении о «видообразовании у наездников» пишет только Александр Марков. Это хороший пример, когда эволюция доказывается не с помощью строгих эмпирических фактов, а с помощью соответствующих ударений и наклонений в нужных местах текста. Обратим внимание - фразы про: «видообразование», «хорошо задокументированное согласованное видообразование», «быстрое видообразование», «появление нового вида», «видообразование у мух и наездников» - эти фразы буквально рассыпаны по всей заметке А.В. Маркова.
А на самом деле, из того, что некая муха перешла на новое растение-хозяин (а за ней последовал и её наездник), и в результате, эти насекомые частично (!) перестали скрещиваться со старыми популяциями... из этих фактов еще совсем не следует, что видообразование теперь неизбежно «пойдет по накатанной дороге». Предположение о «накатанной дороге», на самом деле, следует из постулатов (самой) теории эволюции. Это её постулат (теории эволюции) - если произойдет ограничение скрещивания, то популяции со временем начнут все больше расходиться друг с другом, и в конце концов, образуют разные виды. А потом образуются разные роды, семейства, отряды и классы... Но постулаты необходимо доказывать фактами. А не наоборот, опираться на теоретические постулаты, делая прогнозы о будущем мух и наездников. Постулат о «видообразовании из-за возникновения изоляции» - до сих пор, вроде бы, не имеет строгих подтверждений. А вот строго опровергающие примеры уже установлены. И ниже я приведу целый ряд таких фактических опровержений. Но сейчас мы занимаемся мухами.
А.В. Марков:
...Мухи Rhagoletis pomonella живут в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника...
...до сих пор откладывала яйца только на плоды боярышника...
Разрушитель мифов:
Нет, это не так. Марков просто не знает, что авторы пишут только о тех популяциях мух Rhagoletis pomonella, которые, собственно, жили на боярышнике, причем только в восточной части США. А в целом для биологического вида Rhagoletis pomonella, их личинки развиваются отнюдь не только на боярышнике. Личинки мух Rhagoletis pomonella сегодня развиваются (Bush, 1966):
1. На домашней яблоне (Malus domestica).
2. На ягодной яблоне (Malus baccata). Это тоже интродуцированный вид, сибирская дикая яблоня.
3. На видах рода Боярышник (Crataegus).[57]
4. На двух видах рода Арония (Aronia): арония красная (A. arbutifolia) и арония черноплодная (A. melanocarpa).
5. На ирге (Amelanchier bartramiana).
6. На кизильнике (Cotoneaster).[58]
7. На разных видах рода Слива (Prunus). Например, на двух видах диких слив (P. angustifolia Marsh., P. umbellata Ell.), на культурной сливе и на персиках (Prunus persica).
8. На в ишне (Shervis et al., 1970).
9. Возможно, еще и на других растениях. Например, на груше, айве, абрикосе, черешне. Эти растения тоже указываются, как объекты паразитирования Rhagoletis pomonella в некоторых интернет-источниках, но оригинальных научных публикаций на эту тему я не нашел.
Кроме того, Марков не знает, что мухи Rhagoletis pomonella перешли на яблоки отнюдь не только в США, но еще и в Мексике (Bush, 1966). И этот переход был, скорее всего, совершенно независимым.
Наконец, данный вид мухи с готовностью атакует еще и другой вид яблони - ягодную яблоню (M. baccata), после того как её тоже завезли в Америку.
Очевидно, что это весьма важная информация. Потому что если брать только слова Маркова, то получается сильный пример, серьезно намекающий на эволюцию:
«Мухи раньше жили только на боярышнике, но в какой-то момент что-то случилось, и они перешли на яблони».
А если знать всю информацию, то окажется, что боярышник и яблоня - это лишь два растения из возможного списка тех растений, которые может использовать муха Rhagoletis pomonella для своих личинок. Причём: 1) переход на интересующие нас домашние яблони произошел независимо два раза, 2) муха так же перешла и на другой вид яблони - дикую ягодную яблоню.
Естественно, возникает вопрос - а эволюция ли это вообще? Или же эти мухи всегда так умели, то есть, всегда имели возможность питаться на яблонях? К этому вопросу мы еще вернёмся ниже.
А.В. Марков:
...За прошедшие с тех пор полтора века боярышниковая и яблоневая формы мух стали так сильно различаться по своему поведению, брачным предпочтениям и генетике, что сегодня их уже вполне можно считать двумя разными видами.
