Электрические процессы в синапсах
Законы распространения возбуждения
4.1. Деполяризация безмиелиновых и миелиновых волокон
4.2. Проведение возбуждения
4.1.Деполяризация безмиелиновых и миелиновых волокон
В безмиелиновых и миелиновых волокнах (рис. 9) возбуждение передается по-разному, это обусловлено строением самого волокна. Прежде всего, существуют большие различия по скорости проведения нервного импульса. Тонкие безмиелиновые волокна у позвоночных и человека чаще всего является афферентами болевой чувствительности или волокнами симпатического отдела вегетативной нервной системы. Скорость проведения нервного импульса здесь составляет 1-7м/c. Толстые миелиновые волокна вестибулоспинальной нисходящей системы спинного мозга проводят нервные импульсы со скоростью 120м/c, а ретикулоспинальные волокна медиальной части переднего канатика спинного мозга имеют самую высокую скорость проведения возбуждения до 130 м/c. Они преимущественно иннервируют мотонейроны сгибателей верхних и нижних конечностей.
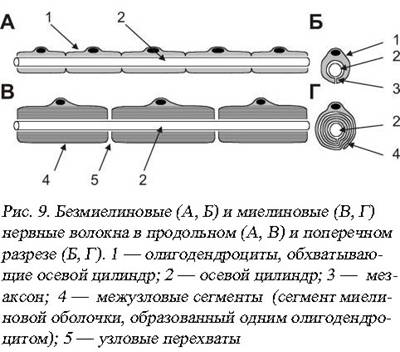 |
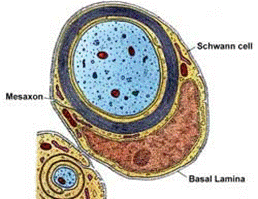
Как же влияет миелинизация волокна на механику проведения нервного импульса? Благодаря слабому развития или полному отсутствию мезаксона (сдвоенная мембрана, формирующая щелевидный канал при погружении нервного волокна в нейролеммоцит Шванна) на протяжении всей своей длины аксон контактирует с внеклеточным матриксом, то есть электрогенен по всей своей длине. Потенциал действия в таком волокне будет возникать непосредственно в точке действия стимула. Безмиелиновые волокна имеют одинаковую плотность Na+-каналов на всем протяжении. Волна деполяризации, а вслед за ней и волна реполяризации будет пробегать по всему волокну в виде круговых электрических возмущений – круговых токов.
|
|
|
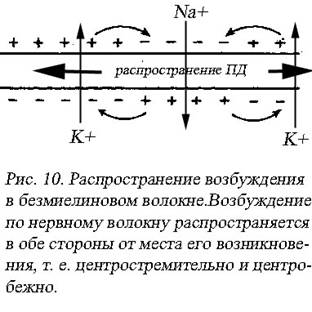
Идея передачи нервного импульса в виде круговых токов между возбужденным и невозбужденным участком волокна принадлежит Л. Герману (1885). В точке приложения стимула на поляризованной клеточной мембране возникает движение ионов Nа+ внутрь клетки. Из физики известно каждому, что движение заряженных частиц есть не что иное, как электрический ток. С внутренней стороны сопряжено с Nа+-током возникает ток от положительного полюса к отрицательному полюсу, иными словами ток направлен от возбужденного участка к невозбужденному. Через эти невозбужденные участки под происходит выход катионов К+, приводящий к развитию процессов реполяризации. На наружной поверхности нервного волокна ток идет от невозбужденного участка к возбужденному. Этот ток не изменяет состояние возбужденного участка, так как он находится в состоянии рефрактерности. Вона деполяризации распространяется независимо в обе стороны по изолированному волокну, когда стимул уже перестал действовать по причине того, что электрические круговые возмущения действуют на внешние невозбужденные участки как стимулы (рис. 10).
