Продукция и энергия в экосистемах
Экосистема может обеспечить круговорот веществ только в том случае, если включает 4 необходимые для этого части: 1) запасы биогенных элементов; 2) продуценты; 3) консументы; 4) редуценты. На их сложном и постоянном взаимодействии основан первый (основной) принцип функционирования экосистем:получение ресурсов и избавление от отходов происходят в рамках круговорота всех элементов.
Данный принцип гармонирует с законом сохранения массы. Так как атомы не возникают, не исчезают и не превращаются один в другой, они могут использоваться бесконечно, переходя в самые различные химические соединения. Именно это и происходит в природных экосистемах.
Необходимо подчеркнуть, однако, что биологический круговорот веществ (точнее элементов) есть результат деятельности организмов, которым для этого требуются постоянные энергетические затраты, компенсируемые Солнцем. Энергия солнечных лучей, поглощаемая зелеными растениями, в отличие от химических элементов, не может использоваться организмами бесконечно. Данное заключение вытекает из второго закона термодинамики: энергия при превращении из одной формы в другую, т.е. при совершении работы, частично переходит в тепловую форму и рассеивается в окружающей среде.
Следовательно, каждый цикл круговорота, зависящий от активности организмов и сопровождаемый потерями энергии из них, требует все новых дотаций энергии. Отсюда существование экосистем любого ранга и вообще Жизни на Земле обусловлено постоянным круговоротом веществ, который, в свою очередь, поддерживается постоянным притоком солнечной энергии. В этом состоит второй основной принцип функционирования экосистем:они существуют за счет не загрязняющей среду и практически вечной солнечной энергии, количество которой относительно постоянно и избыточно.
|
|
|
В отличие от веществ, которые постоянно циркулируют по разным блокам экосистемы и всегда могут вновь входить в круговорот, поступившая энергия может быть использована только 1 раз и может идти только в потоке энергии.
Для изучения развития различных экосистем большой интерес представляет закон максимизации энергии(Г. Одум, Ю. Одум):в соперничестве с другими экосистемами выживает (сохраняется) та из них, которая наилучшим образом способствует поступлению энергии и использует максимальное ее количество наиболее эффективным способом.
Авторы данного закона указывают: «с этой целью система:
1) создает накопители (хранилища) высококачественной энергии (например, запасы жира);
2) затрачивает определенное количество накопленной энергии на обеспечение поступления новой энергии;
|
|
|
3) обеспечивает круговорот различных веществ;
4) создает механизмы регулирования, поддерживающие устойчивость системы и ее способность к приспособлению к изменяющимся условиям;
5) налаживает с другими системами обмен, необходимый для обеспечения потребности в энергии специальных видов».
Закон максимизации энергии справедлив и в отношении информации, следовательно (по Н.Ф. Реймерсу), его можно рассматривать более широко как закон максимизации энергии и информации:наилучшими шансами на самосохранение обладает система, в наибольшей степени способствующая поступлению, выработке и эффективному использованию энергии и информации.
Между организмами биоценоза возникают и устанавливаются прочные пищевые взаимоотношения или цепи питания. Те из них, которые начинаются с фотосинтезирующих организмов, называют цепями выедания(или пастбищными), а цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных, – детритными цепями.
Место каждого звена в цепи питания называют трофическим уровнем, он характеризуется различной интенсивностью протекания потока веществ и энергии. Первый трофический уровень всегда составляют продуценты (например, капуста); растительноядные консументы (заяц) относятся ко второму трофическому уровню; плотоядные, живущие за счет растительноядных форм консументов – к третьему; потребляющие других плотоядных – соответственно к четвертому и т.д. Вследствие этого различают консументов первого, второго, третьего и четвертого порядков, занимающих разные уровни в цепях питания. Следовательно, пищевая цепь – основной канал переноса энергии, заключенного в пище, в сообществе.
|
|
|
Согласно расчетам, на каждом этапе передачи вещества и энергии по пищевой цепи теряется примерно 90 % энергии (принцип Линдемана) и только около одной десятой доли ее переходит к очередному потребителю. Указанное соотношение в передаче энергии в пищевых связях организмов называют «правилом десяти процентов». Например, количество энергии, которое доходит до третичных плотоядных (пятый трофический уровень), составляет лишь около 10-4 энергии, поглощенной продуцентами. Этим объясняется ограниченное количество (5–6) звеньев (уровней) в пищевой цепи независимо от сложности видового состава биоценоза.