Разрушитель мифов:
И опять палеонтолог Марков бежит впереди паровоза. Он очень торопится сделать новую пищевую расу - именно новым видом. И это правильно. Ведь верующие в Дарвина так жаждут узреть настоящее чудо… пардон, наблюдаемую эволюцию. Поэтому палеонтолог Марков объявляет яблоневую расу мухи - новым видом, несмотря на то, что специалисты в данной области (зоологи, энтомологи, систематики, специалисты по роду Rhagoletis) пришли к выводу, что эту муху следует считать именно пищевой расой.
Вот, например, что пишут авторы одной из статей на эту тему (Filchak et al., 2000) прямо в резюме своей работы:
Боярышник (Crataegus spp.) - это природное растение-хозяин для R. pomonella в Северной Америке5. Но в середине 1800-ых, новая популяция сформировалась на ввезённой культурной яблоне (Malus pumila)4,5. Недавние исследования6-10 присвоили яблонным мухам статус «пищевой» расы [host race], как потенциально начинающемуся виду, частично изолированной от боярышниковых мух, вследствие адаптаций, связанных с растением-хозяином.
А вот соответствующие ссылки [6-10], на которые опираются авторы:
6. Prokopy, R.J., Diehl, S.R. & Cooley, S.S. Behavioral evidence for host races in Rhagoletis pomonella flies. Oecologia 76, 138–147 (1988).
7. Feder, J.L., Chilcote, C.A. & Bush, G.L. Genetic differentiation between sympatric host races of Rhagoletis pomonella. Nature 336, 61–64 (1988).
8. McPheron, B.A., Smith, D.C. & Berlocher, S.H. Genetic differences between Rhagoletis pomonella host races. Nature 336, 64–66 (1988).
9. Feder, J.L. et al. Host fidelity is an effective pre-mating barrier between sympatric races of the apple maggot fly. Proc. Natl Acad. Sci. USA 91, 7990–7994 (1994).
10. Feder, J.L. & Filchak, K.E. It’s about time: The evidence for host plant-mediated selection in the apple maggot fly, Rhagoletis pomonella, and its implications for fitness trade-offs in phytophagous insects. Ent. Exp. Appl. 91, 211–225 (1999).
В общем, повторюсь, никто из специалистов по данному таксону даже и не думает называть яблонную муху новым биологическим видом. Однако палеонтолог Марков, наверное, лучше знает, кого следует считать отдельным видом (а кого не считать) в комплексе видов плодовых мух Rhagoletis pomonella (комплекс очень близких видов-близнецов, относимых к группе R. pomonella).
Но зачем это делает Александр Марков? Зачем пишет: «...сегодня их уже вполне можно считать разными видами», игнорируя мнение специалистов? А для того, чтобы усилить эволюционное содержание текста.
Авторы многочисленных работ по яблонной мухе и так уже пишут в достаточно бравурном эволюционном стиле - о яблонных мухах, как примере «возможно начинающегося видообразования». Хотя «начинающееся видообразование» здесь следует не из фактов, а из теоретических посылок самого эволюционного учения, где используется постулат – «новые виды образуются в результате возникновения репродуктивной изоляции (той или иной природы) между популяциями». Например, в работе (Dres & Mallet, 2002) «пищевые» расы рассматриваются как переходный этап в процессе видообразования - где-то между простым полиморфизмом в популяциях и формированием нового, «хорошего» вида.
На самом же деле, наблюдаемым фактом является существование многих видов насекомых-фитофагов[59] в виде нескольких пищевых рас (но в рамках одного и того же биологического вида). Очень много насекомых-фитофагов живут не строго на одном виде растения, а на нескольких (обычно близких). Кроме того, наблюдаемым фактом является периодический переход тех или иных насекомых-фитофагов на новые виды подходящих для них растений-хозяев. Опубликованные примеры некоторых таких переходов приведены еще в работах Шапошникова (см. выше). И если в результате питания на разных кормовых растениях у насекомых возникает частичная репродуктивная изоляция (то есть, возникают частично изолированные популяции), то в этом случае биологи и называют такие частично изолированные популяции, специализированные к конкретному хозяину, «пищевыми» расами (host races). Причем это явление в природе имеет весьма распространенный характер. Из работы (Diegisser et al., 2007):
Большинство растительноядных насекомых - высоко видоспецифичны, питаясь только на нескольких видах растений-хозяев (Bernays & Graham, 1988; Jaenike, 1990). Если кормовая специализация или смена кормового растения у таких видов (олигофагов) приводит к частичной репродуктивной изоляции между внутривидовыми популяциями, то применяется термин «host race» (Diehl & Bush, 1984). Кормовые расы были установлены в разнообразных группах насекомых (например, Bush, 1969; Katakura & Hosogai, 1994; Sezer & Butlin, 1998; Via, 1999; Emelianov et al., 2001; Diegisser et al., 2004), показывая, что видообразование через использование различных растений-хозяев может быть общим феноменом среди насекомых фитофагов (Dres & Mallet, 2002).
