Дыхание микроорганизмовБиологическое окисление (энергетический метаболизм)
Дыхание- биологический процесс переноса электронов через дыхательную цепь от доноров к акцепторам с образованием АТФ. В зависимости от того, что является конечным акцептором электронов, выделяют аэробное и анаэробное дыхание. При аэробном дыхании конечным акцептором электронов является молекулярный кислород (О2), при анаэробном- связанный кислород ( -NO3 , =SO4, =SO3).
Примеры.
 О2
О2
А  эробное дыхание донор водородаH2O
эробное дыхание донор водородаH2O
Анаэробное дыхание
н  итратное окислениеNO3
итратное окислениеNO3
(  факультативные анаэробы) донор водородаN2
факультативные анаэробы) донор водородаN2
с  ульфатное окислениеSO4
ульфатное окислениеSO4
(  облигатные анаэробы) донор водородаH2S
облигатные анаэробы) донор водородаH2S
По типу дыхания выделяют четыре группы микроорганизмов.
1.Облигатные (строгие) аэробы. Им необходим молекулярный (атмосферный) кислород для дыхания.
2.Микроаэрофилы нуждаются в уменьшенной концентрации (низком парциальном давлении) свободного кислорода. Для создания этих условий в газовую смесь для культивирования обычно добавляют CO2, например до 10- процентной концентрации.
3.Факультативные анаэробы могут потреблять глюкозу и размножаться в аэробных и анаэробных условиях. Среди них имеются микроорганизмы, толерантные к относительно высоким (близких к атмосферным) концентрациям молекулярного кислорода - т.е. аэротолерантные, а также микроорганизмы которые способны в определенных условиях переключаться с анаэробного на аэробное дыхание.
4.Строгие анаэробы размножаются только в анаэробных условиях т.е. при очень низких концентрациях молекулярного кислорода, который в больших концентрациях для них губителен. Биохимически анаэробное дыхание протекает по типу бродильных процессов, молекулярный кислород при этом не используется.
Аэробное дыхание энергетически более эффективно (синтезируется большее количество АТФ).
В процессе аэробного дыхания образуются токсические продукты окисления (H2O2- перекись водорода, -О2 - свободные кислородные радикалы), от которых защищают специфические ферменты, прежде всего каталаза, пероксидаза, пероксиддисмутаза. У анаэробов эти ферменты отсутствуют, также как и система регуляции окислительно- восстановительного потенциала (rH2).
Основные методы создания анаэробных условий для культивирования микроорганизмов.
1.Физический - откачивание воздуха, введение специальной газовой безкислородной смеси (чаще - N2- 85%, CO2- 10%, H2- 5%).
2.Химический - применяют химические поглотители кислорода, используют газрегенирирующие пакеты.
3.Биологический - совместное культивирование строгих аэробов и анаэробов (аэробы поглощают кислород и создают условия для размножения анаэробов).
4.Смешанный - используют несколько разных подходов.
Необходимо отметить, что создание оптимальных условий для строгих анаэробов- очень сложная задача. Очень непросто обеспечить постоянное поддержание безкислородных условий культивирования, необходимы специальные среды без содержания растворенного кислорода, поддержание необходимого окислительно- восстановительного потенциала питательных сред, взятие и доставка, посев материала в анаэробных условиях.
Существует ряд приемов, обеспечивающих более подходящие условия для анаэробов- предварительное кипячение питательных сред, посев в глубокий столбик агара, заливка сред вазелиновым маслом для сокращения доступа кислорода, использование герметически закрывающихся флаконов и пробирок, шприцев и лабораторной посуды с инертным газом, использование плотно закрывающихся эксикаторов с горящей свечой. Используются специальные приборы для создания анаэробных условий-анаэростаты. Однако в настоящее время наиболее простым и эффективным оборудованием для создания анаэробных и микроаэрофильных условий является система “Газпак” со специальными газорегенерирующими пакетами, действующими по принципу вытеснения атмосферного воздуха газовыми смесями в герметически закрытых емкостях.
В процессе метаболизма выделяют два вида обмена: 1) пластический (конструктивный): анаболизм (с затратами энергии), катаболизм (с выделением энергии); 2) энергетический обмен (протекает в дыхательных мезосомах): дыхание; брожение.
В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы, факультативные анаэробы и облигатные анаэробы. Для аэробов акцептором является кислород. Факультативные анаэробы в кислородных условиях используют процесс дыхания, в бескислородных – брожение. Для облигатных анаэробов характерно только брожение, в кислородных условиях наступает гибель микроорганизмов из-за образования перекисей, идет отравление клетки.
Облигатные аэробы(бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и размножаются только в присутствии кислорода. Используют кислород для получения энергии путем кислородного дыхания. Они подразделяются на: 1) строгие аэробы(менингококки, бордетеллы), которые растут при парциальном давлении атмосферы воздуха; 2) микроаэрофилы(листерии) растут при пониженном парциальном давлении атмосферного возхдуха.
Облигатные анаэробы(бифидобактерии, лактобактерии,клостридии)не используют кислород для получения энергии. Тип метаболизма у них бродильный. Они подразделяются на: 1) строгие анаэробы – микроорганизмы для которых молекулярный кислород токсичен; он либо убивает микроорганизмы, либо ограничивает их рост. Энергию строгие анаэробы получают маслянокислым брожением; 2) аэротолерантные микроорганизмы(молочнокислые бактерии) используют кислород для получения энергии, но могут существовать в его атмосфере. Энергию получают гетероферментативным молочнокислым брожением.
Факультативные анаэробы(пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы)способны расти и размножаться как в присутствии кислорода, так и в отсутствии его. Они обладают смешанным типом метаболизма. Процесс получения энергии у них может происходить кислородным дыханием в присутствии кислорода, а в его отсутствии переключаться на брожение. Различное физиологическое отношение микроорганизмов к кислороду связано с наличием у них ферментных систем, позволяющих существовать в атмосфере кислорода. В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись водорода Н2О2 и закисный радикал кислорода О2-. Для нейтрализации токсичных форм кислорода, микроорганизмы, способные существовать в его атмосфере, имеют защитные механизмы.
У облигатных аэробов и факультативных анаэробов накоплению закисного радикала кислорода препятствует фермент супероксиддисмутаза.
У аэротолерантных микроорганизмов накоплению закисного радикала кислорода препятствует высокая концентрация ионов марганца, перекись водорода разрушается ферментом пероксидазой.
У строгих анаэробов наличие фермента супероксиддисмутазы коррелирует с их устойчивостью к кислороду. Для культивирования строгих анаэробов создаются условия, позволяющие удалять атмосферный кислород: использование анаэростатов, добавление в питательные среды редуцирующих кислород веществ, например тиогликолята натрия, использование поглоттелей кислорода.