В экосистемах биологическую продукцию создают автотрофы и гетеротрофы. Биомасса, производимая автотрофами на единице площади в единицу времени, называется первичной продуктивностью. Гетеротрофы питаются готовым органическим веществом, а биомасса, создаваемая гетеротрофами, называется вторичной продуктивностью.
|
|
|
Биоценозы динамичны, в них постоянно происходят изменения в состоянии и жизнедеятельности их членов и соотношении популяций.
Изменение внешних условий среды влияет на некоторые виды неблагоприятно, другие же виды могут от этого, наоборот, выиграть. Изменившиеся условия позволяют включиться в экосистему новым видам.
Природные сообщества могут включать сотни и тысячи видов: от микроскопических бактерий до огромных деревьев и многотонных животных. Казалось бы, усложнение экосистемы, ее видового состава, должно негативно отражаться на устойчивости сообщества. Тем не менее, практические наблюдения полностью опровергают это предположение.
Экологические пирамиды
Для наглядности представления взаимоотношений между организмами различных видов в биоценозе принято использовать экологические пирамиды, различая пирамиды численности, биомасс и энергии.
Для построения пирамиды численности подсчитывают число организмов на некоторой территории, группируя их по трофическим уровням:
продуценты — зеленые растения;
первичные консументы — травоядные животные;
вторичные консументы — плотоядные животные;
третичные консументы — плотоядные животные;
п-е консументы («конечные хищники») — плотоядные животные;
редуценты — деструкторы.
Консументы второго, третьего и более высоких порядков могут быть хищниками (охотиться, схватывая и убивая жертву), могут питаться падалью или быть паразитами.
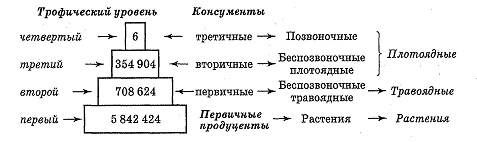
Рис. 17. Экологическая пирамида численности для луга, поросшего злаками: цифры — число особей
В последнем случае они по величине меньше своих хозяев, в результате чего пищевые цепи паразитов необычны по ряду параметров. В типичных пищевых цепях хищников плотоядные животные становятся крупнее на каждом трофическом уровне. Каждый уровень изображается условно в виде прямоугольника, длина или площадь которого соответствуют численному значению количества особей. Расположив эти прямоугольники в соподчиненной последовательности, получают экологическую пирамиду численности (рис. 17), основной принцип построения которой впервые сформулировал американский эколог Ч. Элтон.
Данные для пирамид численности получают достаточно легко путем прямого сбора образцов, однако существуют и некоторые трудности:
продуценты сильно различаются по размерам, хотя один экземпляр злака или водоросли имеет одинаковый статус с одним деревом. Это порой нарушает правильную пирамидальную форму, иногда давая даже перевернутые пирамиды; адиапазон численности различных видов настолько широк, что при графическом изображении затрудняет соблюдение масштаба, однако в таких случаях можно использовать логарифмическую шкалу.
Экологическую пирамиду биомасс строят аналогично пирамиде численности. Ее основное значение состоит в том, чтобы показывать количество живого вещества (биомассу — суммарную массу организмов) на каждом трофическом уровне. Это позволяет избежать неудобств, характерных для пирамид численности. В этом случае размер прямоугольников пропорционален массе живого вещества соответствующего уровня, отнесенной к единице площади или объема. Термин «пирамида биомасс» возник в связи с тем, что в абсолютном большинстве случаев масса первичных консументов живущих за счет продуцентов, значительно меньше массы этих продуцентов, а масса вторичных консументов меньше, чем первичных.
При отборе образцов определяют биомассу на корню или урожай на корню (т.е. в данный момент времени), которая не содержит никакой информации о скорости образования или потребления биомассы.
Скорость создания органического вещества не определяет его суммарные запасы, т.е. общую биомассу всех организмов каждого трофического уровня.
