Организм как открытая система. Теорема Пригожина.
Биологические объекты являются открытыми термодинамическими системами. Они обмениваются с окружающей средой энергией и веществом.
Живой организм – развивающаяся система, которая не находится в стационарном состоянии.
Основа функционирования живых систем – поддержание стационарного состояния при условии протекания диффузионных процессов, биохимических реакций, осмотических явлений. Если организм при изменении внешних условий не способен сохранить стационарное состояние, выходит из этого состояния, то это приводит к его гибели. Организм в этом случае не смог адаптироваться, т. е. не смог сравнительно быстро оказаться в стационарном состоянии, соответствующим условиям.
Принцип Пригожина:
В стационарном состоянии системы скорость возникновения энтропии вследствие необратимых процессов имеет минимальное значение при данных внешних условиях, препятствующих достижению системой равновесного состояния.
49. 12. Значение биологических мембран в процессе жизнедеятельности клетки
Клеточная мембрана (КМ) – это оболочка клетки, выполняющая следующие три основные функции:
барьерную – КМ обеспечивает избирательный (селективный), регулируемый пассивный и активный обмен веществом с окружающей средой;
матричную – КМ отвечает за определенное взаимное расположение и ориентацию мембранных белков для обеспечения их оптимального взаимодействия;
|
|
|
механическую – КМ обеспечивает прочность и автономность клетки и внутриклеточных структур.
Кроме трех основных функций, перечисленных выше, КМ выполняет и другие функции:
энергетическая – синтез АТФ на внутренних мембранах митохондрий и фотосинтез на мембранах митохондрий;
генерация и проведение биоэлектрических потенциалов;
рецепторная – в основе механической, обонятельной, зрительной химической и тепловой рецепции лежат процессы, происходящие на КМ.
50. 13. Молекулярная организация и модели клеточных мембран
Первая модельстроения биологических мембран (БМ) предложена Э. Овертоном в 1902г. Он решил, что БМ состоит из тонкого слоя фосфолипидов. В 1925 Гортер и Грендер проводили опыты по экстрагированию липидов из мембран эритроцитов и установили, что S монослоя липидов примерно в 2раза больше S поверхности эритроцитов. Так появилась билипидная модель БМ. В 1935 Даниелли и Девсон предложили “бутербродную” или “сэндвичную” модель БМ - липидные слои располагаются между 2-мя слоями белковых молекул. Современная модель строениябыла выдвинута в 1972 Сигнером и Никольсоном–жидкостно-мозаичная модель (основа БМ –двойной фосфолипидный слой, вклю-щий белки.)
|
|
|
Современная модель строения БМ была выдвинута в 1972 году Сингером и Никольсоном и получила название жидкостно-мозаичной модели.
Согласно этой модели, структурную основу мембраны составляет двойной фосфолипидный слой, включающий в себя белки. Мембранные белки бывают двух видов – периферические (поверхностные) и интегральные (внедренные в липиды). Схематично данная модель представлена на рис. 19.2 и 19.3.
Модели мембран (искусственные мембраны)
1) липосомы (фосфолипидные везикулы)-конструкции сфероподобной ф-мы, стенки к-рых образованы двойным липид.слоем
 2)плоские бислойные липидные мембраны(БЛМ)
2)плоские бислойные липидные мембраны(БЛМ) 
51. Физические свойства и параметры мембран
Приведем некоторые физические свойства и характеристики биологических мембран.
Толщина мембраны составляет примерно 8-10 нм.
Общая площадь всех мембран очень велика, например, печень крысы имеет массу 6 г, а общая площадь ее мембран достигает сотен квадратных метров.
Диаметр "ионных каналов" или пор составляет 0,35 - 0,8 нм.
Мембрана представляет собой диэлектрик с относительной диэлектрической проницаемостью от 2 до 6.