Интересно, что и здесь авторы не смогли избавиться от желания теоретически порассуждать на тему эволюции. Сначала они озвучили фактический материал - что разные виды насекомых (из разнообразных групп) могут существовать в виде нескольких пищевых рас (биологический факт). Но не удержались на простой констатации этого факта, а перешли к рассуждениям о значении этого факта для «единственно верного учения».
Таким образом, мы видим, что профессиональные энтомологи (специалисты по конкретным таксонам насекомых) тоже не могут удержаться в рамках строгой научности, постоянно «соскакивая» на рассуждения о «возможно начинающемся видообразовании». Однако А.В. Маркову даже таких рассуждений (о возможности) мало. Он решил, что энтомологи проявляют непростительную нерешительность в «деле эволюции». И «доделывает» за энтомологов их работу, объявляя яблонную муху новым видом. Конечно, он ошибся. Ведь энтомологи (специалисты по таксону Rhagoletis)не зря едят свой хлеб, и в советах палеонтолога, специалиста по ископаемым морским ежам, вряд ли нуждаются. Ниже я объясню, почему яблонную муху нельзя считать новым видом.
А.В. Марков:
...В принципе для зарождения «яблоневого» вида было достаточно одной мутантной самки, которой запах яблок понравился больше, чем запах боярышника, и которая передала эту особенность своему потомству...
Разрушитель мифов:
Эта фраза, в принципе, правильная. Но Марков озвучивает только данную версию (о мутации самки) и больше никакую. Понятно, почему он озвучивает именно эту версию. Потому что именно эта версия идеологически соответствует «единственно верному учению», где как известно, правят бал именно случайные мутации+естественный отбор.
Но ведь есть и другие возможные варианты. Например:
1. Реализация уже имевшейся внутривидовой изменчивости. Допустим, где-то в генофонде (среди разных популяций) этого вида уже имелся какой-нибудь редкий аллель (возможно, рецессивный), который вызывает у самок влечение именно к созревающим яблокам. И с ввезением яблонь в США этот аллель просто нашел себе применение. В какой-то момент в какой-то самке встретились сразу два этих редких (рецессивных) аллеля. Вот эта самка и полетела к первому встречному яблоку. Правда, возникает вопрос - а что же этот аллель делал в диких популяциях данных мух до завезения в Америку культурных яблонь?
На это можно ответить, например, таким образом - сохранялся «до лучших времен». Действительно, очень интересным фактом является то, что мухи Rhagoletis pomonella сегодня (как это ни странно) не выводят своих личинок на диких видах американских яблонь. Этот парадокс специалисты по таксону Rhagoletis pomonella объясняют предположительным несоответствием времени созревания диких американских яблонь - времени развития личинок Rhagoletis pomonella (Bush, 1966:462). Яблоки диких видов американских яблонь созревают слишком поздно, чтобы личинка мухи успела пройти в ней все необходимые стадии развития до наступления осенне-зимних холодов. И поэтому муха и не атакует дикие американские яблони. Но возможно, так было не всегда? Возможно, раньше или ареал диких яблонь простирался дальше на юг, или климат был теплее. В этом случае, аллель «привлекательности запаха яблок» вполне находил бы себе применение (хотя бы иногда).
Эту версию (с уже имеющимся аллелем) я выдумал прямо на ходу. Но ведь такая возможность, действительно, не исключена. И если это так, то «строго установленный пример эволюции» сразу же обесценивается. Ведь в этом случае не было никакой эволюции - подходящий признак у мух данного вида уже имелся. Итак (допустим) каких-то мух этого вида всегда привлекал запах яблок. Но до ввезения культурных яблонь этот признак не находил себе применения, потому что дикие местные яблоки созревали тогда, когда у мух уже пропадало всякое желание размножаться. Однако с ввезением в Америку культурных яблонь (которые созревают гораздо раньше, даже раньше созревания боярышника) этот редкий аллель нашел себе применение. Дополнительным фактом, который можно истолковать в пользу этой версии, является то, что Rhagoletis pomonella столь же успешно перешла на завезенные культурные яблони еще и в Мексике. Дикие виды американских яблонь в Мексике вообще не растут, поэтому тот гипотетический аллель (существование которого мы предположили), раньше не находил себе применения и в мексиканских популяциях Rhagoletis pomonella. Но с «приездом» яблонь в Мексику этот аллель тоже быстро нашел себе применение. Чем и объясняется практически синхронное (в исторических рамках) независимое освоение яблонь этими мухами в США и в Мексике.