|
|
|
В миелиновом волокне мембрана по большей части изолирована Шванновскими нейролеммоцитами. Если стимул действует на изолированный участок – он не может возбудиться: глиальные клетки не генерируют ПД, мембрана аксона покрытого рулетообразной миелиновой структурой нейролеммоцита содержит недостаточное для деполяризации число Nа+-каналов. Зато в перехватах Ранвье плотность Nа+-каналов оказывается необычайно высокой и достигает 12000 на 1 мкм2, что в 100 раз выше, чем в мембранах безмиелиновых волокон. Под действием стимула деполяризуется ближайший к месту нанесения раздражения перехват Ранвье. Из-за высокой плотности Nа+-каналов создается довольно мощное электромагнитное возмущение, которое индуцирует деполяризацию 2-3 и даже 5 последующих перехватов Ранвье одновременно. Скорость нервного импульса будет определяться скоростью распространения электромагнитного поля в плотной среде, она ниже, чем скорость распространения в вакууме (С=299792458м/с – скорость света), но многократно выше скорости распространения электрического тока по поверхности проводника. Нервный 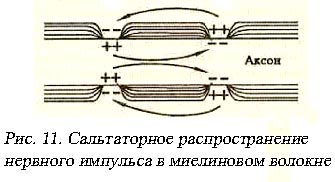
импульс как бы перескакивает от перехвата к перехвату (рис. 11), ретранслируясь, усиливаясь за счет вовлечения все большего числа перехватов Ранвье в процессы деполяризации. Миелин имеет малую емкость, чем и объясняется большая скорость проведения импульса. Теория скачкообразного или сальтаторного проведения нервного импульса в миелиновом волокне впервые была предложена Лилли, прямые доказательства были получены в работах Кубо, Оно и Юге. Количественная теория этого эффекта была развита Тасаки в 1941 году. Многочисленные и разнообразные эксперименты с рассеченными и нативными миелиновыми волокнами показали, что перерезка волокна или локальное блокирование перехватов Ранвье (путем введения новокаина, кокаина, уретана, или под воздействием ультрафиолетового облучения), не препятствует распространению нервного импульса. «Скачок» нервного импульса через участок между перехватами возможен потому, что амплитуда самого ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата.
|
|
|
Участки миелинового волокна, покрытые нейролеммоцитами Шванна, занимающие большую площадь, чем суммарная площадь всех принадлежащих волокну перехватов Ранвье, не включены в процесс деполяризации. Следовательно, здесь нет затраты АТФ на работу натрий-калиевой аденозинтрифосфатазы, она будет активироваться только в перехватах Ранвье. Поэтому затраты энергии на восстановление возбудимости в миелиновом волокне несоизмеримо малы по сравнению с таковыми показателями безмиелинового волокна. Сравнительные данные по характеристикам миелиновых и безмиелиновых волокон приведены в таблице 4.
|
|
|
Таблица 4
Физиология нервных волокон
| Тип волокна | безмиелиновые | миелиновые |
| скорость проведения нервного импульса | кожные афференты боли – 3-15м/с симпатические постганглионарные волокна – 0,5-2 м/с | Первичные афференты мышечных веретен, двигательные волокна скелетных мышц – 70-120 м/с |
| Топографические отношения с нейролеммоцитами | один слой шванновских клеток, мезаксон формирует щелевидные пространства
| многослойная миелиновая оболочка, перехваты Ранвье
|
| Электрогенность (способность генерировать нервные импульсы) | на всем протяжении волокна, ПД возникает в точке действия стимула | только в области перехватов Ранвье, ПД возникает в ближайшем к месту действия стимула перехвате |
| Способ проведения возбуждения | возбуждение имеет вид распространяющейся волны круговых электромагнитных возмущений по поверхности волокна | возбуждение распространяется скачкообразно от перехвата к перехвату, с ретрансляцией |
| Затраты энергии на восстановление возбудимости | высокие – Na-K-АТфаза активируется по всей поверхности волокна | низкие – Na-K-АТфаза активируется только в перехватах Ранвье |
4.2.Проведение возбуждения
Основной закон, которому подчиняется нервный импульс – «все или ничего» был сформулирован в 1871 году американским нейрофизиологом Боудичем Генри Пикерингом. Свои эксперименты ученый проводил на сердечной мышце лягушки. Согласно этому закону, подпороговый раздражитель как бы долго он не действовал, не вызывает возбуждения (ничего), при достижении пороговых значений раздражитель вызывает волну деполяризации с максимальной амплитудой (все). Увеличение силы раздражителя не приводит к усилению возбуждения.