Электрическое сопротивление 1 см2 поверхности мембраны составляет 102 - 105 Ом, что в десятки миллионов раз больше сопротивления внеклеточной жидкости или цитоплазмы.
|
|
|
Вязкость мембран равна 30 - 100 мПа×с, что на два порядка выше вязкости воды и сравнима с вязкостью подсолнечного масла.
Поверхностное натяжение составляет 0,03 - 1 мН/м.
Липиды и белкив мембранах не являются статическими объектами, а участвуют в диффузионных процессах:латеральной диффузии – перемещение молекул в пределах плоскости мембраны;диффузии «флип-флоп» - перемещение молекул в направлении, перпендикулярном плоскости мембраны.
52.
53. Пассивный транспорт веществ и его разновидности. Математическое описание пассивного транспорта
Пассивный транспорт веществ через клеточные мембраны включает в себя следующие разновидности:простую диффузию и облегченную диффузию Фильтрация
Простая диффузия – процесс переноса вещества из области с большей концентрацией в область меньшей концентрации. Диффузия описывается уравнением Фика:
 ,
,
где 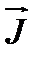 вектор плотности потока массы диффундирующего вещества,Сi и C0 – концентрации диффундирующего вещества внутри и вне клетки. Сmi и Cm0 – концентрации диффундирующего вещества внутри мембраны на границе мембрана.P– проницаемость мембраны.
вектор плотности потока массы диффундирующего вещества,Сi и C0 – концентрации диффундирующего вещества внутри и вне клетки. Сmi и Cm0 – концентрации диффундирующего вещества внутри мембраны на границе мембрана.P– проницаемость мембраны.
|
|
|
Облегченная диффузия.
Срособность мембран обусловлена двумя причинами: наличием в них переносчиков,называемыхионофорами (подвижныхификсированных) и каналов. Облегченная диффузия – процесс транспорта веществ с помощью специальных молекул-переносчиков.
Фильтрация.Фильтрация представляет собой перенос молекул растворителя под действием градиента давления. Таким образом, причиной и движущей силой в процессе фильтрации является разность давлений. Объем растворителя, перенесенного в результате фильтрации, определяется формулой Пуазейля:
 ,
,
где  - перепад давления на расстоянии
- перепад давления на расстоянии  ,
,  - вязкость жидкости,
- вязкость жидкости,  - радиус поверхности фильтрации,
- радиус поверхности фильтрации,  - время переноса.
- время переноса.
Фильтрация.Фильтрация представляет собой перенос молекул растворителя под действием градиента давления. Таким образом, причиной и движущей силой в процессе фильтрации является разность давлений. Объем растворителя, перенесенного в результате фильтрации, определяется формулой Пуазейля:
,
где - перепад давления на расстоянии , - вязкость жидкости, - радиус поверхности
жидкости.
54. Активный транспорт ионов. Механизм активного транспорта вещ-в на примере Na-K насоса. Если бы в клетках сущ. только пассивный транспорт, то конц-ции, давления и др. величины вне и внутри клетки сравнялись бы. Поэтому сущ-т др. механизм, работающий в направлении против электрохимического градиента и происходящий с затратой энергии клеткой. Перенос молекул и ионов против электрохимического градиента,осуществляемый клеткой за счет энергии метаболических процессов, наз.активным транспортом.Он присущ только биологическим мембранам. Активный перенос вещ-ва через мембрану происходит за счет свободной энергии,высвобождающейся в ходе хим. Реакций внутри клетки.Активный транспорт в организме создает градиенты концентраций, электр. потенциалов, давлений,т.е.поддерживает жизнь в организме.Изучены 3 основные системы акт.трансп., кот-ые обеспечивают перенос ионов Na,K,Ca,H через мембрану.Механизм. Ионы К+ и Na+ неравномерно распределены по разные стороны мембраны:концентр. Na+снаружи> ионов K+,а внутри клетки K+> Na+.Эти ионы диффундируют через мембрану по направлению электрохимического градиента,что приводит к его выравниванию. Na-K насосы вх. в состав цитоплазмат. мембран и работают за счет энергии гидролиза молекул АТФ с обр-ем мол-л АДФ и неорганич. фосфата Фн:АТФ=АДФ+Фн.Насос работает обра-тимо: градиенты конц-ций ионов способ-ют синтезу мол-л АТФ из мол-л АДФ и Фн: АДФ+Фн=АТФ.Насос переносит из клетки во внеш. среду 3 иона К+внутрь клетки.