Очевидно, что предложенное объяснение является одним из самых простых, лежащих прямо на поверхности. Почему же такая версия никем не озвучивается? Потому что такие банальные версии никому не интересны. А интересны примеры именно «наблюдаемой эволюции». И чтобы там обязательно звучала мысль про «единственного случайного мутанта». Именно так будет идеологически правильно. Вот из яблонных мух и делают мутантов.
2. Версия скрытой изменчивости. Возможно, в геноме данного вида существует некая «база данных» потенциально привлекательных запахов. В которой периодически происходят переключения (у отдельных представителей вида). Хотя бы просто для того, чтобы «посмотреть, что в мире творится» (то есть, для проверки появления новых подходящих видов растений-хозяев).
Например, у человека имеется примерно 400 функциональных генов, определяющих рецепторы запахов, и еще 600 неработающих, кандидатов в «псевдогены» (вероятно, «сломавшихся»). И еще есть 34 рецептора феромонов, из которых работает только один, да и то под вопросом. И есть еще 41 рецептор горького вкуса, из которых работает только половина.
Возможно, что-то похожее есть и у обсуждаемой нами мухи? Только эта «база данных» у мухи реально работает. И просто время от времени генетически «переключаются» соответствующие предпочтения в этой «базе данных» возможных привлекательных запахов.
И может быть, не только у мух? Может быть, этим объясняются периодические переходы и других видов насекомых-фитофагов с одного подходящего растения на другое? Возможно, это широко распространенное среди насекомых явление (существование в форме разных пищевых рас) просто закодировано в геноме таких видов? И реализуется с помощью периодических переключений в определенной генетической «базе данных» соответствующих запахов, привлекающих насекомое к тем растениям, которые потенциально может использовать данный вид?
В связи с этой мыслью интересен тот факт, что изменение запаховых предпочтений у разных рас мухи Rhagoletis pomonella - выглядит именно как переключение. Например, яблонная раса мухи Rhagoletis pomonella привлекается именно запахом яблок, но не боярышника. А боярышниковая раса Rhagoletis pomonella, наоборот, привлекается запахом боярышника, но не запахом созревающих яблок (Linn et al., 2003). Возникает вопрос - а собственно, почему это так? Ведь оба этих фрукта подходят для мухи данного вида примерно в одинаковой степени (см. ниже). В связи с этим, разве мухе выгоден такой добровольный отказ от части её же кормовой базы? Более того, ряд установленных фактов серьезно свидетельствует в пользу того, что мухи Rhagoletis pomonella не только не привлекаются запахом «неродных» для них фруктов, но этот запах (неродных фруктов) действует вообще отталкивающе на этих мух, вызывая у них поведение избегания (Linn et al., 2003; Linn et al., 2004). Это как вообще понимать? С какой стати плодовая муха, гипотетически участвуя в «напряженной борьбе за существование» (в том числе, и за пищевые ресурсы для своих личинок), стремится откладывать яйца исключительно во фрукты только одного вида растений, игнорируя и даже активно «воротя нос» (!) от всех других подходящих для неё фруктов?!
Тем не менее, мы наблюдаем то, что наблюдаем:
1). Яблонная раса мух «любит» только запах яблок. То есть, только запах тех фруктов, которыми эти мухи питались на личиночной стадии. И не любит ни запах боярышника, ни запах кизила и т.п.
2). А вот боярышниковая раса мух любит запах именно боярышника (то есть, именно тот фрукт, которым питались личинки). И не реагирует положительно ни на запах яблок, ни на запах кизила.
3). А муха еще «неопределенной таксономической ориентации» (то ли раса, то ли вид), питающаяся на одном из видов кизила (Cornus florida) любит... правильно, именно запах ягод кизила и только их. И не любит ни запах яблок, ни запах боярышника.
Это настолько удивительно, что напрашивается мысль, а наследственны ли вообще пищевые предпочтения этих плодовых мух? Или же эти предпочтения имеют приобретенный характер? То есть, насекомое просто запоминает тот запах, которым оно питалось на стадии личинки, и далее, уже на взрослой стадии, стремится именно к этому запаху?
Вообще, нахождение насекомыми своей потенциальной пищи - это сложный процесс, включающий в себя использование: 1) определенных запаховых меток (на дальне-средней дистанции), 2) визуальных сигналов (на ближне-средней дистанции), 3) тактильных сигналов при физическом контакте (Linn et al., 2004). А так же задействование индивидуального опыта и результатов научения насекомого (Linn et al., 2004). То есть, приобретенные эффекты в пищевых, запаховых или охотничьих препочтениях, действительно, вполне могут иметь место.