Одним из следствий являются «перевернутые пирамиды». Зоопланктон биоценозов озер и морей чаще всего обладает большей биомассой, чем его пища — фитопланктон, однако скорость размножения зеленых водорослей настолько велика, что в течение суток они восстанавливают всю съеденную зоопланктоном биомассу. Тем не менее, в определенные периоды года (во время весеннего цветения) наблюдают обычное соотношение их биомасс
Кажущихся аномалий лишены пирамиды энергий. Самым фундаментальным способом отражения связей между организмами разных трофических уровней и функциональной организации биоценозов является пирамида энергий, в которой размер прямоугольников пропорционален энергетическому эквиваленту в единицу времени, т.е. количеству энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за принятый период (рис. 18). К основанию пирамиды энергии можно обоснованно добавить снизу еще один прямоугольник, отражающий поступление энергии Солнца.
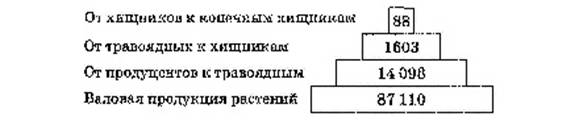
Рис. 18. Пирамида энергии: цифры — количество энергии, кДж • м-2• г -1
Пирамида энергий отражает динамику прохождения массы пищи через пищевую (трофическую) цепь, что принципиально отличает ее от пирамид численности и биомасс, отражающих статику системы.
На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей. Если учтены все источники энергии, то пирамида всегда будет иметь типичный вид (в виде пирамиды вершиной вверх), согласно второму закону термодинамики.
Пирамиды энергий позволяют не только сравнивать различные биоценозы, но и выявлять относительную значимость популяций в пределах одного сообщества. Они являются наиболее полезными из трех типов экологических пирамид, однако получить данные для их построения труднее всего.
Динамика экосистем.
Сложение экосистем - динамический процесс. В экосистемах постоянно происходят изменения в состоянии и жизнедеятельности их членов и соотношении популяций. Многообразные изменения, происходящие в любом сообществе, относят к двум основным типам: циклические и поступательные.
Циклические изменениясообществ отражают суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных ритмов организмов. Суточная динамика экосистем связана главным образом с ритмикой природных явлений и носит строго периодический характер. В каждом биоценозе имеются группы организмов, активность жизни у которых приходится на разное время суток. Одни активны днем, другие - ночью. Отсюда в составе и в соотношении отдельных видов биоценоза той или иной экосистемы происходят периодические изменения, так как отдельные организмы на определенное время выключаются из него. Суточную динамику биоценоза обеспечивают как животные, так и растения. У растений в течение суток изменяются интенсивность и характер физиологических процессов - ночью не происходит фотосинтез, нередко у растений цветки раскрываются только в ночные часы и опыляются ночными животными, другие приспособлены к опылению днем. Суточная динамика в биоценозах, как правило, выражена тем сильнее, чем значительнее разница температур, влажности и других факторов среды днем и ночью.
Более значительные отклонения в биоценозах наблюдаются при сезонной динамике. Это обусловлено биологическими циклами организмов, которые зависят от сезонной цикличности явлений природы. Так, смена времени года значительное влияние оказывает на жизнедеятельность животных и растений (спячка, зимний сон, диапауза и миграции у животных; периоды цветения, плодоношения, активного роста, листопада и зимнего покоя у растений). Сезонной изменчивости подвержена нередко и ярусная структура биоценоза. Отдельные ярусы растений в соответствующие сезоны года могут полностью исчезать, например, состоящий из однолетников травянистый ярус. Длительность биологических сезонов в разных широтах неодинакова. В связи с этим сезонная динамика биоценозов арктической, умеренной и тропической зон различна. Она выражена наиболее четко в экосистемах умеренного климата и в северных широтах.
Многолетняя изменчивость является нормальной в жизни любого биоценоза. Так, количество осадков, выпадающих в Барабинской лесостепи, резко колеблется по годам, ряд засушливых лет чередуется с многолетним периодом обилия осадков. Тем самым оказывается существенное влияние на растения и животных. При этом происходит выработка экологических ниш - функциональное размежевание в возникающем множестве или его дополнение при малом разнообразии.
