Производные вещества протопласта
Анатомия растений
Фитоцитология
Фитоцитология –раздел ботаники, изучающий живые растительные клетки, их происхождение, строение, функционирование, процессы клеточного размножения, старения и гибели
Клетка –наименьшая структурная и функциональная единица живого организма. Клетка – это самовоспроизводящаяся форма материи, состоящая из определённых молекулярных структур, собранных в определённой последовательности. Форма и атомарный состав молекул, формирующих клетку, определяет их функции и свойства.
Все процессы (реакции), проходящие в клетке, называются метаболизмом. Метаболизм представляет собой непрерывный процесс передачи энергии химических связей (удерживающих атомы в составе молекулы) от одних молекул другим. В реакциях метаболизма выделяют две составляющие:
катаболизм – реакции разрушения сложных молекулярных структур на простые, проходящие с выделением энергии;
анаболизм – реакции синтеза, создания сложных молекулярных структур из простых, протекающие с поглощением энергии.
Основным источником энергии для растительных организмов является энергия солнечного света. Под действием световой энергии в хлоропластах происходит синтез глюкозы (С6Н12О6) из углекислого газа (СО2) и водорода (Н) воды, т.е. энергия света преобразуется в энергию химических связей, которая удерживает атомы в молекуле глюкозы. В дальнейшем часть глюкозы в митохондриях сжигается (разрушается) до углекислого газа и водорода, который, взаимодействуя с кислородом, образует воду. Энергия, выделяющаяся при разрушении глюкозы, соединяет аденозиндифосфорную кислоту (АДФ) с ещё одной фосфорной кислотой, в результате чего получается универсальная молекула-энергоноситель аденозинтрифосфорная кислота (АТФ). Энергия химической связи, заключённая в молекуле АТФ, выделяется при её разрушении и используется для перестройки молекулы глюкозы в другие молекулярные вещества с изменённой структурой, составом и функциями. Так из глюкозы через цепь реакций катаболизма-анаболизма образуются липиды и аминокислоты, а из аминокислот – белки.
|
|
|
Вещества, образованные клеткой, в зависимости от условий, формы и свойств молекул, составляющих их, могут выполнять ряд функций:
1. Строительную – формировать долговременные комплектующие структуры протопласта (органеллы) и межклеточного вещества;
2. Запасающую – долговременно храниться в органеллах клетки, в неактивном состоянии, содержа в себе энергию химических связей;
3. Энергетическую – передавать энергию химических связей в реакциях катаболизма-анаболизма от одних молекул другим;
|
|
|
4. Управляющую – регулировать ход и скорость химических реакций в клетке.
Функционально клетку делят на:
протопласт – «живую» организующую часть, создающую условия для прохождения реакций метаболизма;
производные протопласта – «неживую» организованную часть, т.е. вещества и структуры, образованные в результате метаболизма.

Рис. 1. Общее строение растительной клетки
Протопласт
В состав протопласта входят:
плазмалемма – внешняя мембрана протопласта, обладающая свойством полупроницаемости и выполняющая барьерную функцию;
гиалоплазма – коллоидный раствор, заполняющий протопласт внутри, создающий среду для прохождения реакций метаболизма;
органеллы клетки (9 типов) – обновляющиеся молекулярные структуры, характеризующиеся определенной формой, свойствами и функциями. Взаимодействие органелл клетки обеспечивает её работу как открытой подсистемы организма, обеспечивает пространственную изоляцию и прохождение разнонаправленных реакций метаболизма в клетке.
К органеллам клетки относятся:
1. Ядро – двумебранная органелла клетки, заполненная ядерным соком (кариоплазмой), в котором находятся хроматин (ДНК) и ядрышки (РНК) – молекулы генетического программного обеспечения клетки. Основная функция ядра – реализация генетической программы развития и функционирования клетки и всего организма в целом.
|
|
|
2. Пластиды – двумембранные полуавтономные органеллы клетки, заполненные коллоидным раствором – стромой (матриксом), в котором находится ДНК, РНК и рибосомы. Выделяют три типа пластид:
Хлоропласты – пластиды, у которых внутренняя мембрана образует сеть взаимосвязанных между собой плоских пузырьков – ламел (тилакоидов), собранных в стопки – граны. В мембранах ламел находятся световоспринимающие пигменты хлорофилл, каротин, ксантофилл. Функция хлоропластов – фотосинтез – преобразование энергии солнечного света в энергию химических связей молекул, используемую для синтеза глюкозы из углекислого газа и водорода воды. Хлоропласты характерны для клеток хлоренхимы листьев.
Хромопласты – пластиды без сложной внутренней мембранной структуры, в которых отсутствует пигмент хлорофилл, а каротин и ксантофилл сконцентрированы в строме пластида. Хромопласты образуются в результате старения хлоропластов (пожелтение листьев).
Лейкопласты – пластиды со слаборазвитой внутренней мембраной, не имеющие пигментов. Функция – накопление в строме запасных питательных веществ (белков, липидов, углеводов). Характерны для всех тканей, особенно имеющих запасающую функцию.
|
|
|
3. Митохондрии – двумембранные полуавтономные органеллы клетки, заполненные коллоидным раствором – матриксом, в котором находится ДНК, РНК и рибосомы. Внутренняя мембрана митохондрий больше по площади, чем наружная и образует складки – кристы. Основная функция митохондрий – разрушение органических соединений до углекислого газа и водорода воды, и соединение АДФ (аденозиндифосфорной кислоты) с молекулой фосфорной кислоты в АТФ (аденозинтрифосфорную кислоту) – молекулу-энергоноситель.
4. Эндоплазматическая сеть (ЭПС) – одномембранная органелла в виде системы взаимосвязанных мембранных плоских цистерн и трубочек. Выполняет функцию транспорта веществ внутри системы. Различают гранулярную ЭПС (на поверхности которой находятся рибосомы) и гладкую ЭПС (без рибосом на поверхности). Из мембран ЭПС в протопласте образуются органеллы: аппарат Гольджи, вакуоли и оболочка ядра.
5. Аппарат Гольджи – одномембранная органелла, состоящая из нескольких плоских мембранных пузырьков – диктиосом. Образование аппарата Гольджи начинается с выдувания ЭПС мембранных пузырьков, заполненных определёнными веществами. Эти пузырьки сливаются в плоские цистерны диктиосом, где происходят химические реакции преобразования поступивших веществ в другие соединения. Последняя диктиосома, с готовыми веществами, распадается на маленькие секреторные пузырьки, которые встраиваются в плазмалемму, выводя внутреннее содержимое за пределы протопласта на клеточную стенку. Функция аппарата Гольджи – выделительная. Вместе с ЭПС может строить вакуоли, в которые выделять преобразованные вещества.
6. Вакуоли – одномембранные пузырьки, образованные ЭПС или аппаратом Гольджи. В зависимости от функций веществ, заполняющих вакуоли, их делят на:
Сократительные (пульсирующие) вакуоли – заполненные клеточным соком (раствором солей и других веществ). Выполняют функцию поддержания тургорного (осмотического) давления в клетке, накопления растворимых защитных и запасающих веществ;
Запасающие вакуоли – накапливающие нерастворимые запасные питательные вещества (белки, липиды, углеводы);
Лизосомы – вакуоли, заполненныеферментами, разрушающими ненужные клетке макромолекулы и неработающие органеллы;
Пероксисомы – вакуоли, заполненные веществами, нейтрализующими активные формы кислорода, которые образуются при фотосинтезе, дыхании, разрушении макромолекул.
7. Рибосомы – безмембранные органеллы, состоящие из молекул РНК и белка. Выполняют функцию синтеза белка из аминокислот.
8. Микротрубочки – органеллы в виде трубочек, образованных белком тубулином. Выполняют в клетке опорную функцию (служат опорой для всех органелл протопласта, обеспечивают их перемещение по клетке и поддерживают форму протопласта).
9. Микрофиламенты – органелла в виде тяжей, образованных белком актином. Функция – генерация (создание) тока гиалоплазмы по клетке. Вместе с микротрубочками образует опорную структуру клетки – цитоскелет.
Производные вещества протопласта
К производным веществам протопласта относятся:
1. Физиологически активные вещества, регулирующие функционирование, рост и развитие клетки и организма в целом:
Ферменты – белковые молекулы-катализаторы, активирующие химические реакции в клетке.
Витамины – небелковые молекулы-катализаторы, активирующие химические реакции в клетке.
Гормоны – вещества, активирующие деление клеток, ростовые процессы в них и организме в целом.
Фитонциды – защитные отравляющие вещества, защищающие клетку и организм от повреждений другими организмами.
2. Запасные вещества – временно неактивные полимерные вещества, содержащие большое количество энергии в химических связях. Откладываются в лейкопластах и вакуолях.
3. Клеточный сок – раствор солей и других веществ, заполняющий сократительные вакуоли. Выполняет регуляцию давления в клетке, поглощения воды, накопления защитных веществ.
4. Клеточная стенка – целлюлозная оболочка клетки, выполняющая функции внешней опоры, защищающей протопласт, и межклеточного вещества, соединяющего клетки между собой в ткань. Образуется в результате выделения целлюлозы и других веществ аппаратом Гольджи за пределы протопласта (в телофазе митоза или мейоза клеточная стенка делит материнскую клетку на две дочерние). Клеточная стенка в своём строении может иметь три слоя:
серединная пластинка – является общей для двух соседних клеток, образуется аппаратом Гольджи первой. Состоит из пектиновых веществ;
первичная клеточная оболочка – откладывается на серединную пластинку изнутри небольшим слоем. Образована преимущественно целлюлозой, которая пропитывается гемицеллюлозой, гликопротеидами и пектиновыми веществами, эти вещества придают клеточной оболочке гибкость и эластичность;
вторичная клеточная оболочка – образуется в клетках тканей, которые выполняют опорную функцию, имеет несколько слоёв, в которых целлюлоза откладывается разнонаправлено. Эта оболочка намного жёстче и толще чем первичная оболочка. Целлюлоза пропитывается гемицеллюлозой и лигнином.
К структурным компонентам клеточной стенки относятся:
плазмодесмы – участки цитоплазмы, проходящие через клеточную стенку и соединяющие протопласты двух соседних клеток;
десмотрубочки – участки ЭПС внутри плазмодесм, соединяющие ЭПС двух соседних клеток;
поровое поле – тонкий участок первичной клеточной оболочки вокруг плазмодесмы;
пора – отверстие во вторичной клеточной оболочке над поровым полем. Поры бывают простые (с ровным краем) и окаймлённые (с нависающим краем над поровым полем).
Жизненный цикл клетки представляет собой период существования клетки от момента образования до момента деления или гибели. В жизненном цикле клетки выделяют два периода:
интерфаза – период существования и функционирования клетки от момента образования до начала деления;
деление – период образования из одной клетки двух (митоз) или четырёх (мейоз).
Интерфаза включает в себя три периода: G1 (G0), S и G2.
Период интерфазы G1 начинается с момента образования клетки и связан с ростом клетки и увеличения количества её органелл. В зависимости от местоположения в организме клетки функционально специализируются, приобретают характерное строение и выполняют необходимую работу. Если клетка не программируется организмом на дальнейшее деление, она остаётся на этой фазе развития, которая обозначается как (G0) и продолжает выполнять свои функции до момента гибели. Если клетка программируется организмом на дальнейшее деление, она переходит в период интерфазы S.
Период интерфазы S связан с удвоением наследственной генетической информации в ядре клетки, т.е. происходит удвоение молекул ДНК, для передачи дубликатов генетической программы в две дочерние клетки во время деления.
Период интерфазы G2 связан с подготовкой клетки к делению, образуются структуры и вещества, используемые клеткой во время деления.

