Первые реакции окисления ненасыщенных жирных кислот и роль изомераз
Расчет энергетического баланса β-окисления ненасыщенной жирной кислоты
При расчете количества АТФ, образуемого при β-окислении ненасыщенных жирных кислот необходимо учитывать:
· количество образуемого ацетил-SКоА – определяется обычным делением числа атомов углерода в жирной кислоте на 2.
· число циклов β-окисления. Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно подсчитать по формуле (n/2 -1), где n – число атомов углерода в кислоте.
· число двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество необразованных ФАДН2 соответствует числу двойных связей. Остальные реакции цикла идут без изменений.
· количество энергии АТФ, потраченной на активацию (всегда соответствует двуммакроэргическим связям).
Пример. Окисление линолевой кислоты
· так как число атомов углерода равно 18, то количество молекул ацетил-S-КоАравно 9. Последний поступает в ЦТК, при его окислении в одном обороте цикла образуется 3 молекулы НАДН (7,5 АТФ), 1 молекула ФАДН2 (1 АТФ) и 1 молекула ГТФ, что эквивалентно 10 молекулам АТФ. Итак, 9 молекул ацетил-SКоА обеспечат образование 9×10=90 молекул АТФ.
· исходя из формулы (n/2 - 1) число циклов β-окисления равно 8. В каждом цикле образуется 1 молекула ФАДН2 (1,5 АТФ) и 1 молекула НАДН (2,5 АТФ). Поступая в дыхательную цепь, в сумме они "дадут" 4 молекулы АТФ. При расчете получаем 8×4=32 молекулы АТФ.
· в кислоте имеются 2 двойные связи. Следовательно, в двух циклах β-окисления необразуется 2 молекулы ФАДН2, что равноценно потере 3 молекул АТФ.
· на активацию жирной кислоты тратятся 2 макроэргические связи.
· таким образом, энергетический выход 90 + 32 - 3 - 2 =117 молекул АТФ.
Биосинтез жирных кислот (уметь писать реакции): органная и внутриклеточная локализация процесса, коферменты, биологическая роль в организме.
Высшие жирные кислоты могут быть синтезированы в организме из метаболитов углеводного обмена. Исходным соединением для этого биосинтеза является ацетил-КоА, образующийся в митохондриях из пирувата – продукта гликолитического распада глюкозы. Место синтеза жирных кислот – цитоплазма клеток, где имеется мультиферментный комплекс синтетаза высших жирных кислот. Этот комплекс состоит из шести ферментов, связанных с ацилпереносящим белком, который содержит две свободные SH-группы (АПБ-SH). Синтез происходит путём полимеризации двууглеродных фрагментов, конечным продуктом его является пальмитиновая кислота – насыщенная жирная кислота, содержащая 16 атомов углерода. Обязательными компонентами, участвующими в синтезе, являются НАДФН (кофермент, образующийся в реакциях пентозофосфатного пути окисления углеводов) и АТФ.
20.1.2.Ацетил-КоА поступает из митохондрий в цитоплазму при помощи цитратного механизма (рисунок 20.1). В митохондриях ацетил-КоА взаимодействует с оксалоацетатом (фермент –цитратсинтаза), образующийся цитрат переносится через митохондриальную мембрану при помощи специальной транспортной системы. В цитоплазме цитрат реагирует с HS-КоА и АТФ, вновь распадаясь на ацетил-КоА и оксалоацетат (фермент – цитратлиаза).
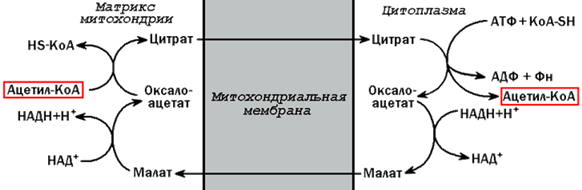
Рисунок 20.1. Перенос ацетильных групп из митохондрий в цитоплазму.
20.1.3.Начальной реакцией синтеза жирных кислот является карбоксилирование ацетил-КоА с образованием малонил-КоА (рисунок 20.2). Фермент ацетил-КоА-карбоксилаза активируется цитратом и ингибируется КоА-производными высших жирных кислот.

Рисунок 20.2. Реакция карбоксилирования ацетил-КоА.
Затем ацетил-КоА и малонил-КоА взаимодействуют с SH-группами ацилпереносящего белка (рисунок 20.3).
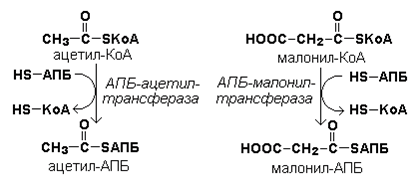
Рисунок 20.3. Взаимодействие ацетил-КоА и малонил-КоА с ацилпереносящим белком.
Далее происходит их конденсация, декарбоксилирование и восстановление образовавшегося продукта (рисунок 20.4).
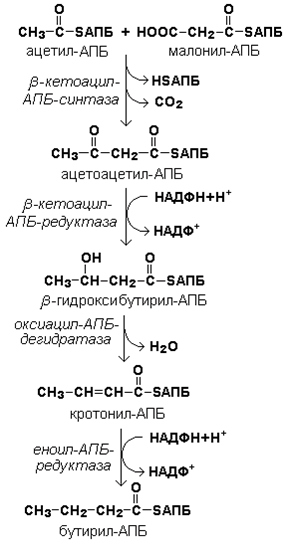
Рисунок 20.4. Реакции одного цикла биосинтеза жирных кислот.
Продукт реакции взаимодействует с новой молекулой малонил-КоА и цикл многократно повторяется вплоть до образования остатка пальмитиновой кислоты.
основные особенности биосинтеза жирных кислот по сравнению с β-окислением:
· синтез жирных кислот в основном осуществляется в цитоплазме клетки, а окисление – в митохондриях;
· участие в процессе связывания СО2 с ацетил-КоА;
· в синтезе жирных кислот принимает участие ацилпереносящий белок, а в окислении – коэнзим А;
· для биосинтеза жирных кислот необходимы окислительно-восстановительные коферменты НАДФН, а для β-окисления – НАД+ и ФАД.
Дата добавления: 2018-08-06; просмотров: 489; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!