Цикл Кребса – центральный метаболический путь.
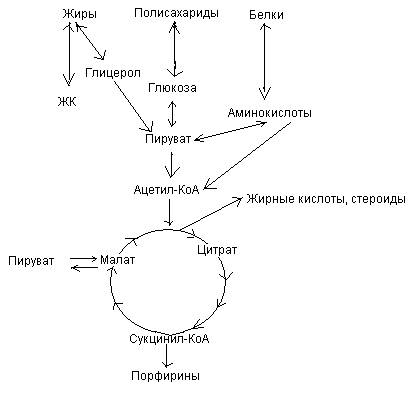
| Цикл Кребса является центральным метаболическим путем, т.к. 1) связывает в клетке процессы катаболизма и анаболизма (пример: образующийся оксалоацетат используется в процессах глюконеогенеза, синтеза заменымых АК, цитрат – в синтезе жирных кислот и т.д.) 2) объединяет белки, жиры, углеводы, в нем сходятся почти все метаболические пути 3) является общим путем окончательного окисления большей части органических молекул в форме ацетил-КоА, играющих роль клеточного «топлива» 4) является основным путем, обеспечивающим энергией большинство клеток. |
21. 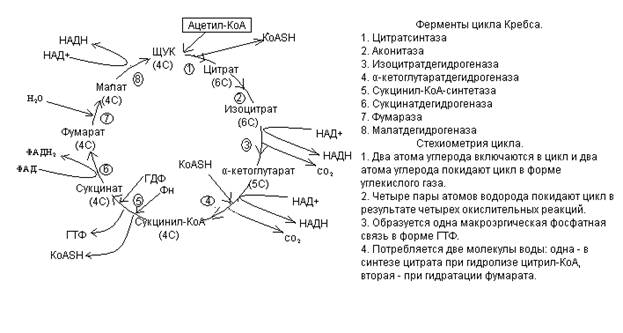
Энергетический баланс окисления ацетил~КоА: 1 ГТФ в ходе ЦТК + 3*3 АТФ (за счет НАД-переносчика) + 2 АТФ (за счет ФАД-переносчика) = 12 АТФ
22. См. вопрос 21.
Витамины, принимающие участие в работе цикла:
| Витамин | Кофермент в составе витамина |
| Тиамин | ТПФ |
| Никотиновая кислота | НАД |
| Рибофлавин | ФАД |
| Пантотеновая кислота | КоА |
Также для нормальной работы цикла необходимо витаминоподобное вещество – липоевая кислота.
Недостаток хотя бы одного из витаминов, используемых в синтезе небелковых компонентов ферментов цикла, сопровождается лактатным ацидозом и нарушением энергопродукции в клетке.
23. Катаболическая функция цикла Кребса:
1) конечный путь окисления большинства органических веществ
2) основной источник запасания энергии в клетке в форме АТФ
|
|
|
Кол-во АТФ, образуемых при окислении тирозина до фумарата: 3 АТФ (за счет НАД в результате остатка цикла) + 2 * 12 АТФ (т.к. фумарат содержит 4 атома углерода, то они элиминируются через два полных оборота цикла, а в каждом цикле образуется 12 АТФ) = 27 АТФ.
24. Кол-во АТФ, образуемых при окислении пролина до α-кетоглутарата: 3*2 АТФ (от НАД) + 2 АТФ (от ФАД) + ГТФ (в результате остатка цикла) + 2 * 12 АТФ (т.к. α-кетоглутарат содержит 5-1 атом углерода, то они элиминируются через два полных оборота цикла, а в каждом цикле образуется 12 АТФ) = 33 АТФ.
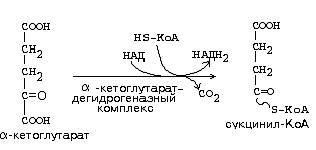
25. Кол-во АТФ, образуемых при окислении моля аспарагина до оксалоацетата: 2 * 12 АТФ (т.к. оксалоацетат содержит 4 атома углерода, то они элиминируются через два полных оборота цикла, а в каждом цикле образуется 12 АТФ) = 24 АТФ.