| Рис. 2. Жизненный цикл клетки: НК – начало и конец жизненного цикла, G1 – фаза роста клетки, G0 – фаза специализации клетки, S – фаза репликации (удвоения) ДНК, G2 – фаза подготовки к делению клетки, М – деление клетки митозом или мейозом |
Митоз – деление клетки на две дочерние с сохранением хромосомного набора (дубликаты генетической программы, зашифрованной на молекулах ДНК, передаются в две образующиеся дочерние клетки).
Митоз состоит из четырёх фаз:
1. Профаза. В клетке разрушается ядерная оболочка, хроматин (рабочая форма ДНК) и ядрышки (РНК) упаковываются в хромосомы (транспортную форму ДНК), т.е. генетическая программа архивируется для транспортировки.
2. Метафаза. Хромосомы выводятся на экватор клетки и к их половинкам (хроматидам) присоединяются микротрубочки веретена деления.
3. Анафаза. Микротрубочки веретена деления сокращаются и разводят половинки хромосом к разным полюсам клетки.
4. Телофаза. На полюсах клетки вокруг половинок хромосом ЭПС формирует новые ядерные оболочки. Половинки хромосом распаковываются (разархивируются) в хроматин, т.е. генетическая программа разархивируется. Аппарат Гольджи строит клеточную стенку, разделяющую материнскую клетку на две дочерние.

Рис. 3. Деление растительной клетки митозом
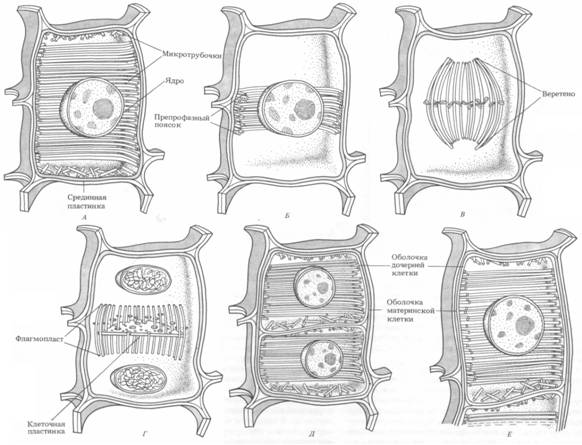
Рис. 4. Перераспределение микротрубочек в течение клеточного цикла
по мере образования клеточной стенки: А – интерфаза G2 , Б – перед профазой,
В – метафаза, Г – телофаза (цитокинез), Д, Е – интерфазы G1
Мейоз – редукционное деление клетки на четыре дочерние с уменьшением двойного (диплоидного 2n) набора хромосом вдвое и рекомбинацией (изменением) генетической программы в новых клетках.
Мейоз проходит в два этапа, каждый из которых состоит из четырёх фаз.
1-й этап:
1. Профаза 1. В клетке разрушается ядерная оболочка, хроматин (рабочая форма ДНК) и ядрышки (РНК) упаковываются в хромосомы (транспортную форму ДНК), т.е. генетическая программа архивируется для транспортировки. Гомологичные одинаковые хромосомы (отцовские и материнские) скручиваются между собой – конъюгация и обмениваются генами (участками хромосом) – кроссинговер, после чего расплетаются.
2. Метафаза 1. Хромосомы выводятся на экватор клетки и к гомологичным хромосомам присоединяются микротрубочки веретена деления, идущие из разных полюсов клетки.
3. Анафаза 1. Микротрубочки веретена деления сокращаются и разводят гомологичные хромосомы к разным полюсам клетки.
4. Телофаза 1. Гомологичные хромосомы группируются на полюсах клетки. Аппарат Гольджи формирует клеточную стенку, которая делит материнскую клетку на две дочерние по экватору.
После первого этапа деления мейозом в дочерних клетках остаётся половинный (гаплоидный n) набор хромосом. Период подготовки клеток ко второму делению называется интеркинез.
2-й этап деления клеток мейозом идентичен митозу и проходит в четыре фазы синхронно в двух дочерних клетках от первого деления (смотри митоз).
В результате деления клеток мейозом образуются четыре гаплоидные клетки, в каждой из которых наследственная информация изменена и отличается от генетической программы материнской клетки.