Многолетние изменения в составе биоценозов повторяются и в связи с периодическими изменениями общей циркуляции атмосферы, в свою очередь, обусловленной усилением или ослаблением солнечной активности.
В процессе суточной и сезонной динамики целостность биоценозов обычно не нарушается. Биоценоз испытывает лишь периодические колебания качественных и количественных характеристик.
Поступательные измененияв экосистеме приводят в конечном итоге к смене одного биоценоза другим, с иным набором господствующих видов. Причинами подобных смен могут являться внешние по отношению к биоценозу факторы, действующие длительное время в одном направлении, например, увеличивающееся загрязнение водоемов, возрастающее в результате мелиорации иссушение болотных почв, усиленный выпас скота и т.д. Данные смены одного биоценоза другим называютэкзогенетическими.В том случае, когда усиливающее влияние фактора приводит к постепенному упрощению структуры биоценоза, обеднению их состава, снижению продуктивности, подобные смены называютдигрессивными или дигрессиями.
Эндогенетические сменывозникают в результате процессов, которые происходят внутри самого биоценоза.Последовательная сменаодного биоценоза другим называется экологической сукцессией(от лат. succession - последовательность, смена). Сукцессия является процессом саморазвития экосистем. В основе сукцессии лежит неполнота биологического круговорота в данном биоценозе. Известно, что живые организмы в результате жизнедеятельности меняют вокруг себя среду, изымая из нее часть веществ и насыщая ее продуктами метаболизма. При сравнительно длительном существовании популяций они меняют свое окружение в неблагоприятную сторону и как результат - оказываются вытесненными популяциями других видов, для которых вызванные преобразования среды оказываются экологически выгодными. В биоценозе происходит таким образом смена господствующих видов. Здесь четко прослеживаетсяправилоэкологического дублирования(рис. 19).
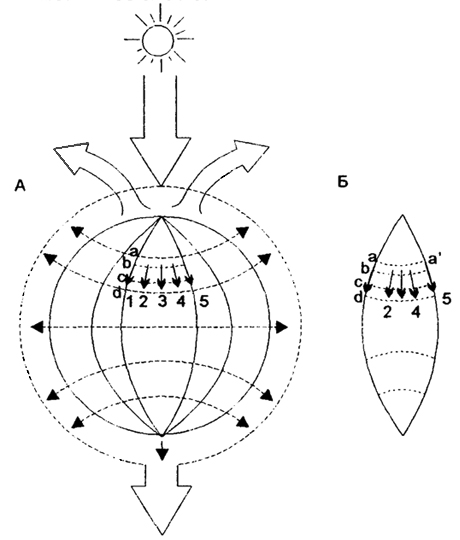
Рис. 19. Потоки энергии и механизм обеспечения надежности биотических систем в биосфере (no H.Ф. Реймерсу, 1994):
1, 2, 3... - потоки энергии через виды; а-а... - связи между ними, А - состояние до исчезновения вида; Б - вид 3 исчез, проходившие через него потоки энергии идут через дублирующие виды 2 и 4
Длительное существование биоценоза возможно лишь в том случае, если изменения среды, вызванные деятельностью одних живых организмов благоприятны для других, с противоположными требованиями.
На основе конкурентных взаимодействий видов в ходе сукцессии происходит постепенное формирование более устойчивых комбинаций, соответствующих конкретным абиотическим условиям среды. Пример сукцессии, приводящей к смене одного сообщества другим, - зарастание небольшого озера с последующим появлением на его месте болота, а затем леса (рис. 20).
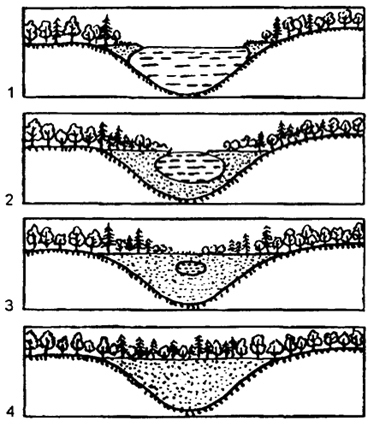
Рис. 20. Сукцессия при зарастаниинебольшого озера(по А.О. Рувинскому и др., 1993)
Вначале по краям озера образуется сплавна - плавающий ковер из осок, мхов и других растений. Постоянно озеро заполняется отмершими остатками растений - торфом. Образуется болото, постепенно зарастающее лесом. Последовательный ряд постепенно и закономерно сменяющих друг друга в сукцессии сообществ называетсясукцессионной серией.