Более поздние исследования с использованием микроэлектродной техники показали, что существует множество исключений. Ученик М.И. Сеченова русский физиолог Николай Евгеньевич Введенский посвятил свою жизнь исследованиям по выявлению ответной реакции живых тканей на различные раздражители. Именно он ввел фактор времени как значимую компоненту, влияющую на ответ со стороны возбудимой ткани. В своей книге: «Возбуждение, торможение и наркоз» Введенский показал, что живая ткань реагирует на внешние раздражители неодинаково. Если нерв подвергнуть альтерации (воздействовать повреждающим агентом), то это приведет, ,прежде всего, к резкому снижению лабильности. То есть при воздействии стимула на такой поврежденный участок частота генерации ПД будет значительно ниже, чем в неповрежденных зонах. Это явление получило название парабиоза (от греч. para—возле и bios- жизнь). Впоследствии оказалось, что именно явление парабиоза является механизмом действия многих анестезирующих веществ. Ответная реакция в нервном волокне при парабиозе развивается в три стадии:
· Провизорная стадия – исчезновение различий в действии слабых и сильных, или частых и редких ритмических раздражений. Эта фаза чаще известна как уравнительная. Термин введен учеником Введенского К.М. Быковым. Механизм развития этих изменений состоит в том, что в поврежденном участке одновременно при снижении лабильности происходит удлинение периода рефрактерности. При действии слабых или редких стимулов участок успевает восстановить свою возбудимость. Слабые ритмичные раздражения 25Гц будут вызывать такие же слабые ПД с такой же частотой. Частые или сильные раздражения, например, в 100 Гц будут проводиться через поврежденный участок с такой же частотой, как и слабые (редкие) – 25 Гц, Поврежденный участок пропускает один импульс из четырех по причине того, что воздействие других стимулов будет приходиться на рефрактерное состояние волокна.
· Парадоксальная стадия – на сильное или частое раздражение возникает слабая реакция ткани, в ответ на слабые и редкие раздражения – более сильный ответ, чем на сильное раздражение. Это связано с тем, что лабильность ткани продолжает снижаться. Сильные и частые стимулы вызывают еще большее удлинения рефрактерного периода.
· Тормозная стадия – ткань утрачивает способность отвечать на раздражение. Никакие раздражители не в состоянии вызвать возбуждение поврежденного участка, так как мембрана нервного волокна деполяризована и не переходит в стадию реполяризации по причине инактивации натрий-калиевого насоса, т. е. отсутствует возможность восстановления ее исходной возбудимости. Многие анестетики также понижают лабильность ткани, инактивируя Na+-каналы.
Законы проведения возбуждения в отдельном волокне и в нерве будут также сильно различаться. Зависимость ответной реакции от параметров раздражителя описывается тремя законами: законом силы, законом длительности и законом градиента раздражителя.
Закон силы раздражителя: ответная реакция ткани пропорциональна силе наносимых раздражений до определенного предела. Этому закону подчиняются сложные структуры – нервы и мышцы, состоящие из большого числа отдельных волокон. Механизм – вовлечение в процесс возбуждения все большего числа составных компонентов. Поскольку число нервных волокон в нерве не безгранично, при определенной силе стимула, когда все они будут задействованы, дальнейшее увеличение силы стимула не вызовет усиления ответной реакции.