55.Способы проникновения вещ-в через биологические мембраны. . Одной из важнейших хар-к клеточных мембран(КМ) явл-ся избират. проницаемость. КМ избирательно снижает скорость передвижения мол-л в клетку и из нее. Чем меньше мол-ла и чем меньше она обр. водород. связей,тем быстрее она диффун-дирует ч/з мембрану. =>, чем меньше мол-ла и чем более она жирорастворима, тем быстрее она будет проникать через мембрану.Малые неполярные мол-лы легко растворимы в липидах КМ и быстро диффундируют.Клетка была вынуждена создать спец.мех-мы для транспорта растворимых в воде вещ-в через мембрану-через поры в мембране и посредством транспортных белков-переносчиков мол-л. Для жиронерастворимых вещ-в и ионов мембрана выступает как молекулярное сито:чем больше размер частицы,тем меньше проницаемость мембраны для этого вещ-ва. Избирательность переноса обеспечивается набором в мембране пор определенного радиуса,соответствующих размеру проникающей частицы.Это распределение зависит от мембранного потенциала. Перенос малых водорастворимых мол-лосуществляется при помощи специальных транспортных белков.Это особые белки,каждый из которых отвечает за транспорт определенных мол-л или групп мол-л. За перенос сахара,аминокислот и др. полярных мол-л ответственны специальные мембранные транспортные белки.Каждый из них предназначен для определенного класса мол-л.Все они обеспечивают перенос мол-л через мембрану, формируя в ней сквозные проходы.Транспортные белки делятся набелки-переносчики, и каналообразующие белки.Переносчикивзаим-ют с молекулой переносимого вещ-ва и каким-либо способом перемещают ее сквозь мембрану. Каналообразующие-формируют в мембране водные поры,через кот-ые могут проходить вещ-ва. Отличия облегченной диффузии от простой:1)перенос ионов с участием переносчиков происх. значительно быстрее; 2)обладает св-вом насыщения- при ув. концентр. С одной стороны мемраны плотность потока вещ-ва возрастает лишь до некоторого предела. Разновидностью облегч.дифф.-транспорт с помощью неподвижных мол-л переносчиков,фиксированных поперек мембраны. Осмос-движение мол-л воды через полупроницаемые мембраны из мест с меньшей концентрацией растворенного вещ-ва в места с большей концентр. Осмос обусл-ет гемолиз эритроцитов в гипотонических растворах и тургор в растениях.