26. См. вопрос 24.
27. Кол-во АТФ, образуемых при окислении моля метионина до сукцинил-КоА: 1 ГТФ + 2 АТФ (за счет ФАД) + 3АТФ (за счет НАД) в результате остатка цикла + 2 * 12 АТФ (т.к. сукцинил-КоА содержит 4 атома углерода, то они элиминируются через два полных оборота цикла, а в каждом цикле образуется 12 АТФ) = 30 АТФ.
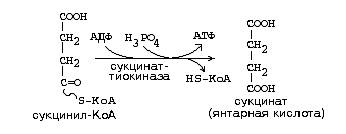
28. См. вопрос 21.

29. Реакции ЦТК, катализируемых дегидрогеназами:
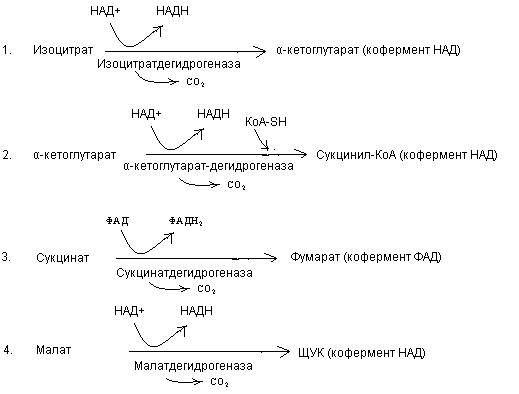
30. Окислительное фосфорилирование – синтез АТФ за счет энергии, выделяющейся при окислении водорода органических субстратов кислородом с участием дыхательной цепи.
|
|
|
Этапы (комплексы) дыхательной цепи:
I комплекс (НАДН: убихинон-оксидоредуктаза) – принимает электроны от митохондриального НАДН и транспортирует их на коэнзим Q. Протоны при этом транспортируются в ММП. Промежуточным акцептором и переносчиком протонов и электронов являются ФМН и железо-серные белки. Разделяет поток протонов и электронов.
II комплекс (Сукцинат: убихинон-оксидоредуктаза) – включает сукцинатдегидрогеназу, транспортирует электроны от сукцината на убихинон (КоQ) с образованием промежуточного ФАДН2и с участием железосерных белков.
III комплекс (Убихинол: цитохром с-оксидоредуктаза) – имеет в составе цитохромы b и c1 и железо-серные белки. Транспортирует электроны от КоQ через цитохромы b и c1 на цитохром c. Разделяет поток протонов и электронов (протоны транспортируются в ММП, а электроны следуют далее по цепи).
IV комплекс(Цитохромоксидаза – в составе - атомы меди и группы гема) – содержит цитохромы а и а3 и обладает свойствами фермента. Электроны транспортируются с III на IV комплекс с участием промежуточного подвижного переносчика цитохрома c. Транспортирует электроны с на молекулярный кислород. Способен перекачивать протоны из матрикса в ММП.
|
|
|
Субклеточная локализация: митохондрии (в основном, внутренняя мембрана, матрикс и ММП).
При недостатке кислорода процесс окислительного фосфорилирования прекращается. Механизм: НАД и ФАД в митохондриях могут быть регенерированы только при условии переноса электронов на молекулярный кислород. При недостатке кислорода переноса протонов и электронов из ЦТК на дыхательную цепь не происходит и процесс ингибируется.
31. См. вопрос 18.
Разобщители окислительного фосфорилирования – вещества, нарушающие систему сопряжения тканевого дыхания и фосфорилирования.
Могут обладать следующими свойствами:
а) увеличивают проницаемость внутренней мембраны для протонов и других катионов
б) являются альтернативными акцепторами электронов, транспортирующими электроны в обход одного или нескольких комплексов дыхательной цепи.