Однако некоторые результаты, которые были получены в исследованиях, посвященных изучению именно запаховых предпочтений обсуждаемых нами мух, крайне серьезно свидетельствуют в пользу всё-таки врождённости этих предпочтений. Например, сохранение предпочтений к «родному» (для расы) запаху фруктов, несмотря на выращивание личинок в «неродном» фрукте (Linn et al., 2003). Или же специфическая реакция гибридов первого поколения на запахи, которую тоже вряд ли можно объяснить приобретенным характером пищевых предпочтений (Linn et al., 2004).
Итак, запаховые предпочтения именно к «родным» фруктам (и только к ним!) у разных рас Phagoletis pomonella имеют почти наверняка врожденный характер. Однако почему-то эти предпочтения очень напоминают действие переключателя – «включается» привлекательность именно определенного запаха (именно «родного» фрукта), но одновременно «выключается» привлекательность запахов всех других фруктов, в которых развитие личинок этого вида мух тоже вполне возможно.
В связи с этим напрашивается вопрос - может быть, всё-таки эти предпочтения имеют хотя бы полу-приобретенный характер?
Например, при установлении генетической метки привлекательности на определенный запах имеют место какие-то эпигенетические механизмы? Эпигенетические механизмы сейчас модно упоминать. И это правильно, поскольку эпигенетические механизмы, действительно, имеют место в живой природе. Авторы соответствующих работ обсуждают некоторые такие возможности в отношении разбираемых нами мух. Например, возможность пресловутых материнских эффектов («maternal effects»). И даже вроде бы частично обнаруживают такие эффекты в качестве одного из результатов своих исследований. Например, было обнаружено, что гибриды первого поколения немного больше предпочитают те фрукты, которыми питались матери этих гибридов, но не отцы (Linn et al., 2004).
Однако Александр Марков обходит полным молчанием все перечисленные выше возможные механизмы «включения» привлекательности запаха яблок для новой расы мух Rhagoletis pomonella, лаконично сосредотачиваясь только на версии «одной мутантной самки».
Интересно, что со стороны такое «переключение», действительно, может выглядеть именно как «случайная мутация». А на самом деле, это может быть отнюдь не «случайная мутация», а реализация уже имевшихся скрытых возможностей (конкретного вида), о которых говорил еще Шапошников. И реализация этих скрытых возможностей могла осуществляться по тем гипотетическим механизмам, которые я уже перечислил выше.
Например, у человека, при попадании в организм инфекции, в соответствующих иммунных клетках происходит быстрый «гипермутагенез», направленный на получение антител, наиболее подходящих против этой инфекции. То есть, осуществляется процесс, аналогичный подбору компьютерного пароля (компьютерными хакерами). Этот процесс чисто внешне тоже напоминает «случайные мутации». Но никто не называет этот процесс эволюцией. Все понимают, что это один из специальных приспособительных механизмов организма.
Однако если какие-то аналогичные процессы (допустим, случайные генетические переключения) осуществляются в уже имеющейся «базе данных» привлекательных запахов мухи Rhagoletis pomonella, то почему это следует называть уже эволюцией, а не считать тоже каким-то специальным биологическим механизмом? Например, характерным для насекомых-фитофагов? Может быть, это такой специальный биологический механизм, который и приводит к возникновению столь характерного для этих насекомых феномена «пищевых» рас (или даже видов-близнецов)?
Но если предположение о «скрытой изменчивости, уже заложенной в данном виде», действительно соответствует истине, то отсюда следует, что у тех же мух Rhagoletis pomonella - возможность адаптации к разным привлекательным запахам ограничена. Ведь «случайные» мутации здесь, на самом деле, являются «псевдослучайными». То есть, на самом деле, реализуются уже имеющиеся возможности, заложенные в генетической «базе данных» этого биологического вида.
Отсюда вытекает проверяемое следствие - если это, действительно, лишь реализация скрытой изменчивости (о которой я уже подробно писал выше), тогда мы должны наблюдать у этой мухи «приспособление» лишь к определенному кругу запахов. И чуть ниже мы попробуем выяснить - справедливо ли это для мухи Rhagoletis pomonella.
Наконец (еще раз), не исключена возможность, что переключение предпочтений к определенным запахам вообще имеет эпигенетический характер (то есть, «полуприобретенный»).