Рис. 5. Схема мейоза с двумя парами хромосом (показаны не все стадии)
Фитогистология
Фитогистология – наука, изучающая строение, развитие и функции растительных тканей.
Ткань – совокупность клеток, которые имеют общее происхождение, одинаковое строение и выполняют одинаковую функцию в теле организма.
По происхождению ткани делят на первичные, вторичные и третичные. Первичные ткани возникают из развивающейся зиготы (оплодотворённой яйцеклетки), вторичные ткани развиваются из первичных, а третичные – из вторичных.
Ткани также делят на живые (в клетках которых сохраняется живой протопласт) и мёртвые (в клетках которых протопласт отмирает и остаётся только клеточная стенка, выполняющая защитную, опорную и др. функции в органах растений).
Образование новых тканей и органов связано с изменением формы организма и переходом его на новый этап развития.
В растительных организмах выделяют шесть типов тканей:
1. Меристематические (меристемы) или образовательные,
2. Основные,
3. Граничные,
4. Проводящие,
5. Механические,
6. Выделительные.
1. Меристематические ткани, или меристемы, состоят из клеток, управляющих направлением и темпом роста организма. Функция меристематических клеток – деление митозом или мейозом с образованием новых (дочерних) клеток. Образованные клетки разделяются (дифференцируются) по выполняемым функциям (в интерфазе G0) в зависимости от местоположения в растительном организме, т.е. преобразовываются во все остальные типы тканей.
По месторасположению в теле растения меристемы делятся на:
А. Верхушечные или апикальные – расположенные в точках роста почек и кончиков корней, например: туника формирует покровную ткань эпидермис, меристема конуса нарастания – боковые меристемы.
Б. Боковые или латеральные – находятся внутри стеблей и корней и обеспечивают их рост в толщину, например: первичные меристемы перицикл и прокамбий формируют ткани первичных проводящих пучков, основной паренхимы (залегающей между ними) и вторичные боковые меристемы камбий и феллоген. Последние обеспечивают вторичный рост (утолщение) стеблей и корней за счёт формирования вторичных комплексных проводящих тканей – ксилемы (К2) и флоэмы (Ф2).
В. Вставочные или интеркалярные – находятся в междоузлиях побегов и обеспечивают их вытяжение (удлинение).
Г. Раневые или травматические – возникают на местах повреждений и закрывают раны, образовывая клетки основной заполняющей паренхимы.
2. Основные паренхимные ткани – окружают проводящую систему растений, заполняя их органы. В зависимости от выполняемой функции выделяют три типа основных паренхим:
А. Хлоренхима или хлорофиллоносная паренхима – находится в листьях (между жилками) и молодых побегах; зеленого цвета; выполняет ассимиляционную функцию (образования углеводов в результате фотосинтеза); по строению клеток бывает трёх типов:
губчатая – у однодольных и двудольных покрытосеменных растений,
столбчатая – у двудольных покрытосеменных растений,
складчатая – у голосеменных растений.
Б. Аэренхима или воздухоносная паренхима – образуется вокруг проводящих пучков корней и побегов растений, растущих в переувлажнённых, заболоченных почвах или воде. Имеет большие межклетники, по которым воздух подаётся к подземным и подводным органам растений, находящимся в среде с низким содержанием кислорода.
В. Запасающая паренхима – накапливает в своих клетках запас питательных веществ, располагается в запасающих органах растений: корнях, стеблях, листьях.
3. Пограничные ткани – выполняют барьерную функцию, ограничивая органы растений снаружи и окружая проводящую систему внутри. В зависимости от местоположения и функции пограничные ткани делят на:
А. Покровные – внешние пограничные ткани, покрывающие органы растений снаружи, выполняющие защитную, газообменную и терморегулирующую функции. К ним относятся:
Эпидермис – живая первичная покровная ткань, покрытая защитным слоем кутина и воска. В составе ткани находятся плоские плотно сомкнутые эпидермальные клетки; вертикальные вытянутые клетки трихом (волосков), наклоном которых управляют клетки в их основании; и парные клетки устьиц, которые в тургорном состоянии образуют щель между собой, а без тургора смыкаются, останавливая газообмен и транспирацию (испарение) воды из органов растения.
Пробка – мертвая вторичная покровная ткань, клетки которой заполнены воздухом, а клеточная стенка пропитана суберином. Пробка образуется на многолетних осевых органах растений (корнях и стеблях) под эпидермисом из вторичной меристемы – феллогена, заменяя его. Покровная ткань – пробка, меристематическая ткань – феллоген и основная паренхимная ткань – феллодерма (расположенная под феллогеном) вместе образуют комплексную вторичную покровную ткань – перидерму. Перидерма закладывается ежегодно в более глубоких слоях живых клеток основной паренхимы.
Корка или ритидом – мертвая комплексная третичная покровная ткань, образованная несколькими отмершими слоями перидермы, колленхимы (механической ткани) и участков основной паренхимы коры, отделяемых новым закладывающимся слоем перидермы.
Б. Всасывающие – внешние пограничные ткани с функцией поглощения:
Ризодерма или эпиблема – живая пограничная ткань, покрывающая корень в зоне поглощения, имеющая вытянутые клетки корневых волосков, через которые в корень всасывается вода с растворенными в ней минеральными веществами.
Веламен – многослойная живая пограничная ткань, покрывающая воздушные корни растений эпифитов, поглощающая воду из воздуха.
Гаустории – ткань органов растений-паразитов, которыми они проникают в растение хозяина, откуда поглощают воду с органическими и минеральными веществами.
В. Фильтрационные – внутренние пограничные ткани с функцией фильтрации водных растворов.
Экзодерма – живая фильтрационная ткань, расположенная под ризодермой корня, пропускающая воду через клеточные оболочки своих клеток – апопласт («фильтр грубой очистки»).
Эндодерма – живая фильтрационная ткань, расположенная вокруг проводящего пучка, пропускающая воду через протопласты своих клеток – симпласт («фильтр тонкой очистки»).
4. Проводящие ткани – составляют основу транспортной (проводящей) системы, представленной в растениях проводящими пучками. Проводящие пучки состоят из двух комплексных проводящих тканей – ксилемы и флоэмы.
А. Ксилема или древесина – комплексная проводящая ткань, обеспечивает поступление воды с минеральными веществами из корней в листья.
У покрытосеменных растений ксилема состоит из трёх тканей:
1. Сосуды или трахеи – мёртвая проводящая ткань, проводит непосредственно воду с минеральными веществами;
2. Либриформ или склеренхима ксилемы – мёртвая механическая ткань, являющаяся опорой для сосудов;
3. Запасающая паренхима – живая основная ткань, накапливающая запас питательных веществ и воды в протопластах своих клеток.
У голосеменных растений основу ксилемы составляют клетки мёртвой проводящей ткани – трахеиды, выполняющие одновременно и водопроводящую, и механическую (опорную) функции.
Б. Флоэмаили луб – комплексная проводящая ткань, транспортирующая органические вещества из листьев в корни растений.
У покрытосеменных растений включает в себя три ткани:
1. Ситовидные трубки – живая проводящая ткань, проводит непосредственно органические вещества;
2. Лубяные волокна или склеренхима флоэмы – мёртвая механическая ткань, дающая опору ситовидным трубкам и проводящему пучку в целом;
3. Запасающая паренхима – живая основная ткань, накапливающая резерв питательных веществ в протопластах своих клеток.
У голосеменных растений основу флоэмы составляют клетки живой проводящей ткани – ситовидные клетки, выполняющие одновременно и проводящую, и механическую (опорную) функции.
В зависимости от размещения ксилемы и флоэмы в проводящих пучках, их разделяют на:
1. Коллатеральные – ксилема и флоэма в пучке расположены одна на другой (например, у кукурузы),
2. Биколлатеральные – ксилема зажата между двумя пучками флоэмы или наоборот (например, у тыквы),
3. Радиальные – ксилема и флоэма чередуются между собой по окружности (радиусу) (например, в первичном строении корней),
4. Концентрические – ксилема расположена в центре, флоэма – по периметру (периферии) проводящего пучка (например, в стеблях древесных растений).
5. Механические ткани – имеют утолщённые клеточные стенки, выдерживают статические и динамические механические нагрузки, выполняют опорную и защитную функции в органах растений. Различают три типа механических тканей:
1. Колленхима – живая механическая ткань, наряду с опорной функцией, может выполнять фотосинтетическую и запасающую функции. В зависимости от места утолщения клеточной стенки колленхиму подразделяют на угловую, пластинчатую и губчатую.
2. Склеренхима – мёртвая механическая ткань, клетки которой прозенхимные в виде волокон, входит в состав ксилемы и флоэмы.
3. Склереиды – мёртвая механическая ткань, клетки которой паренхимные с очень утолщёнными каменистыми клеточными стенками, образуют в плодах косточки (вишня) и скорлупу (орех).
6. Выделительные ткани – представляют собой систему клеток, которые накапливают или выделяют наружу различные вещества (отходы метаболизма). По размещению на растении различают:
А. Выделительные ткани внешней секреции, расположенные снаружи и выделяющие вещества наружу растения:
Железистые чешуйки и волоски – вытянутые клетки (трихомы) эпидермиса, в которых накапливаются или через которые выделяются вещества (например, муравьиная кислота у крапивы);
Пищеварительные железы растений-хищников (например, росянки);
Гидатоды – водные устьица, через которые выделяются излишки воды из листьев;
Нектарники – железы, выделяющие сахарный сироп в основании тычинок цветков.
Б. Выделительные ткани внутренней секреции, находятся внутри растения и накапливают секреторные вещества внутри клеток либо в межклетниках:
Схизогенные вместилища – каналы или камеры, образованные межклетниками железистых (секреторных) клеток, в которые они выделяют свой секрет (например, смоляные ходы у сосны);
Лизогенные вместилища – камеры, образованные на месте разрушения (распада) группы клеток, после накопления в них отработанных веществ (например, в кожице цитрусовых плодов);
Идиобласты – накопительные одиночные клетки в разных тканях, которые собирают отработанные вещества из соседних клеток;
Молочники – живые клетки, которые содержат в вакуолях млечный сок (латекс), выполняют функцию резервуаров дезинфицирующих и запасных веществ (например, молочай, одуванчик).
Органография
Органография – наука, изучающая анатомическое строение растительных организмов.
В строении тел растений выделяют два основные части – корень и побег. Побеги в свою очередь включают в себя осевую часть (орган) – стебель, периферийную часть (орган) – лист, верхушечную часть – почку. Побеги функционально делят на вегетативные (ростовые) и генеративные (органы размножения). Генеративные побеги у покрытосеменных растений представлены цветками, у голосеменных – шишками.
Своё развитие растительный организм начинает из оплодотворённой яйцеклетки, которая в результате деления формирует зародыш семени. Зародыш семени состоит из вегетативных органов: зачаточного корешка, стебля, двух семядолей (у однодольных покрытосеменных растений одна семядоля не развита) и почки. Генеративные побеги развиваются на вегетативных после прохождения растением стадии яровизации.
Корень – вегетативный орган растения, выполняющий функции:
1. Закрепления растения в почве,
2. Накопления питательных веществ,
3. Вегетативного размножения,
4. Поглощения из почвы воды с минеральными веществами и транспортировка их в стебель.
В зависимости от происхождения корни делят на:
1. Главный корень – закладывается и развивается из зародыша семени,
2. Боковые корни – растут из главного корня,
3. Придаточные корни – растут из побега.
Корневая система представляет собой совокупность всех корней растения. В зависимости от того, какие корни участвуют в формировании корневой системы, их делят на три типа:
1. Стержневая корневая система – образована главным и боковыми корнями,
2. Мочковатая корневая система – образована придаточными корнями,
3. Смешанная корневая система – образована главным и придаточными корнями, которые не отличаются по размерам и строению между собой.
Так же у корней наблюдаются морфологические и анатомические видоизменения в зависимости от выполняемой ими функции:
1. Корнеплоды – утолщённые запасающие главные корни (свёкла, морковь),
2. Корнеклубни – утолщённые запасающие придаточные корни (георгин),
3. Воздушные корни – поглощают воду из атмосферы,
4. Дыхательные корни – поглощают воздух, выходя на поверхность заболоченных почв и воды.
5. Втягивающие корни – погружающие многолетние стебли травянистых растений в почву на зимовку, за счёт сокращения (люцерна, эспарцет).
6. Ходульные корни – корни-подпорки, выполняющие опорную функцию.
7. Корни-присоски – корни растений паразитов.
8. Микориза – симбиоз растения и мицелия гриба, который поставляет в корневую систему воду, а извлекает органические вещества.
9. Бактериальные клубеньки – симбиоз растения и азотфиксирующих бактерий, которые создают колонии в коре корня.
В анатомическом строении корня выделяют четыре зоны:
| 1. Зона деления – апикальная зона, защищённая корневым чехликом, в которой находятся меристематически активные клетки (постоянно делящиеся), формирующие все остальные ткани корня; 2.Зона роста – характеризуется ростом (вытягиванием) и специализацией клеток, образованных в зоне деления, по выполняемым функциям, формируются фильтрационные и проводящие ткани; 3. Зона поглощения имеет наибольшую поверхность за счёт большого количества корневых волосков ризодермы, которые всасывают воду из почвы и подают её через фильтрационные ткани в проводящий пучок; 4. Зона проведения отвечает за транспорт воды из зоны поглощения в стебель. Здесь могут закладываться боковыекорни. | 
|
В своём развитии корень растений может трижды менять своё строение.
Первичное строение корня:
Ростовые процессы в корне начинаются в зоне деления, где находятся три слоя инициальных клеток. Инициальные клетки путём деления образуют апикальные меристемы, клетки которых, вытягиваясь в зоне роста, формируют:
1-й слой – покрывную всасывающую ткань – ризодерму и корневой чехлик, защищающий точку роста (деления);
2-й слой – первичную кору корня: экзодерму – апопластную фильтрационную ткань, мезодерму – запасающую основную паренхиму и эндодерму – симпластную фильтрационную ткань. Первичная кора корня находится под ризодермой и окружает центральный цилиндр;
3-й слой – центральный цилиндр корня (радиальный проводящий пучок), под эндодермой выстлан слоем клеток перицикла (первичной боковой меристемы), в центре заполнен основной паренхимой, в которую радиально погружены проводящие ткани ксилемы и флоэмы, между которыми остаются участки прокамбия – боковой меристематической ткани.