56. История открытия биопотенциалов. Гипотеза Бернштейна. . Фр.священник аббат Нолле в 1746г открыл явление осмоса.1826г. Дютрошедоказал,что осмос есть результат проявления не особых,мифических сил,а законов физики и химии. Немецк.ботаник Пфеффер-изобрел осмометр и измерил величину осмотического давления.Он обнаружил,что для каждого раствора величина давления пряма-пропорциональна концентр. раств.вещ-ва,не проходящего через полупроницаемую мембрану (Росм=m/V m-масса растворенного вещ-ва, V-объем раствора).Вант-Гофф пришел к заключению,что мол-лы растворенного вещ-ва в растворителе ведут себя подобно мол-ам идеального газа(Росм=СмRT См-молярная плотность растворенного вещ-ва).Теория Вант-Гоффа давала точные значения величины осм.давления для многих вещ-в,но для некоторых оказ. больше расчетной в 2раза.С.Аррениуспредположил что в р-ре мол-ла соли распадается на 2 частицы-электролитич. диссоциация.Вальтер Нернст обосновал идею диффузного потенциала, возн-го при соприк-нии двух жидкостей. Вел-на дифф.потенц. рассчит-ся: Фн=(u-v/u+v)×(RT/F)×Ln(C1/C2),где uи v-скорости быстрого и медл. ионов, R-газ.постоянная,C1 и С2-конц-ции электролита. Для возн-ния дифф.потенц. нужна разность конц-ций электролита, различная подвиж-ть анионов и катионов. Бернштейн начал объяснять электрич св-ва мышц не устройством этих органов в целом,а свойствами клето,из которых эти органы состояли.1902г-год рождения мембранной теории биопотенциалов.Согласно гипотезе Бернштейна, каждая клетка им. оболочку,кот-ая представляет собой полупроницаемую мембрану. Внутри и вне клетки имеется много свободных ионов,среди кот-ых нах-ся ионы K+. Разность пот-лов между внутр. стороной БМ и ее наружной стороной наз-ся потенциалом покоя (ПП).Величина ПП опис-ся формулой Нернста: ФН=-(RT/F)×Ln[(K+)I/(K+)0] где [K+]i-конц-ция ионов К внутри клетки,[K+]0-конц-ция ионов К снаружи клетки.
57.Мембранно-ионная теория генерации биопот-лов клеткой и основ. опыты, ее подтвер-щие. Ю.Бернштейн (опыты на мышце лягушки). Нагрев 1 конца целой мышцы, от нагретого участка к холодному потечет ток. Электрич. ток течет по направлению от точек пространства с более высоким потенциалом к месту с более низким значением электрического потенциала.В 1905г. Геберобнаружил,что все соли,сод-е К, оказ. на мышцу схожее д-вие: участок на кот-ый действовал раствор соли К, приобретал отрицательный потенциал по отношению к другим участкам мышцы.Все соли К при диссоциации в воде повышали наружную конц-цию ионов К, при этом отношение (K+)i\(K+)0 умен-ся, ум-ся и пот-ал той области мышцы, на кот. д-ют соли К. Однако эксперименты Бернштейна и Гебера были косвенными. Чтобы подтвердить правильность гипотезы,требовалось доказать следующее:1)клетки им. мембрану,кот-ая проницаема лишь для одного иона;2)конц-ция этого иона по обе стороны БМ различная;3)потенциал на мембране возникает только за счет проницаемости мембраны для этого иона и он равен нернстовскому потенциалу. В 1936г.Дж.Юнг обнаружил кольмара,у кот-ого диаметр нервного волокна доходил до миллиметра.Аксон кальмара был гигантской клеткой,хотя сам моллюск не был гигантом.Нервное волокно выняли из моллюска и поместили в морскую воду,и оно не погибло.=>(эксперимент на клет.уровне). В 1939г. А.Ходжкин и Хаксли измерили разность потенциалов на аксоне кальмара.Они доказали,что внутри аксона им. много ионов К ,и они обр. ионный газ,т.е.нах-ся в свобод. сост-нии.
58. Потенциал покоя. Уравнение Нернста. Уравнение Гольдмана-Ходжкина-Катца
Потенциал покоя (ПП, мембранный потенциал покоя). ПП называется разность потенциалов между внутренней и наружной сторонами клеточной мембраны, возникающей в состоянии покоя клетки. У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует отрицательная разность потенциалов (ПП) порядка 60—90 мВ, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной. ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К+.
ПП можно рассчитать с помощью формулы Нернста:
 ,
,
где  - концентрация ионов калия внутри клетки,
- концентрация ионов калия внутри клетки,  - концентрация ионов калия снаружи клетки.
- концентрация ионов калия снаружи клетки.