Сукцессии в природе чрезвычайно разномасштабны. Их можно наблюдать в банках с культурами, представляющими собой планктонные сообщества - различные виды плавающих водорослей и их потребителей - коловраток, жгутиковых в лужах и прудах, на болотах, лугах, в лесах, на заброшенных пашнях, выветрившихся скалах. В организации экосистем иерархичность проявляется и в сукцессионных процессах - более крупные преобразования биоценозов складываются из более мелких. В стабильных экосистемах с отрегулированным круговоротом веществ также постоянно осуществляются локальные сукцессионные смены, поддерживающие сложную внутреннюю структуру сообществ.
Типы сукцессионных смен. Выделяют два главных типа сукцессионных смен: 1 - с участием автотрофного и гетеротрофного населения; 2 - с участием только гетеротрофов. Сукцессии второго типа совершаются лишь в таких условиях, где создается предварительный запас или постоянное поступление органических соединений, за счет которых и существует сообщество: в кучах или буртах навоза, в разлагающейся растительной массе, в загрязненных органическими веществами водоемах.
Процесс сукцессии. По Ф. Клементсу (1916), процесс сукцессии состоит из следующих этапов: 1. Возникновение незанятого жизнью участка. 2. Миграция на него различных организмов или их зачатков. 3. Приживание их на данном участке. 4. Конкуренции их между собой и вытеснение отдельных видов. 5. Преобразование живыми организмами местообитания, постепенной стабилизации условий и отношений. Сукцессии со сменой растительности могут быть первичными и вторичными.
Первичной сукцессиейназывается процесс развития и смены экосистем на незаселенных ранее участках, начинающихся с ихколонизации. Классический пример - постоянное обрастание голых скал с развитием в конечном итоге на них леса. Так, в первичных сукцессиях, протекающих на скалах Уральских гор, различают следующие этапы.
1. Поселение эндолитических и накипных лишайников, сплошь покрывающих каменистую поверхность. Накипные лишайники несут своеобразную микрофлору и содержат богатую фауну простейших, коловраток, нематод. Мелкие клещи - сапрофаги и первичнобескрылые насекомые обнаруживаются сначала только в трещинах. Активность всего населения прерывиста, отмечается главным образом после выпадения осадков в виде дождя или смачивания скал влагой туманов. Данные сообщества организмов называютпионерными.
2. Преобладание листоватых лишайников, которые постепенно образуют сплошной ковер. Под круговинками лишайников в результате выделяемых ими кислот и механического сокращения слоевищ при высыхании образуются выщербленности, идет отмирание слоевищ и накопление детрита. В большом количестве под лишайниками встречаются мелкие членистоногие: коллемболы, панцирные клещи, личинки комаров-толкунчиков, сеноеды и другие. Образуется микрогоризонт, состоящий из их экскрементов.
3. Поселение литофильных мхов Hedwidia u Pleuroziumschreberi. Под ними погребаются лишайники и подлишайниковые пленочные почвы. Ризоиды мхов здесь прикрепляются не к камню, а к мелкозему, который имеет мощность не менее 3 см. Колебания температуры и влажности под мхами в несколько раз меньше, чем под лишайниками. Усиливается деятельность микроорганизмов, увеличивается разнообразие групп животных.
4. Появление гипновых мхов и сосудистых растений. В разложении растительных остатков и формировании почвенного профиля постепенно уменьшается роль мелких членистоногих и растет участие более крупных беспозвоночных - сапрофагов: энхитреид, дождевых червей, личинок насекомых.
5. Заселение крупными растениями, способствующее дальнейшему накоплению и образованию почвы. Ее слой оказывается достаточным для развития кустарников и деревьев. Их опадающие листья и ветви не дают расти мхам и большинству других мелких видов, начавших сукцессию. Так, постепенно на изначально голых скалах идет процесс смены лишайников мхами, мхов травами и наконец лесом. Такие сукцессии в геоботанике называют экогенетическими, так как они ведут к преобразованию самого местообитания.