Рис. 6. Гистологическая схема первичного строения корня
В зоне всасывания вода, поглощённая корневыми волосками ризодермы, проходит через клеточные стенки экзодермы, затем по межклетникам мезодермы и фильтруется через протопласты эндодермы. Попадая в центральный цилиндр, вода по сосудам ксилемы поднимается в зону проведения и транспортируется в стебель. В зоне проведения ризодерма, покрывающая корень, отмирает и на поверхность выходит экзодерма.
У двудольных растений в зоне проведения происходит переход от первичного строения корня к вторичному. Этот переход связан с меристематической деятельностью клеток перицикла и прокамбия, зажатого между пучками ксилемы и флоэмы. Делящиеся клетки перицикла формируют под эндодермой: перидерму (пробку, феллоген и феллодерму), колленхиму и основную паренхиму коры.
Клетки прокамбия, функционально соединяясь с отдельными участками перицикла, образуют непрерывный изогнутый слой камбия (вторичной боковой меристемы), который разделяет первичную ксилему и флоэму. Деятельность камбия направлена на трансформацию закрытого радиального проводящего пучка в открытый концентрический. Клетки камбия делятся, образуя из внутренних слоёв вторичную ксилему (поверх первичной ксилемы), из внешних – вторичную флоэму (под первичной флоэмой). Участки камбия, сформированные клетками перицикла, откладывают основную паренхиму и в центр и наружу, создавая сердцевинный луч. Образование вторичной ксилемы идёт быстрее, чем флоэмы и основной паренхимы, поэтому через некоторое время слой камбия в виде звезды выгибается в кольцо. Разрастающиеся вторичные ткани разрывают первичную кору, которая постепенно отмирает. Этот процесс называется линька корня.

Рис. 7. Вторичное строение корня в поперечном срезе
В корне свёклы происходит переход от вторичного строения корня к третичному, и связан он с образованием дополнительных слоёв камбия из меристематически активных клеток основной паренхимы коры. На дополнительных слоях камбия формируются открытые коллатеральные проводящие пучки, образованные третичной флоэмой и третичной ксилемой. Каждое кольцо камбия соответствует паре листьев на укороченном стебле корнеплода, т.е. связано с их проводящей системой.