Покоящиеся клетки прокариот
При переходе к неблагоприятным условиям существования многие прокариоты образуют особые дифференцированные формы – покоящиеся клетки, основным назначением которых является обеспечение переживания вида в неблагоприятных условиях. Их общей морфологической особенностью являются утолщенные клеточные стенки, а также частое образование дополнительных поверхностных структур на фоне одновременной дегидратации и компактизации метаболического и генетического аппаратов. В результате этого покоящиеся клетки характеризуются чрезвычайно низким (фоновым) уровнем обмена веществ и энергии, но приобретают повышенную устойчивость к действию разнообразных повреждающих факторов: высоких и низких температур, кислотности среды, радиации и пр. При попадании же в благоприятные условия покоящиеся клетки прорастают с образованием активно метаболизирующих вегетативных клеток.
Покоящиеся формы прокариот можно условно разделить на несколько видов: эндоспоры, экзоспоры, цисты, акинеты.
Эндоспоры– это особый тип покоящихся клеток эубактерий, формирующихся внутри цитоплазмы родительской клетки и обладающих специфическими структурами: внутренней и наружной мембранами, кортексом и многослойными белковыми покровами. К спорообразующим относится большое число эубактерий, относящихся к 15 родам, и имеющих клеточную стенку типичную для грамположительных бактерий. У аэробных микроорганизмов процесс спорообразования лучше всего изучен у р. Bacillus, и у анаэробов р. Clostridium. Переход к процессу спорообразования является дефицит питательных веществ, накопление в среде продуктов метаболизма, действующих по принципу кворум-зависимых регуляторных факторов. В упрощенном виде у бациллюссубтилисспоруляция регулируется системой из сигнальногоолигопептида, фосфатазы и белковых факторов споруляции. При достижении высокой плотности популяции, олигопептид в большом количестве поступает в клетку, инактивирует фосфатазу,активирует таким образом белковые факторы споруляции и запускает программу споруляции.
Последующий процесс споруляции можно рассматривать как неравновеликое бинарное деление особого типа, при котором одна из образующихся клеток формирует отмирающий спорангий, а вторая превращается в покоящуюся эндоспору. Спорообразованию всегда предшествует репликация ДНК, часть которого компактизируется и в составе т.н. «ядерного тяжа» переходит в ту часть клетки, где в последующем и будет формироваться спора. Компактизированный материал теряет свою метаболическую активность на ранних стадиях спорообразования, а сам процесс формирования споры контролируется геномом спорангия.
Помимо разделения генетического материала, в клетке активно идут процессы дегидратации (в споре остается 15% воды), ДНК переходит из В-формы в А-форму, что существенно повышает устойчивость к УФ лучам, белки распадаются до специфичного для спор и отсутствующего у вегетативных клеток вещества – дипиколиновой кислоты (пиридин-2,6-дикарбоновой) кислоты. В ходе ее образования происходит также интенсивное поглощение клеткой ионов кальция, образующего с кислотой хелатное соединение, составляющее 10-15% сухого вещества зрелых спор. Визуально же это проявляется как блестящие тела на фоне темных клеток.
Далее начинается процесс обособления протопласта: ЦПМ впячивается вовнутрь как при делении, но потом протопласт обрастает ЦПМ родительской клетки вторым слоем. Между внутренней и внешней мембраной начинает формироваться кортекс – многослойныйпептидогликан, отличающийся высокой степенью поперечной сшивки. Вслед за этим поверх наружной мембраны синтезируются дополнительные многослойные «споровые покровы», состоящие из белков, липидов и гликолипидов. Белки эндоспор богаты цистеином и гидрофобными аминокислотами, что обеспечивает их дополнительную устойчивость, в частности радиорезистентность. Далее у некоторых бактерий формируется еще одна структура – «экзоспориум», окружающий спору в виде свободного чехла и имеющий характерное для каждого вида тонкое многослойное строение.
После завершения процесса спорообразования происходит лизис спорангия, после чего спора высвобождается в окружающую среду. Споры могут сохраняться в течении сотен лет в окружающей среде. При наступлении благоприятных условий происходит прорастание споры, проходя три стадии: активацию, инициацию и вырастание.
Экзоспорыфототрофных бактерий формируются в результате отпочковывания от одного из полюсов материнской клетки. Они не имеют каких-либо особенных поверхностных структур, однако клеточная стенка значительно плотнее и толще аналогичной структуры вегетативной клетки. Экзоспорымиксобактерий - миксоспоры – формируются в т.н. «плодовом теле» псевдоплазмодия, клетки переходят в покоящееся состояние, их клеточная стенка утолщается и окружается капсулой.
Цистыраспространенный тип покоящихся клеток, образуемые представителями разных групп эубактерий, виды Azotobacter и скользящие бактерии.При истощении пищевых ресурсов в цисту превращается вся палочковидная клетка, а не только какая-то ее часть, что сопровождается изменением ее морфологии. Происходящие изменения включают потерю жгутиков, накопление в цитоплазме гранул поли-бета-гидроксимаслянной кислоты и образованием дополнительных клеточных покровов. Между ЦПМ и КС – формируется «интина», а поверх КС – «экзина». Возникающие толстостенные шарообразные покоящиеся клетки – цисты – устойчивы к высушиванию, механическим нагрузкам и облучению, но не к дейтсвию высоких температур.
Акинеты- это особая форма сохранения у планктонных нитчатых цианобактерий, обладающая повышенной устойчивостью к ряду пониженным температурам, высушиванию, но не к повышенным температурам. В акинету превращается вся вегетативная клетка, которая начинает этот процесс с увеличения своих размеров и накопленияв цитоплазме гранул запасных веществ (гликогеновых, полифосфатных, цианофициновых), а также карбоксисом при одновременном уменьшении количества хлорофилла и фикобилиновых пигментов. Пептидогликановый слой КС утолщается, окружающий слизистый чехол уплотняется за счет полисахаридных фибрилл. Под действием силы тяжести акиеты опускаются на дно водоемов и перезимовывают, а весной прорастают, после чего молодые, снабженные газовыми везикулами вегетативные клетки всплывают к поверхности воды.
Заверщая рассмотрение данного вопроса, следует также указать, что многие бактерии способны некоторое время сохранятся в неблагоприятных условиях существования и без каких-либо явных признаков структурной дифференцировки. Однако в этом случае в них происходят существенные физиологические перестройки, заключающиесяв резком угнетении активности метаболических процессов, снижении содержании АТФ и абсолютного значения мембранного потенциала. В результате микроорганизмы впадают в так называемое «некультивируемое» состояние, аналогичное переходу в анабиоз.
Приложение:

Рис.4 Молодые, интенсивно делящиеся клетки Clostridiumsporopenitum. Видны делящиеся нуклеоиды. Окраска ядерного вещества по методу Романовского — Гимза. Увел. x3500.

Схема 1. Процесс спорообразования по данным люминесцентной микроскопии (объяснение в тексте).
Схема 2. Цикл развития спорообразующих бактерий: в — деление вегетативных клеток; 1-7 стадии спорообразования: 1 — образование осевого хроматинового тяжа; 2 — образование споровой перегородки (септы); 3 — «поглощение» протопластом материнской клетки септированного участка цитоплазмы с ядром; 4 — формирование кортекса; 5 — начало формирования оболочки споры; 6 — завершение образования оболочки и созревание споры; 7 — лизис материнской клетки и освобождение спор; с — свободная зрелая спора; п — прорастание споры. Темные извитые нити — ДНК, красным цветом обозначены мембраны вегетативной клетки и споры, голубым — цитоплазма, желтым — клеточная стенка, синим — оболочка споры.
Схема 3. Строение зрелой споры: с — сердцевина споры; цпм — цитоплазматическая (внутренняя) мембрана споры; вмс -внешняя мембрана споры; кс — зародышевый слой клеточной стенки; к — кортекс; цс — слой цитоплазмы между внешней мембраной споры и оболочкой; всо — внутренний слой споровой оболочки; нсо — наружный слой споровой оболочки; в — выросты на спорах; п — подушечка, прикрепляющаяся к споре; лс — линзовидная структура, состоящая из гранулярного вещества.
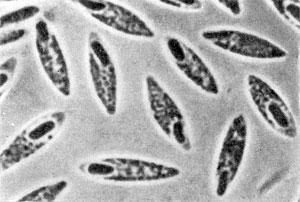
Клостридиальная форма клеток Clostridiumbutyricum. Увел. x3500.