Итак, возможных причин изменения запаховых предпочтений мух - немного больше, чем та одинокая версия (о «случайной мутации единственной самки»), которую озвучивает Александр Марков. Но эта единственная озвученная версия - самая идеологически правильная. Вот Марков её и озвучивает. Или он уже настолько привык рассуждать именно в рамках «эволюции под действием случайных мутаций и естественного отбора», что какие-то другие механизмы даже не может себе представить.
В заключение поправлю еще несколько незначительных неточностей, уже не имеющих принципиального характера:
А.В. Марков:
Со своей стороны, наездники стимулируют видообразование у мух, заставляя их переходить на новые растения...
Разрушитель мифов:
Это весьма вольное размышление автора, но опять вынесенное в резюме в утвердительном наклонении. На самом деле, даже если бы не было никаких наездников, то переход мух на новое растение-хозяин всё равно теоретически мог бы произойти, если новое растение достаточно подходит для данных мух, и если у этих мух произошла смена прежних пищевых предпочтений на новые. После этого, перешедшие мухи спокойно продолжали бы жить на яблоках, даже если бы никаких наездников вообще не существовало в природе. Конечно, уменьшение давления паразитов при переходе их жертв на новые растения, в принципе, возможно. Более того, уже зарегистрирован целый ряд таких случаев (Feder, 1995). Однако (еще раз) для успешного перехода насекомого-фитофага на новое для него (подходящее) растение - вполне достаточно только одной лишь смены пищевых предпочтений. Без всяких «наездников, заставляющих мух куда-то переходить».
А.В. Марков:
Позднее энтомологи обнаружили еще два вида Rhagoletis, которые размножаются на чернике и снежноягоднике.
Разрушитель мифов:
Не очень понятно, что здесь имеет в виду Марков под словом «позднее». Муха из группы Rhagoletis pomonella, паразитирующая на снежноягоднике - это Rhagolethis zephyria Snow 1894. Для тех, кто не в курсе – «Snow» - это фамилия того человека, который первым описал данный вид. А 1894 - это год описания. Итак, описание вида в конце 19 века - это как (с точки зрения Маркова) достаточно поздно? И по сравнению с чем? А муха, размножающаяся на «чернике» - это Rhagolethis mendax Curran 1932. Может быть, именно эта дата имелась в виду под «позднее»? Но дело в том, что энтомологи обнаружили мух, крайне похожих на «яблонную муху», но поражающих разные виды голубики («черники» по Маркову), снежноягодника, кизила и др. - еще в конце 19, начале 20 века. Просто долгое время эти мухи считались не разными видами, а либо отдельными расами вида Rhagoletis pomonella, либо подвидами. И в отношении некоторых мух этот взгляд господствовал вплоть до середины 20 века. Например, в отношении Rhagoletis mendax, которая морфологически очень близка к Rhagoletis pomonella, большинство специалистов считали её лишь подвидом (или расой) вида Rhagoletis pomonella вплоть до 60-х годов 20 века (Bush, 1966).
Кроме того, когда Марков пишет, что мухи «размножаются на чернике» (имея в виду, наверное, Rhagoletis mendax?) он еще раз повторяет свою предыдущую ошибку, которую уже сделал в отношении кормовых растений яблонной мухи. На самом деле, Rhagolethis mendax, так же как и «яблонная муха», питается отнюдь не только на одном-двух растениях. «Черничная муха» паразитирует на широком круге ягодных растений, относящихся к семейству вересковые - на разных видах голубики, на клюкве, на бруснике, на видах рода гейлюссакия (Gaylussacia), на гаультерии (Gaultheria) и на других ягодных растениях этого семейства (Bush, 1966). И лишь Rhagolethis zephyria, действительно, питается на растениях только одного рода (снежноягодник, Symphoricarpos), которых в Америке насчитывается примерно полтора десятка видов (Bush, 1966).
Наконец, непонятно, почему Марков говорит только о «еще двух позднее обнаруженных видах Rhagoletis». И кого он, собственно, подразумевает под «видами Rhagoletis». Если он подразумевает виды рода Rhagoletis, то таких видов, на самом деле, не три, а несколько десятков (больше пятидесяти). Если же Марков подразумевает только близкие виды из группы Rhagoletis pomonella (комплекс видов-близнецов группы Rhagoletis pomonella внутри рода Rhagoletis), тогда Марков всё равно забыл, по крайней мере, еще один вид (из этой же группы видов-близнецов) - Rhagoletis cornivora Bush 1966, который размножается на разных видах кизила (род Cornus). Этой мухе (размножающейся на кизиле) энтомологи, действительно, присвоили статус отдельного вида позднее (уже во второй половине 20 века). Понятно, почему Марков «забыл» этот вид. Потому что в статье, которую он комментировал, фигурировали только три вида из группы Rhagoletis pomonella (R. pomonella, R. mendax и R. zephyria). Вот Марков и пишет только о них.