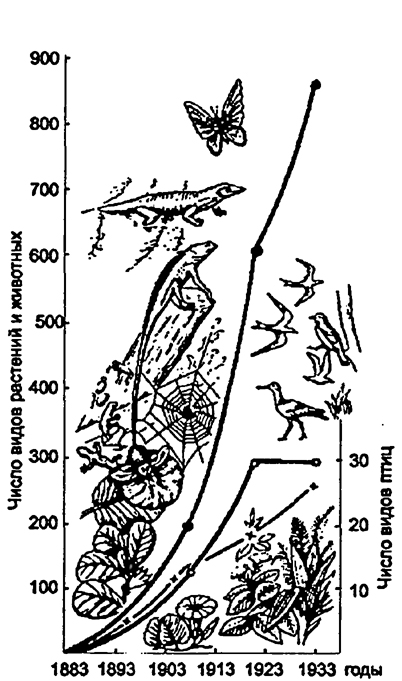
Рис. 21. Динамика видового разнообразия нао. Кракатау после полного уничтоженияаборигенной флоры и фаунывулканическим взрывом в 1893 г.(по Р. МакАртуру и Е.О. Вильсону, 1967)
Примечаниекрестики - число видов растений,светлые кружки - число видов гнездящихся птиц,зачерненные кружки - суммарное число видоврастений и животных.
Вторичная сукцессия- это восстановление экосистемы, когда-то уже существовавшей на данной территории. Она начинается в том случае, если уже в сложившемся биоценозе нарушены установившиеся взаимосвязи организмов в результате извержения вулкана, пожара, вырубки, вспашки и т.д. Смены, ведущие к восстановлению прежнего биоценоза, получили в геоботанике названиедемутационных.Примером может служить динамика видового разнообразия на острове Кракатау после полного уничтожения аборигенной флоры и фауны вулканическим взрывом в 1893 году (рис. 21).
Другой пример, вторичная сукцессия сибирского темно-хвойного леса (пихтово-кедровой тайги) после опустошительного лесного пожара (рис. 12.38). На более выжженных местах из спор, занесенных ветром, появляются мхи-пионеры: через 3-5лет после пожара наиболее обильны "пожарный мох" -Funariahygrometrica, Geratodonpurpureus, и др. Из высших растений весьма быстро заселяют гари Иван-чай (Chamaenerionangustifolium), который уже через 2-3 месяца обильно цветет на пожарище, а также вейник наземный (Calamagrostisepigeios) и другие виды.
Наблюдается дальнейшее происхождение фаз сукцессии: вейниковый луг сменяется кустарниками, затем березовым или осиновым лесом, смешанным сосново-лиственным лесом, сосновым лесом, сосново-кедровым лесом, и, наконец, через 250 лет происходит восстановление кедрово-пихтового леса.
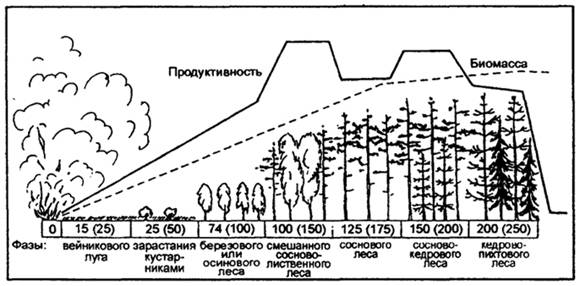
Рис. 22. Вторичная сукцессия сибирского темно-хвойного леса (пихтово-кедровой тайги) после опустошительного лесного пожара (по Н.Ф. Реймерсу, 1990)
Примечание:числа в прямоугольниках - колебания в длительности прохождения фаз вторичной сукцессии (в скобках указан срок их окончания). Биомасса и биологическая продуктивность показаны в произвольном масштабе (кривые отражают качественную и количественную стороны процесса)
Вторичные сукцессии совершаются, как правило, быстрее и легче, чем первичные, так как в нарушенном местообитании сохраняется почвенный профиль, семена, зачатки и часть прежнего населения и прежних связей. Демутация не является повторением какого-либо этапа первичных сукцессий.