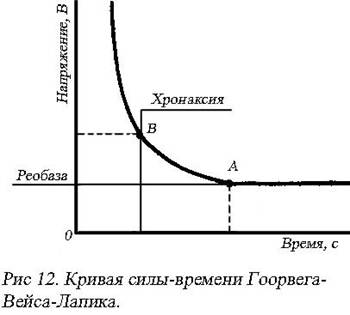 |
Закон длительности действия раздражителя: ответная реакция ткани зависит от времени действия раздражителя, но до определенного предела. Графически эти два закона выражаются гиперболой известной как кривая силы-времени Гоорвега-Вейса-Лапика (рис. 12). Голландский физик Л. Горвег (1892) и французский физиолог Ж. Вейс (1901) экспериментально доказали, что величина стимула, вызывающего возбуждающий эффект в тканях, обратнопропорциональна длительности его действия. Подпороговые раздражители, как бы долго они не действовали, не вызывают ответной реакции в нерве. Минимальная величина стимула (в эксперименте – величина электрического тока), способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, через которое возникает возбуждение при действии стимула в одну реобазу, называется полезным временем (точка А, рис.12). Определение этого времени экспериментально бывает сильно затруднено, так как соседние точки, мало от нее отличаются. Вправо и влево от точки А кривая изменяется незначительно. Поэтому французский физиолог Луи Лапик (1909) предложил вместо полезного времени определять минимальное время действия тока в две реобазы, которое обозначается как хронаксия. Точка В – хронаксия, она легко и точно определяется, поскольку изменения кривой вправо и влево от нее значительны.
Как и в случае с законом силы раздражителя ответная реакция ткани не безгранична. Когда в процесс возбуждения вовлечены все составляющие волокна нерва, дальнейшее увеличение времени воздействия стимула не приведет к усилению ответной реакции.
В клинических исследованиях измерение возбудимости тканей методом хронаксиметрии довольно широко распространено. Уменьшение хронаксии указывает на улучшение функциональных свойств, увеличение хронаксии — на ухудшение. Применяется для исследования возбудимости мышечных тканей, органов чувств (зрительная, вестибулярная, болевая), для выявления патологии нервных структур. Однако у этого метода есть недостаток: одиночная стимуляция – редкое явление в организме, современные методы, как то: электромиография, электроэнцефалография, томография, являются более физиологичными и репрезентативными.
Закон градиента раздражителя: ответная реакция ткани зависит от градиента раздражителя до определенных пределов. Под градиентом силы раздражителя понимают крутизну нарастания силы раздражителя во времени. Этот закон известен и под другим названием – закон раздражения Дюбуа-Реймона (аккомодации): раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени. Если раздражитель по силе нарастает очень медленно, то на него может вообще не возникнуть ответной реакции. Механизм этого феномена достаточно прост: в обозначенных условиях развивается инактивация натриевых каналов, вследствие чего происходит постоянное повышение порога раздражения.
Справедливости отметим, что законов проведения возбуждения много больше, чем перечислено в данном пособии. Среди них законы полярного действия тока (при замыкании тока возбуждение возникает под катодом, а при размыкании - под анодом), физиологического электротона (действие постоянного тока на ткань сопровождается изменением ее возбудимости), двустороннего проведения возбуждения по нервному волокну, анатомической и физиологической целостности нервного волокна, изолированного проведения возбуждения по нервному волокну. Со всеми этими законами студент-гуманитарий при желании может ознакомиться в соответствующей литературе по физиологии. В настоящем пособии приведены лишь те законы, без которых просто невозможно было бы раскрыть процессы проведения нервного импульса, показать его материальную, физико-химическую природу.
Электрические процессы в синапсах
5.1. Возбуждающий постсинаптический потенциал (ВПСП)
5.2. Торможение в нервной системе
Дата добавления: 2018-04-05; просмотров: 125; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!