Клостридиальная форма клеток. Споры снабжены светлыми конусовидными колпачками. Clostridiumspecies, шт. 1. Увел. x3500.

Клостридиальная форма клеток при спорообразовании. Споры со светлыми конусовидными колпачками. Clostridiumsp. Увел. x3500.
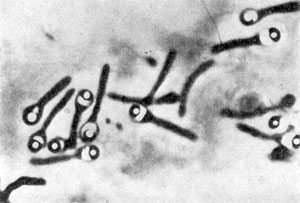
Плектридиальная форма клеток при спорообразовании. Споры с серповидными колпачками. Clostridiumsp. Увел. x3500.

Спорообразование у анаэробных бактерий: 1 — Clostridiumsporotrichum, инвагинация мембраны и образование септы в начале спорообразования: цпм — цитоплазматическая мембрана; вм — внутренняя мембрана проспоры; нм — наружная мембрана проспоры; мз — мезосомы; н — нуклеоид; м — инвагинирующие мембраны; 2 — Cl. sporofasciens, образование проспоры, окруженной двумя мембранами: нм — наружная мембрана проспоры; вм — внутренняя мембрана проспоры; кс — клеточная стенка: ят — ядерный тяж; 3 — Cl. sporotrichum, начало закладки споровой оболочки: о — листки оболочки; нм — наружная мембрана проспоры; ят — ядерный тяж; кс — клеточная стенка; вм — внутренняя мембрана проспоры; к — кортекс; 4 — Cl. penicillum, нуклеоид в проспоре: о — оболочка проспоры; к — кортекс; э — экзоспориум; цпм — цитоплазматическая мембрана; ят — ядерный тяж; кс — клеточная стенка материнской клетки; 5 — Cl. penicillum, завершение образования споровой оболочки, созревание сердцевины: кс — клеточная стенка материнской клетки; о — оболочка; э — экзоспориум: цпм — цитоплазматическая мембрана; к — кортекс; с — сердцевина споры.

Бациллярный тип спорообразования у Clostridiumsporopenatum. Увел. x3500

Спорообразующие анаэробные бактерии: 1 — Clostridiumpenicillum, трубчатые выросты на спорах; 2 — Cl. sporotrichum, жгутиковидные отростки на спорах; 3 — Cl. taeniosporum, лентовидные выросты на спорах; 4 — Cl. sporopenatum, перистые выросты на спорах; 5 — Cl. sartagoformum, трубчатые выросты на спорах; 6 — Clostridiumsp., споры с шипами на поверхности оболочки.

Ультратонкий срез клеток Cl. taeniosporum. Реакция на дегидрогеназы. Стрелками показаны отложения теллура. Видны выросты и светлые шаровидные включения гранулезы. Увел. x45 000 .

Стадии прорастания спор Cl. sporopenitum. Увел. x2100
Дата добавления: 2018-05-12; просмотров: 769; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!