Климаксовая экосистема.Сукцессия завершается стадией, когда все виды экосистемы, размножаясь, сохраняют относительно постоянную численность и дальнейшей смены ее состава не происходит. Такое равновесное состояние называютклимаксом,а экосистему -климаксовой.В разных абиотических условиях формируются неодинаковые климаксовые экосистемы. В жарком и влажном климате это будет дождевой тропический лес, в сухом и жарком - пустыня. Основные биомы земли - этоклимаксовые экосистемы соответствующих географических областей.
Изменения в экосистеме во время сукцессии. Продуктивность и биомасса.Как уже отмечалось, сукцессия является закономерным, направленным процессом, а изменения, которые происходят на той или иной ее стадии, свойственны любому сообществу и не зависят от его видового состава или географического местоположения. Основными называют четыре типа сукцессионных изменений.
1. В процессе сукцессии виды растений и животных непрерывно сменяются. 2. Сукцессионные изменения всегда сопровождаются повышением видового разнообразия организмов. 3. Биомасса органического вещества увеличивается по ходу сукцессии. 4. Снижение чистой продукции сообщества иповышение интенсивности дыхания - важнейшие явления сукцессии.
Следует также отметить, что смена фаз сукцессии идет в соответствии с определенными правилами. Каждая фаза готовит среду для возникновения последующей. Здесь действуетзакон последовательности прохождения фаз развития:фазы развития природной системы могут следовать лишь в эволюционно закрепленном (исторически, экологически обусловленном) порядке, обычно от относительно простого к сложному, как правило, без выпадения промежуточных этапов, но, возможно, с очень быстрым их прохождением или эволюционно закрепленным отсутствием. Когда экосистема приближается к состоянию климакса, в ней, как и во всех равновесных системах, происходит замедление всех процессов развития. Это положение находит отражение взаконе сукцессионного замедления:процессы, идущие в зрелых равновесных экосистемах, находящихся в устойчивом состоянии, как правило, проявляют тенденцию к снижению темпов. При этом восстановительный тип сукцессии меняется на вековой их ход, т.е. саморазвитие идет в пределах климакса или узлового развития. Эмпирический закон сукцессионного замедления является следствиемправила Г. Одума и Р. Пинкертона, или правила максимума энергии поддержания зрелой системы:сукцессия идет в направлении фундаментального сдвига потока энергии в сторону увеличения ее количества, направленного на поддержание системы. Правило Г. Одума и Р. Пинкертона, в свою очередь, базируется на правиле максимума энергии в биологических системах, сформулированном А. Лоткой. Вопрос этот в дальнейшем был хорошо разработан Р. Маргалефом, Ю. Одумом и известен как доказательствопринципа "нулевого максимума",или минимализации прироста взрелой экосистеме:экосистема в сукцессионном развитии стремится к образованию наибольшей биомассы при наименьшей биологической продуктивности.
Линдеман (1942) экспериментально доказал, что сукцессии сопровождаются повышением продуктивности вплоть до климаксового сообщества, в котором превращение энергии происходит наиболее эффективно. Данные исследований сукцессии дубовых и дубово-ясеневых лесов показывают, что на поздних стадиях их продуктивность действительно возрастает. Однако при переходе к климаксному сообществу обычно происходит снижение общей продуктивности. Таким образом, продуктивность в старых лесах ниже, чем в молодых, которые, в свою очередь, могут иметь меньшую продуктивность, чем предшествовавшие им более богатые видами ярусы травянистых растений. Сходное падение продуктивности наблюдается и в некоторых водных системах. Для этого есть несколько причин. Одна из них то, что накопление питательныхвеществ в растущей биомассе леса на корню может вести к уменьшению их круговорота. Снижение общей продуктивности могло быть просто результатом уменьшения жизненности особей по мере увеличения их среднего возраста в сообществе.
По мере прохождения сукцессии все большая доля доступных питательных веществ накапливается в биомассе сообщества, и соответственно уменьшается их содержание в абиотическом компоненте экосистемы (в почве или воде).