Рис. 8. Третичное строение корня в поперечном срезе
Побег в своём строении представляет собою серию метамеров – повторяющихся частей, закладка которых производится апикальной меристемой почки в точке роста. Метамер состоит из осевой части – стебля и периферийной части – листа. В стеблевой части метамера выделяют зону узла – места крепления листа и междоузлия – зону между узлами. В основании междоузлия находятся клетки интеркалярной (вставочной) меристемы, которая обеспечивает вытяжение междоузлий и выход метамеров из состава почки. Размещение листьев на стебле может быть очерёдным (когда в узле закладывается один лист) или мутовчатым (когда в узле закладывается два – супротивное расположение, и более листьев).
Видоизменения побегов у растений представлены:
1. Клубнями – утолщёнными запасающими подземными побегами (картофель),
2. Луковицами – увеличенными подземными запасающими почками (тюльпан),
3. Корневищами – горизонтальными подземными запасающими побегами (пырей),
4. Кочаном – увеличенной запасающей надземной почкой (капуста),
5. Усами – горизонтальными надземными побегами (земляника),
6. Усиками – видоизменениями листа цепляющихся растений (горох),
7. Каудексом – укороченным стеблем многолетних трав (люцерна),
8. Кладодиями – уплощёнными стеблями в форме листьев (иглица, аспарагус),
9. Иглами – видоизменениями листьев (кактус).
Анатомическое строение почки:
Нарастание побега происходит верхушкой. Верхняя часть конуса нарастания почки защищена туникой – зачаточной покровной тканью, которая позднее развивается в эпидермис. Под туникой находятся клетки корпуса – апикальной меристемы конуса нарастания, которые дают объём побега, формируя метамеры. Клетки корпуса концентрически образуют две зоны – центральную и периферийную меристемы. Клетки центральной меристемы становятся основной паренхимой, а из клеток периферийной меристемы формируются зачатки листьев – примордии, коровая часть стебля и проводящая система побега.
Проводящая система у растений закладывается:
1. Одним концентрическим пучком (у древесных растений);
2. Несколькими коллатеральными (биколлатеральными) пуками, расположенными по окружности, которые позже могут соединяться в один концентрический пучок (у двудольных травянистых растений);
3. Коллатеральными пучками по нескольким окружностям, за счёт раздвоения пучков, где проводящие пучки одной окружности связаны с проводящей системой одного листа (у однодольных растений).
В зависимости от анатомического строения (тканевого состава) и морфологических особенностей стебли растений классифицируют:
По механическим свойствам:
1. Древесные – стебли деревьев (стволы) и кустов (стволики);
2. Одревесневающие – многолетние стебли кустарничков, полукустарников и трав;
3. Травянистые – однолетние побеги трав без признаков одревеснения.
По направлению роста:
1. Прямостоячие – стебли, растущие в вертикальном положении (пшеница);
2. Восходящие – стебли, из горизонтального положения поднимающиеся под углом к поверхности почвы (просо куриное);
3. Вьющиеся – стебли, обвивающие вертикальные опорные конструкции (вьюнок, повой);
4. Цепляющиеся – стебли, закрепляющиеся на вертикальных опорных конструкциях при помощи усиков (видоизменённых листьев или побегов) (виноград);
5. Ползучие – стебли, закрепляющиеся в горизонтальном направлении при помощи усов и усиков, а также придаточными корнями, образующимися в местах контакта стеблей с поверхностью почвы (тыква, огурец, арбуз);
6. Стелящиеся – стебли, растущие в горизонтальном направлении без дополнительных креплений (портулак).
Первичное строение стеблейоднодольных покрытосеменных растений
Стебель (соломина) ржи покрыт эпидермисом, под которым находится опорная склеренхима стебля с участками фотосинтезирующей хлоренхимы. В склеренхиме стебля, как и основной паренхиме под ней, вертикально проходят коллатеральные проводящие пучки, строение которых подобно у всех однодольных злаковых растений. В центральной части стебель пустой, что облегчает конструкцию побега и делает его механически более прочным.
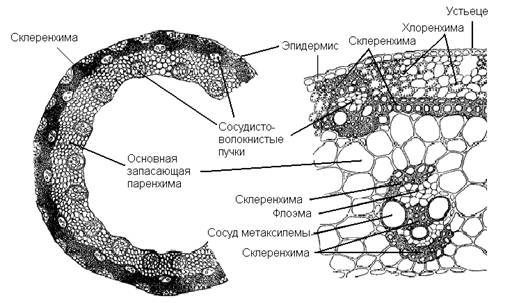
Рис. 9. Поперечный срез соломины ржи
Похожее строение стебля имеет кукуруза. Утолщение стебля на первых этапах развития растения обеспечивает меристематическая ткань – перицикл, расположенная под эпидермисом. Перицикл позже дифференцируется на хлоренхиму (фотосинтезирующую ткань) и склеренхиму (опорно-механическую ткань). Под склеренхимой находится запасающая паренхима, в которую погружены закрытые коллатеральные проводящие пучки. Эти пучки окружены обкладывающими клетками (эндодермой). Проводящие элементы в пучках (первичная ксилема и первичная флоэма) зафиксированы снаружи склеренхимой.

Рис. 9а. Строение стебля однодольного растения (кукурузы), поперечный разрез.
I – часть стебля: 1 – эпидермис; 2 – механическая ткань (склеренхима); 3 – основная ткань; 4 – сосудисто-волокнистый пучок (схема). II – замкнутый сосудисто-волокнистый пучок: 1 – основная ткань; 2 – механическая ткань (склеренхима); 3 – ситовидные трубки луба (флоэмы) и сопровождающие клетки; 4 – полость крупного сосуда, окруженного одревесневшими клетками; 5 – спиральный сосуд; 6 – кольчатый сосуд; 7 – воздушная полость.
Вторичное строение стеблейдвудольных покрытосеменных растений.
Для двудольных растений характерен вторичный рост осевых органов (корней и побегов) в толщину, за счёт образования вторичных тканей. Стебель тыквы покрыт эпидермисом, под которым находится угловая колленхима и хлоренхима. Жёсткость конструкции стебля предаёт склеренхима, образованная из перицикла. Над перициклом находится слой клеток эндодермы – фильтрационной граничной ткани. В основной паренхиме, под перициклом расположены десять биколлатеральных открытых проводящих пучков. Если рассматривать ткани проводящих пучков тыквы, то они от внешнего края к центру расположены в следующем порядке. Первичная флоэма, образованная прокамбием и вторичная флоэма, образованная камбием, вместе формируют наружную флоэму пучка. Камбий образуется из клеток прокамбия. Вторичная ксилема (большие сосуды) образуется из клеток камбия. Первичная ксилема (маленькие сосуды) закладывается прокамбием. Внутренняя первичная флоэма также образована прокамбием. В центре стебля находится пятилучевая воздушная полость.
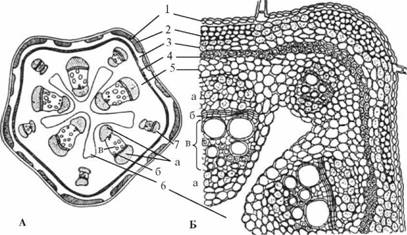
Рис. 10. Поперечный срез стебля тыквы:
А – схема; Б – фрагмент поперечного среза; 1 – эпидерма; 2, 3 – первичная кора: 2 – уголковая колленхима в ребрах и межреберьях; 3– хлоренхима; 4–7 – центральный цилиндр: 4 – перициклическая склеренхима; 5 – основная паренхима; 6 – полость; 7 – биколлатеральный проводящий пучок: а – флоэма наружная и внутренняя; б – многорядный камбий; в – ксилема