32. При увеличении концентрации АДФ потребление кислорода митохондриями возрастет, т.к. увеличится интенсивность катаболических реакций, направленных на синтез АТФ в результате дыхательного контроля.
Механизм. Потребление кислорода находится в прямо пропорциональной зависимости от потребности в АТФ. Когда увеличивается потребность в АТФ, т.е. когда возрастает соотношение АДФ/АТФ, адениловые нуклеотиды стимулируют по аллостерическому механизму метаболические процессы и поглощение кислорода возрастает.
|
|
|
33. Пункты фосфорилирования – участки в дыхательной цепи, где энергия транспорта электронов используется на генерацию протонного градиента, а затем в ходе фосфорилирования запасается в форме АТФ:
1-ый: между пиридинзависимой и флавинзависимой дегидрогеназами
2-ой: между цитохромами b и c1
3-ий: между цитохромами а и а3
Количество энергии, выделяемой в реакции переноса электронов по дыхательной цепи, можно определить по изменению редокс-потенциала (потенциала, образуемого переносимыми электронами), который пропорционален изменению свободной энергии системы ∆G0’.
34. Нет. НАДФН служит донором электронов в восстановительных биосинтезах и располагается преимущественно в цитозоле в отличие от НАДН, который окисляется кислородом дыхательной цепи с сопутствующим генерированием АТФ и располагается в матриксе митохондрий.
35. Пиридинзависимыесубстраты тканевого дыхания: изоцитрат, малат, α-кетоглутарат, β-гидроксиацил-КоА, гидроксибутират, пируват, α-кетоглутарат, пролин.
Место включения в дыхательную цепь: I комплекс (НАДН: убихинон-оксидоредуктаза).
36. При окислении сукцината в фумарат коэффициент фосфорилирования P/O=2.
37. С первым комплексом дыхательной цепи связаны реакции окисления: 1) изоцитрата в α-кетоглутарат, 2) α-кетоглутарата в сукцинил-КоА, 3) малата в оксалоацетат (ЩУК).
При этом можно получить 3 АТФ на каждую реакцию окисления.
38. При окислении изоцитрата в α-кетоглутарат можно получить три молекулы АТФ. Место включения α-кетоглутарата в дыхательную цепь - I комплекс (НАДН: убихинон-оксидоредуктаза).
39. При окислении α-кетоглутарата в сукцинил-КоА коэффициент фосфорилирования Р/О=3. См. вопрос 38.
40. При окислении яблочной кислоты (малата) в ЩУК P/O=3. Участки сопряжения транспорта электронов и фосфорилирования см. вопрос 33.
41. При окислении сукцината в фумарат образуется 2 АТФ.
42. При блокаде дыхательной цепи производными барбитуровой кислоты (ротеноном, аминобарбиталом (амиталом) I комплекс (ФМН и железо-серные белки) будет находиться
в восстановленном состоянии, остальные переносчики – в окисленном.
43. Малонат – конкурентный ингибитор сукцинатдегидрогеназы (II комплекс), поэтому в митохондриях будет накапливаться сукцинат.
44. При блокаде дыхательной цепи цианидами (а также СО, азидом натрия) все комплексы дыхательной цепи, включая цитохромоксидазу (место действия ингибитора), будут в восстановленном состоянии, т.к. ингибитор препятствует транспорту электронов на конечный акцептор – кислород.
45. См. вопрос 42.
46. При блокаде цепи антимицином А блокируется окисление цитохрома b, поэтому ФМН, КоQ, цит b будут восстановленными, цит с1, с, а и а3 будут окисленными.
47. СО ингибирует цитохромоксидазу дыхательной цепи и препятствует переносу электронов на конечный акцептор – кислород. Также угарный газ связывается с гемоглобином эритроцитов, образуя устойчивое соединение – карбоксигемоглобин. При этом из-за высокого сродства к СО гемоглобин теряет способность связывать кислород и дыхательная цепь, работающая только при наличие O2, перестает функционировать.
Дата добавления: 2018-08-06; просмотров: 359; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!