А.В. Марков:
Со снежноягодниковым видом исследователи пока не разобрались (не сумели добыть материал в достаточных количествах), а по остальным трем видам картина получилась следующая.
Разрушитель мифов:
Опять те же ошибки. Марков с одной стороны ошибается, думая, что в группе Rhagoletis pomonella имеются только те виды, которые он уже озвучил. А с другой стороны ошибается, считая новую (яблоневую) расу отдельным видом.
А.В. Марков:
Интересно, что яблоки являются менее благоприятной средой обитания для личинок мух Rhagoletis, чем плоды боярышника. В боярышнике личинки выходят из половины отложенных мухой яиц, а в яблоке — только из четверти.
Разрушитель мифов:
Здесь похвально, что Александр Марков попытался почитать про яблонных мух не только ту статью, о которой он написал свою заметку, но и еще одну научную публикацию, на которую в этой работе была ссылка.[60]
Но плохо здесь то, что Марков прочитал только одну (дополнительную) статью. Поэтому Марков и пишет, что «в боярышнике личинки выходят из половины отложенных мухой яиц, а в яблоке - только из четверти».
А на самом деле, речь (в прочитанной Марковым дополнительной статье) во-первых, идет вообще не о том,какой процент личинок выходит из отложенных яиц. А об общей выживаемости этих мух от стадии яйца до стадии куколки через стадию личинки.
Во-вторых, если бы Марков почитал еще какие-нибудь статьи (кроме той, которую он прочитал), то он бы узнал, что это только в одном исследовании получилось так, как он пишет - что выживание мух от стадии яиц до стадии куколок (через личиночную стадию) составило 52% для личинок, питающихся в боярышнике, и только 27% для личинок, питающихся в яблоках (Prokopy et al., 1988). А вот в другом исследовании выживаемость личинок составила примерно 53% для личинок, питающихся в боярышнике, и уже 43% для личинок, питающихся в яблоках (Feder et al., 1995). Более того, Марков бы узнал (если бы больше почитал), чем обусловлена эта разница в выживаемости личинок в яблоках, установленная в разных исследованиях. Cкорее всего, выживаемость личинок зависит от конкретного сорта того яблока, в которое попадает личинка. Максимальная выживаемость личинок наблюдалась в более сладких и более ранних сортах яблок. Например, еще в одном (уже третьем) исследовании, выживаемость личинок в ранних сортах яблок составила 46% (т.е. лишь ненамного меньше, чем в боярышнике), по сравнению с поздними сортами яблок, где выживаемость личинок составила всего 25% (Cameron & Morrison, 1977). Теперь если вспомнить о том, что и разных боярышников в Северной Америке - тоже далеко не один вид, то наверное, правомерно будет предположить, что выживаемость личинок мух Rhagoletis pomonella в разных видах боярышников тоже будет варьировать в той или иной степени. Таким образом, разница между выживаемостью личинок R. pomonella в яблоках и в боярышнике во многих случаях совсем не такая драматичная, как пишет Марков, и сильно зависит от того, насколько тот или иной сорт яблока подходит для данной личинки.
А.В. Марков:
Таким образом, в переходе с боярышника на яблоки были не только плюсы, но и минусы. Это можно рассматривать как косвенный аргумент в пользу того, что паразиты были важным фактором, способствовавшим видообразованию у мух. Если бы не паразиты, от которых яблоки дают лучшую защиту, естественный отбор не поддержал бы переход мух на менее подходящие для них плоды.
Разрушитель мифов:
Здесь в одном абзаце у Маркова собраны сразу три неточности:
Во-первых, как я уже только что объяснил выше, «плюсы и минусы» имеются не столько в отношении перехода мух с боярышника на яблоки в целом, сколько в отношении лишь отдельных сортов культурных яблонь.
Во-вторых, даже если бы вообще не было никаких паразитов, то переход на яблоки вполне мог бы осуществиться. Причем даже на сравнительно неподходящие сорта. Если яблоки хотя бы немного подходят для этих мух, и если эти мухи имеют запаховые предпочтения именно к яблокам, но не к боярышнику (а исследования показали, что так оно и есть), то этих условий уже достаточно, чтобы мухи перешли на яблони и образовали там новую пищевую расу. Даже если выживаемость личинок будет составлять всего половину от такой же выживаемости личинок в боярышнике, то всё равно, существование новой кормовой расы теоретически возможно. Потому что, используя яблони в качестве нового растения-хозяина, такая раса мух вообще уходит от конкуренции с исходной предковой расой (и не только). Например, яблоневая раса получает больше возможностей пристроить своих личинок в плоды, свободные от других насекомых (см. ниже про конкуренцию личинок насекомых в плодах боярышника).