Рис. 10а. Поперечный срез стебля клевера
Однолетний молодой стебель клевера, как и у предыдущего растения, покрыт эпидермисом. Механическая ткань колленхима под эпидермисом одновременно выполняет и фотосинтетическую функцию. Под колленхимой в основной паренхиме кольцом расположены открытые коллатеральные проводящие пучки, соединённые между собой губчатой колленхимой, и образующие вместе жёсткое (опорное) механическое кольцо (трубчатую структуру). Пучки в гистологическом (тканевом) строении состоят из (от периферии к центру): склеренхимы, первичной флоэмы, вторичной флоэмы, камбия, вторичной ксилемы и первичной ксилемы. Пучки по периметру окружены клетками обкладки (эндодермой). В центре стебля клетки основной паренхимы со временем разрушаются, образуя воздушную полость.
Вторичное строение стеблейдревесных семенных растений
Строение стеблей древесных растений характеризуется наличием в них одного концентрического открытого проводящего пучка. Рассмотрим строение стебля древесного растения на примере липы. В начале развития стебель снаружи покрыт эпидермисом, который сменяется перидермой. В состав перидермы входят пробка (феллема), феллоген и феллодерма. Под феллодермой перидермы находятся колленхима, основная паренхима коры и эндодерма, окружающая центральный проводящий пучок. Флоэма, расположенная под эндодермой в виде трапеций и разделённая вставками основной паренхимы, включает в себя первичную и вторичную флоэмы (ситовидные трубки) закреплённые склеренхимой (лубяными волокнами). Под флоэмой находится кольцо камбия, а ниже вторичная ксилема (древесина). Водопроводящая ксилема, расположенная под камбием у древесных растений, называется заболонь; опорная ксилема, не проводящая воду и расположенная вокруг сердцевины, называется ядровой древесиной. Под вторичной ксилемой находится кольцо первичной ксилемы, окружающее сердцевину стебля, образованную основной запасающей паренхимой.

Рис. 11. Концентрический тип строения стебля липы в поперечном разрезе (I) и схема строения стебля на разных уровнях (II):
А - срез на уровне появления прокамбия; Б - на уровне появления камбия; В - на уровне сформированной структуры. 1 - прокамбий, 2 - остатки эпидермы, 3 - пробка, 4 - колленхима, 5 - паренхима коры, 6 - эндодерма (4-6 - первичная кора), 7 - перициклическая зона, 8 - первичная флоэма, 9 - твердый луб, 10 - мягкий луб (вторичная флоэма), 11 - сердцевинный луч (7-11 - вторичная кора), 12 - камбий, 13 - осенняя древесина, 14 - весенняя древесина (13-14 - годичное кольцо древесины), 15 - вторичная древесина, 16 - первичная древесина (15-16 - древесина), 17 - перимедуллярная зона, 18 - основная паренхима (17-18 - сердцевина, 7-18 - центральный цилиндр).
Стебель сосны по своей конструкции аналогичен строению стебля липы – древесного покрытосеменного растения. Отличия состоят в том, что ксилема образована водопроводящими клетками – трахеидами, а флоэма – ситовидными клетками. В основной паренхиме коры и древесине находятся смоляные ходы. Снаружи стебель, как и у липы, покрыт перидермой.

Рис. 11. Поперечный срез стебля сосны
Лист – периферийный орган побега, выполняющий преимущественно фотосинтетическую, транспирационную и газообменную функции. Листья закладываются в узлах почки в виде примордиальных зачаточных бугорков в периферийной меристеме. Развиваясь из примордия, у листа формируется листовая пластина, соединённая со стеблем черешком (черешковые, влагалищные листья) или напрямую без него (сидячие, стеблеохватывающие, низбегающие).

Рис. 12. Морфологическое строение листьев.
Анатомическое строение листовой пластинки однодольного растения ириса.
Снаружи лист ириса покрыт эпидермисом. Устьица в эпидермисе расположены параллельными рядами с двух сторон листа. Внутренняя часть листа под эпидермисом заполнена губчатой хлоренхимой с большими межклетниками, в которой параллельно проходят закрытые коллатеральные проводящие пучки. Пучки окружены фильтрационными клетками обкладки (эндодермой). Пучки листьев являются продолжением проводящей системы стеблей и состоят из первичной флоэмы, зажатой между первичной ксилемой (с внутренней стороны) и склеренхимой (с наружной стороны). Пучки крепятся к эпидермису колленхимой.

Рис. 13. Поперечный срез листа однодольного растения
Анатомическое строение листовой пластинки двудольного растения камелии.
Снаружи лист покрыт эпидермисом. Устьица в эпидермисе расположены только с нижней (наружной) стороны листа. Внутри листа к наружному (нижнему) эпидермису прилегает губчатая хлоренхима, к внутреннему (верхнему) двумя рядами столбчатая хлоренхима (несущая основную фотосинтетическую нагрузку). Проводящая система жилок листа представляет собою закрытые коллатеральные проводящие пучки, прикреплённые к эпидермису колленхимой. В пучках, окруженных клетками обкладки (эндодермой), первичная флоэма зажата между внутренней первичной ксилемой и опорной наружной склеренхимой.

Рис. 14. Поперечный срез листа двудольного растения
Анатомическое строение листовой пластинки голосеменного растения сосны.
Лист сосны называется хвоя, имеет вытянутую иглоподобную (линейную) форму. Снаружи лист покрыт эпидермисом, под которым расположено кольцо механической ткани колленхимы (гиподермы). Устьица в эпидермисе расположены рядами вдоль листа. Мезофилл листа – внутренняя его часть, заполнена складчатой хлоренхимой, в которой по периметру расположены смоляные ходы. В середине листа находится центральный проводящий пучок, окружённый клетками обкладки (эндодермой). В основной паренхиме, заполняющей проводящий пучок, расположены два закрытых коллатеральных проводящих тяжа состоящих из трахеид первичной ксилемы сверху и ситовидных клеток первичной флоэмы снизу. Проводящие тяжи под флоэмой соединены между собой склеренхимой, дающей опору всему проводящему пучку.

Рис. 15. Поперечный срез листа сосны
Морфологически листья растений отличаются между собой по степени (порядку) сложности, форме листовых пластинок (рис. 16), степени их рассечения (рис. 17), форме верхушек, основ и краёв листовых пластинок (рис. 18-1,2), типу жилкования (рис. 19). По этим и другим морфологическим особенностям растения систематизируют и определяют их виды и разновидности.
По сложности различают: простые листья – имеющие одну листовую пластинку (клён, морковь), и сложные – несущие несколько листовых пластинок на одном общем черешке – рахисе (каштан, земляника). В сложном листе каждая листовая пластинка опадает отдельно.
По членению листовых пластинок листья делят на тройчатые, пальчатые и перистые. По глубине расчленения выделяют лопастные, раздельные и рассеченные листья (рис. 17).

Рис. 16. Формы листовых пластинок по очертанию: 1 – широкояйцевидный, 2 – округлый, 3 – обратноширокояйцевидный, 4 – яйцевидный, 5 – эллиптический, 6 – обратнояйцевидный, 7 – узкояйцевидный, 8 – ланцетный, 9 – удлиненный, 10 – обратноузкояйцевидный, 11 – линейный
|
| Рис. 17. Формы листовых пластинок по степени расчленения 1 – тройчато-лопастный, 2 – пальчато-лопастный, 3 – перисто-лопастный, 4 – тройчато-раздельный, 5 – пальчато-раздельный, 6 – перисто-раздельный, 7 – тройчато-рассечённый, 8 – пальчато-рассечённый, 9 – перисто- рассечённый. | |
| Рис. 18-1. Формы основ листовых пластинок (а): 1 – округлая, 2 – клиновидная, 3 – сердцевидная, 4 – стреловидная, 5 – копьевидная; формы верхушек листовых пластинок (б): 1 – тупая, 2 –острая, 3 – заостренная, 4 – остроконечная, 5 – выемчатая |
| |
|
| ||
| Рис. 18-2. Формы краёв листовых пластинок: а – цельнокрайняя; б – зубчатая; в – пильчатая; г – двоякопильчатая; д – городчатая; е – выемчатая; є – извилистая; ж – реснитчатая | ||
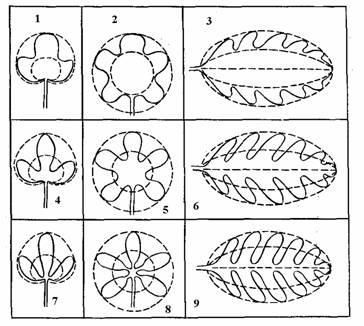