 точные значения для ПП вычисляются по формуле Гольдмана-Ходжкина-Катца (ГХК), которая учитывает проницаемость КМ в покое не только для ионов калия, но и для ионов натр ия и хлора:
точные значения для ПП вычисляются по формуле Гольдмана-Ходжкина-Катца (ГХК), которая учитывает проницаемость КМ в покое не только для ионов калия, но и для ионов натр ия и хлора: 
где PK, PNa, PCl – проницаемость КМ для ионов калия, натрия и хлора
59. Механизм генерации потенциала действия
Потенциал действия (ПД)
Все раздражители, действующие на клетку, вызывают в первую очередь снижение ПП, когда оно достигает критического значения (порога), возникает активный распространяющийся ответ — ПД. Во время восходящей фазы ПД кратковременно изменяется потенциал на мембране: её внутренняя сторона, заряженная в покое электроотрицательно, приобретает в это время положительный потенциал. Достигнув вершины, ПД начинает падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню, близкому к исходному, — ПП.
Полное восстановление ПП происходит только после окончания следовых колебаний потенциала — следовой деполяризации или гиперполяризации, длительность которых обычно значительно превосходит продолжительность пика ПД. Согласно мембранной теории, деполяризация мембраны, вызванная действием раздражителя, приводит к усилению потока Na+ внутрь клетки, что уменьшает отрицательный потенциал внутренней стороны мембраны — усиливает её деполяризацию. Это, в свою очередь, вызывает дальнейшее повышение проницаемости для Na+ и новое усиление деполяризации и т.д. В результате такого взрывного кругового процесса, т. н. регенеративной деполяризации, происходит изменение мембранного потенциала, характерное для ПД.
Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110—120 мВ. Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, — 0,3—0,4 мс, у волокон же мышц сердца — 50—600 мс. В растительных клетках пресноводной водоросли хара ПД продолжается около 20 с.
60. Распространение потенциала действия по миелиновым и безмиелиновым нервным волокнам
нервного волокно представляет собой цилиндр, боковую поверхность которого образует мембрана, отделяющая внутренний раствор электролита от наружного. Это придает волокну свойства коаксиального кабеля, изоляцией которого служит клеточная мембрана. Но нервное волокно — очень плохой кабель. Сопротивление изоляции этого живого кабеля примерно в 105 раз меньше. Было установлено, что уменьшение электрического потенциала на мембране нервного волокна по мере удаления от источника возбуждения убывает по экспоненциальному закону
 ,
,
где j0 – значение потенциала в точке возбуждения, jl –значение потенциала в точке, расположенной на расстоянии l от источника возбуждения, l - константа длины нервного волокна, равная расстоянию, на котором величина потенциала убывает в. Константа l зависит от удельного электрического сопротивления оболочки нервного волокна rm, удельного электрического сопротивления цитоплазмы ri, радиуса нервного волокна r:
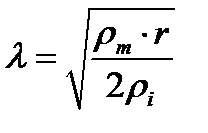
Чем больше l, тем больше скорость распространения нервного возбуждения. Как следует из приведенной выше формулы, l тем больше, чем больше радиус нервного волокна и чем больше удельное электрическое сопротивление мембраны нервного волокна.
Рассмотрим процесс распространения возбуждения по безмиелиновым нервным волокнам.При возбуждении участка нервного волокна происходит деполяризация данного участка. При деполяризации с наружной стороны мембран возникает отрицательный электрический потенциал, а с внутренней стороны – положительный потенциал. Между возбужденным и невозбужденным участками нервного волокна возникнут локальные электрические токи, так у возбужденного участка внутренняя поверхность имеет положительный заряд, а у невозбужденного участка – отрицательный электрический заряд и между ними возникает разность потенциалов. По поверхности нервного волокна локальный электрический ток течет от невозбужденного участка к возбужденному, внутри волокна электрический ток течет в обратном направлении – от возбужденного участка к невозбужденному.
Дата добавления: 2018-05-12; просмотров: 811; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!