Но даже если наличие свободных плодов, на самом деле, не является лимитирующим фактором для этого вида, то новая раса яблоневых мух всё равно может существовать в природе самым спокойным образом – просто в виде популяций с меньшей плотностью особей (чем у боярышниковых мух). Даже если плотность популяций новой расы будет ниже, чем у боярышниковой расы в два, три, четыре (или даже в десять) раз, то теоретически, такие популяции всё еще могут спокойно выживать в природе. Эту теоретическую возможность следовало бы учесть, и проверить в соответствующих исследованиях, прежде чем заявлять, что «естественный отбор не поддержал бы»…
В-третьих, как я уже говорил выше - ни о каком «видообразовании мух» здесь не может быть и речи. В крайнем случае, речь здесь может идти лишь о «возможно начинающемся процессе видообразования».
Таким образом, упомянутый Марковым естественный отбор здесь вообще не причём. Если бы пресловутый «естественный отбор не поддержал бы переход мух на менее подходящие для них плоды» (как пишет Марков), то яблоневые мухи ведь не умеют читать. Поэтому они бы всё равно не узнали об этом. И так и продолжали бы жить на яблоках, несмотря на то, что «процент вышедших из яиц личинок в яблоках в два раза ниже, чем в боярышнике». Ведь у них просто нет конкурентов на яблонях. И еще потому, что на самом деле, «плохой выход личинок из яиц в яблоках» это просто результат неточности изложения материала (см. выше).
КОНЕЦ СКАЗКИ
Ну а теперь подошло время разобрать завершение новой эволюционной легенды, которое сочинил уже Михаил Гельфанд. Выделенное жирным шрифтом в его тексте - это тоже (извиняюсь за выражение) враньё.
М.С. Гельфанд:
Это такой стандартный запрос учёным: «Покажите нам, как один вид превратился в другой». Самый красивый пример – это история с яблоневой мухой в северной Америке.
Разрушитель мифов:
Отрадно, что Гельфанд (в отличие от Маркова) нигде не называет новую пищевую расу мух – «новым видом». Гельфанд вообще никак не озвучивает таксономический ранг яблонных мух. Однако всё равно продолжает сомнительные традиции Маркова, потому что рассказывает именно о яблонных мухах в контексте примера «превращения одного вида в другой».
М.С. Гельфанд:
Там не было яблонь, их туда завезли в середине XIX века. Соответственно, часть популяции плодовой мухи, которая жила на боярышнике, переселилась на яблони, жила в яблоках.
Разрушитель мифов:
Здесь Гельфанд перепутал. Это такое подобие «глухого телефона», когда степень искажения фактов усиливается при переходе от одного рассказчика к другому. На самом деле, не яблони завезли в середине 19 века, а мухи Rhagoletis pomonella перешли на яблони в середине 19 века. Сами же яблони «приехали» в Новую Англию в 17 веке. А в Мексику домашние яблони «приехали» еще на сто лет раньше. В начале 16 века. И на эти (мексиканские) яблони мухи Rhagoletis pomonella тоже перешли, правда, точно неизвестно когда. Причем этот переход был, скорее всего, совершенно независимым от обсуждаемого перехода этих мух на яблоки в Новой Англии (Bush, 1966).
М.С. Гельфанд:
С точки зрения конкретной мухи, это на самом деле плохо. Те мухи, которые живут на яблонях, у них меньше потомства, личинки хуже выживают.
Разрушитель мифов:
Выше я уже объяснил, что это может быть не так. В зависимости от конкретного сорта яблок разница может быть совсем небольшой. И самих свободных яблок (для личинок) может быть больше.
М.С. Гельфанд:
Тем самым, мухи хуже живут с точки зрения еды, но у них лучше защита от хищника.
Разрушитель мифов:
То же самое. Не во всех сортах яблок мухи живут хуже с точки зрения еды. Да и термин «хищник», употребленный биологом в отношении паразитической осы, немного режет слух.
М.С. Гельфанд:
Самая полная вершина этой картинки была, когда оказалось, что образовалась раса наездников с более длинными яйцекладами, которые специально паразитируют на тех мухах, которые живут в яблонях.
Дата добавления: 2018-09-20; просмотров: 396; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!