Рис .19. Жилкование листков: 1 – параллельное, 2 – дуговое,
3 – пальчатосетчатое, 4 – перистосетчатое, 5 – дихотомическое
Генеративные побеги семенных растений представляют собой специализированные части растений, на которых происходит формирование половых органов с половыми клетками.
К генеративным побегам голосеменных растений относятся мужские и женские шишки, покрытосеменных растений – цветки. Строение шишек сосны обыкновенной и цветка сои обыкновенной подробно будут рассмотрены в циклах развития этих растений.
Цветок – генеративный побег (орган размножения) покрытосеменных растений, состоящий из цветоноса, его расширенной части – цветоложа, на котором растут четыре типа цветолистиков:
Чашелистики (Са) – образуют чашечку цветка;
Лепестки (Со) – формируют венчик цветка. Венчик вместе с чашелистиками составляют двойной околоцветник цветка. Если в цветке нет лепестков (свекла) или чашелистиков (тюльпан), околоцветник называется простым (Р).
Тычинки (А) – формируют андроцей цветка; тычинки состоят из тычиночной нити и пыльника. В пыльниках из спор формируется пыльца.
Плодолистики (G) – в цветке срастаются в пестик (гинецей), в котором выделяют три части: рыльце, столбик и завязь. В завязи формируются семенные зачатки, которые расположены по стыковым краям плодолистиков.

Рис. 20 Строение цветка
Цветки растений можно описать формулой или диаграммой (рисунком). В формулах цветков знаками указывается пол цветка и симметрия его околоцветника, буквами обозначаются цветолистики, а их количество цифрами. Элементы цветка (цветолистики), которые срастаются между собой, обозначаются цифрой, взятой в скобки, а их расположение в несколько рядов – через знак плюс. Например, формула цветка тюльпана описывается формулой: ×Са0Со3+3А3+3G(3); где × – обозначает симметричный цветок, Са0 – отсутствие чашелистиков, Со3+3 – расположение лепестков по три в двух рядах, А3+3 – расположение тычинок по три в двух рядах, G(3) – пестик образован тремя сросшимися плодолистиками, черта под цифрой обозначает верхнюю завязь.
Для полного описания строения цветков их классифицируют:
– по полу:
Обоеполые – в цветке есть и тычинки, и пестик (в формулах цветков специальным знаком часто не обозначается);
Мужские (♂) – в цветке образуются только тычинки, без пестика;
Женские (♀) – цветок содержит только пестик, без тычинок.
– по симметрии околоцветника:
Симметричные (актиноморфные) (×) – через околоцветник можно провести две и больше осей симметрии (вишня);
Зигоморфные (↑) – через околоцветник можно провести только одну ось симметрии (фасоль, шалфей);
Асимметричные (ϟ) – через околоцветник нельзя провести ни одной оси симметрии, все лепестки (чашелистики) разные (зигокактус).
– по размещению в цветке завязи относительно других цветолистиков:
С верхней завязью (черта под цифрой) – завязь расположена выше остальных цветолистиков;
Со средней завязью (черта за цифрой) – завязь погружена в чашечку, как в бокал, но не срастается с ней;
С нижней завязью (черта над цифрой) – завязь погружена в цветоложе, остальные цветолистики расположены выше завязи.
С полунижней завязью (черта за цифрой) – завязь погружена в цветоложе наполовину.
 1 1
|  2 2
|  3 3
|  4 4
|
Рис. 21. Типы завязи в цветках: 1 – верхняя, 2 – средняя, 3 – нижняя, 4 – полунижняя
По способу срастания плодолистиков в пестике гинецеи делят на:
1) Апокарпный – образован одним замкнутым плодовым листом с креплением семязачатков по брюшному шву завязи пестика.
2) Ценокарпный – образован несколькими сросшимися плодовыми листами. Выделяют три разновидности ценокарпных гинецеев:
а) Синкарпный – плодолистики срастаются боковыми частями, семязачатки расположены в центральной части пестика по углам плодолистиков (томат),
б) Лизокарпный – происходит из синкарпного гинецея, боковые перегородки плодолистиков по мере развития плода растворяются, образуя центральную колонну, несущую семязачатки (семена) и оболочку плода (перец).
в) Паракарпный – плодолистики срастаются краями, семязачатки крепятся по швам срастания плодолистиков (огурец, мак).
 1 1
|  2 2
|  3 3
|  4 4
|
Рис. 22. Типы гинецеев в цветках: 1 – апокарпный,
2 – синкарпный, 3 – лизокарпный, 4 – паракарпный
Цветки на растениях могут располагаться на цветоносах одиночно или по несколько штук, формируя соцветия. По типу нарастания и направлению раскрытия цветков соцветия делят на:
Рацемозные, или ботрические – у которых, выделена гласная ось соцветия, нарастающая неограниченно долго (моноподиальный тип нарастания) и раскрытие цветков происходит снизу вверх (акропетально).
Цимозные – при развитии которых каждая ось на вершине заканчивается цветком и рано прекращает рост; её перерастают боковые оси (ветви соцветия: одна в монохазии, две в дихазии, несколько в плейохазии), также заканчивающиеся цветками (симподиальный тип нарастания). Цветение начинается с главного цветка и распространяется на цветки последовательных порядков «волнами» (рис. 25).
По степени разветвления соцветия разделяют на:
Простые – на главной оси располагаются одиночные цветки, таким образом, ветвление не превышает двух порядков (например, черёмуха, подорожник).
Сложные – главная ось несёт частные соцветия, то есть ветвление достигает трёх, четырёх и более порядков (например, сирень, калина и др.). В сложных соцветиях могут комбинироваться разные типы простых соцветий, например, колоски, собранные в кисть (камыш), корзинки, собранные в щиток (тысячелистник) и др.

Рис. 23. Простые рацемозные соцветия: 1 – кисть (смородина), 2 – колос (подорожник), 3 – початок (кукуруза, рогоз), 4 – зонтик (лук), 5 – головка (клевер), 6 – корзинка (одуванчик), 7 – щиток (груша)
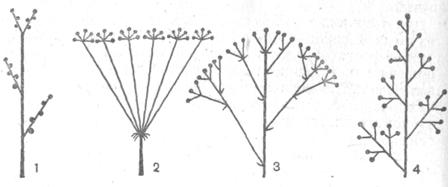
Рис. 24. Сложные рацемозные соцветия: 1 – сложный колос (пшеница), 2 – сложный зонтик (укроп), 3 – зонтики, собранные в метёлку (аралия), 4 – метёлка, или сложная кисть (виноград)