Возрастает также количество образующегося детрита. Главными первичными консументами становятся не травоядные, а детритоядные организмы. Соответствующие изменения происходят и в трофических сетях. Детрит становится основным источником питательных веществ.
В ходе сукцессии увеличивается замкнутость биогеохимических круговоротов веществ. Примерно за 10 лет с момента начала восстановления растительного покрова разомкнутость круговоротов уменьшается со 100 до 10%, а далее она еще больше снижается, достигая минимума в климаксовой фазе.Правило увеличения замкнутости биогеохимического круговорота веществ в ходе сукцессиинарушается антропогенной трансформацией растительности и вообще естественных экосистем. Несомненно, это ведет к длинному ряду аномалий в биосфере и ее подразделениях.
Снижение разнообразия видов в климаксе не означает малой его экологической значимости. Разнообразие видов формирует сукцессию, ее направление, обеспечивает заполненность реального пространства жизнью. Недостаточное количество видов, составляющих комплекс, не могло бы сформировать сукцессионный ряд, и постепенно, с разрушением климаксовых экосистем произошло бы полное опустынивание планеты. Значение разнообразия функционально как в статике, так и в динамике. Следует отметить, что там, где разнообразие видов недостаточно для формирования биосферы, служащей основой нормального естественного хода сукцессионного процесса, а сама среда резко нарушена, сукцессия не достигает фазы климакса, а заканчивается узловым сообществом -параклимаксом,длительно или кратковременно производным сообществом. Чем глубже нарушенность среды того или иного пространства, тем на более ранних фазах оканчивается сукцессия.
При потере одного или группы видов в результате их уничтожения (антропогенное исчезновение местообитаний, реже вымирание) достижение климакса не является полным восстановлением природной обстановки. Фактически это новая экосистема, потому что в ней возникли новые связи, утеряны многие старые, сложилась иная "притертость" видов. В старое состояние экосистема вернуться не может, так как утерянный вид восстановить невозможно.
При изменении любого абиотического или биотического фактора, например, при устойчивом похолодании, интродукции нового вида, вид, который плохо приспособлен к новым условиям, ожидает один из трех путей (рис. 23).

Рис. 23. Эволюционная сукцессия (по Б. Небелу, 1993)
1. Миграция.Часть популяции может мигрировать, найти местообитания с подходящими условиями и продолжить там свое существование.
2. Адаптация.В генофонде могут присутствовать аллели, которые позволят отдельным особям выжить в новых условиях и оставить потомство. Через несколько поколений под действием естественного отбора возникает популяция, хорошо приспособленная к изменившимся условиям существования.
3. Вымирание.Если ни одна особь популяции не может мигрировать, опасаясь воздействия неблагоприятных факторов, а те уходят за пределы устойчивости всех индивидов, то популяция вымрет, а ее генофонд исчезает. Если одни виды вымирают, а выжившие особи других размножаются, адаптируются и изменяются под действием естественного отбора, можно говорить обэволюционной сукцессии.
Закон эволюционно-экологической необратимостигласит: экосистема, потерявшая часть своих элементов или сменившаяся другой в результате дисбаланса экологических компонентов, не может вернуться к первоначальному своему состоянию в ходе сукцессии, если в ходе изменений произошли эволюционные(микроэволюционные) перемены в экологических элементах (сохранившихся или временно утерянных). В том случае, когда какие-то виды утеряны в промежуточных фазах сукцессии, данная потеря может быть функционально скомпенсирована, но не полностью. При снижении разнообразия за критический уровень, ход сукцессии искажается, и фактически климакс, идентичный прошлому, достигнут не может быть.
Для оценки характера восстановленных экосистем закон эволюционно-экологической необратимостиимеет важное значение. При потере элементов это, по сути дела, совершенно экологически новые природные образования с вновь образовавшимися закономерностями и связями. Так, перенос в прошлом выбывшего из состава экосистемы вида в ходе его реакклиматизациине является механическим его возвращением. Это фактически внедрение нового вида в обновленную экосистему. Закон эволюционно-экологической необратимости подчеркивает направленность эволюции не только на уровне биосистем, но и на всех других иерархических уровнях сложения биоты.
Дата добавления: 2018-04-15; просмотров: 624; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!