Рис. 25. Цимозные соцветия: 1 – монохазий завиток (белена),
2 – монохазий извилина (петуния), 3 – дихазий (бегония), 4 – плейохазий (молочай)
Соцветия различного типа, у которых главная ось является повислой, называются серёжками. Среди серёжек встречаются тирсы (ольха, берёза, лещина), простые кисти и колосья. Соцветие тирс представляет собою сложное соцветие с моноподиально нарастающей главной осью и боковыми частными соцветиями-цимоидами (березовые).
Плод – орган распространения, развивающийся из пестика цветка и в некоторых случаях части цветоложа. Плод состоит из околоплодника, развивающегося из стенок завязи, и семян, сформированных из семенных зачатков. В околоплоднике выделяют наружный, средний и внутренний слои, образованные в каждом отдельном случае разными тканями. Например, в костянке вишни наружный слой – экзокарп, представлен кожицей эпидермиса, средний слой – мезокарп – сочной запасающей паренхимой, внутренний слой – эндокарп – косточкой, защищающей семя, образованной склереидами.
Различают следующие типы плодов:
1) Ореховидные односемянные с сухим околоплодником:
а. Зерновка – паракарпный односемянный плод, у которого срастаются плодовая и семенная оболочки (рожь, рис, ячмень),
б. Семянка – паракарпный односемянный плод, у которого семенная и плодовая оболочки не срастаются (подсолнечник, одуванчик),
в. Орех – синкарпный односемянный плод с одревесневшим околоплодником (лещина, дуб),
г. Орешек – синкарпный односемянный плод с тонким одревесневшим околоплодником (гречиха, осока),
д. Крылатка – паракарпный односемянный плод с крыловидным выростом (клён, ясень);
2) Коробочковидные многосемянные с сухим околоплодником:
а. Листовка – апокарпный многосемянный плод, раскрывающийся по брюшному шву (живокость, дельфиниум),
б. Боб – апокарпный одно-, многосемянный плод, раскрывающийся по брюшному шву и спинной жилке (фасоль, горох),
в. Стручок – синкарпный многосемянный плод, раскрывающийся двумя створками с семенной пластинкой между ними (капуста, редис),
г. Стручочек – укороченный стручок (яснотка, пастушья сумка),
д. Коробочка – паракарпный плод (мак, белена);
3) Ягодовидные многосемянные с сочным околоплодником:
а. Ягода – синкарпный или лизокарпный многосемянный плод (картофель, перец),
б. Яблоко – апокарпный многосемянный плод-многолистовка, образованный нижней завязью, сросшейся с цветоложем (груша, айва),
в. Тыквина – паракарпный многосемянный плод (кабачок, огурец, дыня),
г. Померанец (гесперидий) – синкарпный плод, близкий к ягоде (лимон, мандарин).
4) Костянковидные односемянные с сочным или сухим околоплодником:
а. Костянка – апокарпный (ценокарпный) односемянный плод (слива, персик, миндаль).
Соплодие – совокупность зрелых плодов, образовавшихся в пределах одного соцветия, чётко обособленная от побега и других соплодий (шелковица, свёкла, рябина, калина, виноград).
Соплодие следует отличать от сборного плода, который возникает из одного цветка, а не из соцветия; отдельные части сборного плода – «плодики» – есть части плода, развившиеся из завязей обособленных плодолистиков (малина, пион, клён, укроп), а части соплодия – это настоящие плоды, развившиеся из завязей всего цветка.
Семена являются органом распространения и продолжительного сохранения, формируются в завязи цветков из семенных зачатков. Состоят из семенной защитной оболочки, зародыша (почки, стебля, корня, семядолей) и запасающей ткани. У однодольных растений развита только одна семядоля, прилегающая к запасающей ткани – эндосперму. Двудольные растения в зародыше имеют две равновеликие семядоли, а запас питательных веществ может содержаться как в самих семядолях (горох, огурец), так и в запасающей ткани, образованной из семенного зачатка, – перисперме, или из зародышевого мешка – эндосперме.

1 2
Рис. 26. Строение семян однодольных и двудольных растений: 1 – пшеницы, 2 – фасоли
Прорастание семян –это физиологический процесс развития зародыша семени во взрослое растение, связано с трансформацией запасных питательных веществ в ростовые и структурные компоненты клеток развивающегося организма. Прорастание семян происходит при:
а) физиологической зрелости семян (которые полноценно сформировались и прошли период покоя),
б) наличии достаточного для набухания и прорастания количества воды,
в) доступе кислорода к прорастающему семени,
г) оптимальной для прорастания температуре.
Первым этапом прорастания семян является набухание (впитывание семенем воды). При доступе кислорода и оптимальной температуре запасные питательные вещества подвергаются разложению на простые, с образованием из них компонентов, необходимых для деления клеток меристем зародыша, их роста и специализации в формирующемся теле растения. Первым в рост идёт зачаточный корешок зародыша. После его закрепления в субстрате (почве) он начинает всасывать воду и подавать её по ксилеме в побег. Вследствие ростовых процессов стебель побега выносит почку на поверхность почвы, где начинают разворачиваться и зеленеть (фотосинтезировать) листочки и стебель побега, переводя растение на автотрофное питание.
У пшеницы и других однодольных растений прорастающий зачаточный корешок защищен специальным чехлом – колеоризой, которую главный корень вскоре разрывает. Вслед за главным корнем у пшеницы развиваются два придаточных, которые вскоре догоняют его в росте. Три первых корня функционируют недолго, им на смену вырастают другие дополнительные корни, формируя мочковатую корневую систему. У кукурузы формируется смешанная корневая система, в которой главный корень сохраняется, но по форме не отличается от придаточных корней. Первый настоящий лист побега однодольных растений при прорастании в почве защищает колеоптиль, ненастоящий лист в виде чехла. При выходе на поверхность почвы колеоптиль в апикальной части разрушается, и первый настоящий лист выходит из колеоптиля и разворачивается.

Рис. 27. Прорастание семян пшеницы (А), кукурузы (Б), лука (В) и строение их проростков:
З – зерновка, К – главный корень, Клп – колеоптиль, Клр – колеориза, ПК – придаточные корни, Ст – стебель, Щ – щиток (развитая семядоля), Э – эндосперм, Эп – эпикотиль
При прорастании семян двудольных растений первым в рост также идёт главный корень, после чего трогается в рост стеблевая часть побега. У растений, которые выносят семядоли на поверхность почвы, удлинняется подсемядольная часть стебля – гипокотиль (соя, тыква), у растений, семядоли которых остаются в почве, удлинняется надсемядольная часть стебля – эпикотиль. При прорастании стеблевая часть проростка (эпикотиль или гипокотиль) выгибается дугой, разворачивая почку вниз, что предотвращает её повреждение. При выходе на поверхность почвы стебель выпрямляется, выводя почку в вертикальное положение.

Рис. 28. Прорастание семян фасоли (А) и гороха (Б)
Систематика живых организмов
Размножение организмов
Размножение организмов представляет собою их самовоспроизведение с увеличение их в количестве. Распространение – это размещение организмов в жизнеобеспечивающем пространстве (на определённой площади).
Растения размножаются:
вегетативно – вегетативными (ростовыми) органами или их частями: корнями, побегами;
бесполым путём – спорами – специализированными вегетативными клетками (споровые растения: мхи, хвощи, плауны, папоротники);
половым путём – семенами – органами, образованными растениями в результате оплодотворения женских гамет мужскими (семенные голосеменные и покрытосеменные растения).
Жизненный цикл растительных организмов представляет собой смену двух поколений: бесполого – спорофита и полового – гаметофита.

Рис. 29. Схема жизненного цикла высших растений
Спорофит – растение или часть растения, клетки которого имеют диплоидный (двойной – 2n) набор хромосом. Спорофит развивается из зиготы – первой диплоидной клетки. На спорофите формируется спорангий – орган, в котором из спорогенной ткани (2n) в результате мейоза образуются гаплоидные (n) споры.
Споры представляют собой клетки, предназначенные для распространения и длительного сохранения в неактивном состоянии (кроме спор семенных растений). В благоприятных условиях споры прорастают в гаметофиты.
Гаметофит – это растение или часть растения, клетки которого имеют гаплоидный (одинарный – n) набор хромосом. На гаметофитах образуются мужские и женские половые органы – гаметангии. Женский гаметангий споровых и голосеменных растений называется архегоний, покрытосеменных растений – зародышевый мешок. В женском гаметангии образуется женская половая клетка (гамета) – яйцеклетка. Мужской гаметангий споровых растений называется антеридий, у семенных растений мужской гаметангий одновременно является гаметофитом и представляет собой пыльцу. Мужские гаметангии формируют мужские половые клетки (гаметы) – сперматозоиды у споровых растений и спермии у семенных растений.
Слияние, при определённых условиях, мужской и женской гамет (половых клеток) называется сингамией, половым процессом или оплодотворением, в результате которого образуется первая диплоидная клетка нового спорофита.
Таким образом, в жизненном цикле развития растений во время мейоза (формирования спор) происходит перемешивание (рекомбинация) генетической информации, а вовремя оплодотворения мужской гаметой яйцеклетки создаётся новая комбинация генетической программы развития организма. Разнообразие генетических программ в клетках организмов обеспечивает возможность адаптации видов растений к меняющимся условиям и выживание наиболее приспособленных экземпляров.
Дата добавления: 2018-08-06; просмотров: 4045; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!