Основные этапы эмбрионального развития
Период эмбрионального развития наиболее сложен у высших животных и состоит из нескольких этапов.
Период начинается с этапа дробления зиготы(рис. 1), т. е. серии последовательных митотических делений оплодотворенной яйцеклетки. Образующиеся в результате деления две клетки (и все последующие их поколения) на этом этапе называются бластомерами. Одно деление следует за другим, причем не происходит роста образующихся бластомеров и с каждым делением клетки становятся все более мелкими. Такая особенность клеточных делений и определила появление образного термина «дробление зиготы».
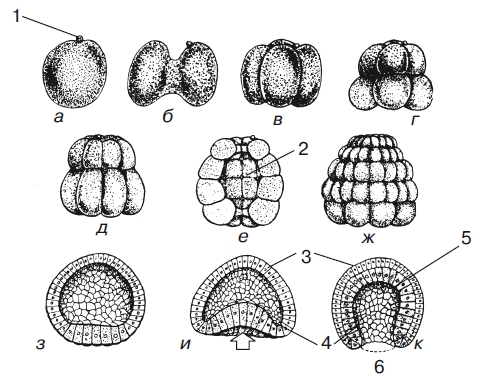
Рис. 1.Дробление и гаструляция яйца ланцетника (вид сбоку)
На рисунке обозначены: а— зрелое яйцо с полярным тельцем; б— 2-клеточная стадия; в— 4-клеточная стадия; г— 8-клеточная стадия; д— 16-клеточная стадия; е— 32-клеточная стадия (в разрезе, чтобы показать бластоцель); ж — бластула; з — разрез бластулы; и — ранняя гаструла (на вегетативном полюсе — стрелка — начинается инвагинация); к — поздняя гаструла (инвагинация закончилась и образовался бластопор; 1 — полярное тельце; 2 — бластоцель; 3 — эктодерма; 4 — энтодерма; 5 — полость первичной кишки; 6 — бластопор).
В результате дробления (когда количество бластомеров достигнет значительного числа) образуется бластула (см. рис. 1, ж, з). Часто она представляет собой полый шар (например, у ланцетника), стенка которого образована одним слоем клеток — бластодермой. Полость бластулы — бластоцель, или первичная полость, заполнена жидкостью.
На следующем этапе осуществляется процесс гаструляции — формирование гаструлы. У многих животных она образуется путем впячивания бластодермы внутрь на одном из полюсов бластулы при интенсивном размножении клеток в этой зоне. В результате и возникает гаструла (см. рис. 1, и, к).
Наружный слой клеток получил название эктодермы, а внутренний — энтодермы. Внутренняя полость, ограниченная энтодермой, полость первичной кишки сообщается с внешней средой первичным ртом, или бластопором. Существуют и другие типы гаструляции, но у всех животных (кроме губок и кишечнополостных) этот процесс завершается образованием еще одного клеточного пласта — мезодермы. Она закладывается между энто- и эктодермой.
По завершении этапа гаструляции появляются три клеточных пласта (экто-, эндо- и мезодерма), или три зародышевых листка.
Далее начинаются процессы гистогенеза (образования тканей) и органогенеза (образования органов) у зародыша (эмбриона). В результате дифференцировки клеток зародышевых листков формируются различные ткани и органы развивающегося организма. Из эктодермы образуются покровы и нервная система. За счет энтодермы формируются кишечная трубка, печень, поджелудочная железа, легкие. Мезодерма продуцирует все остальные системы: опорно-двигательную, кровеносную, выделительную, половую. Обнаружение гомологии (сходства) трех зародышевых листков едва ли не у всех животных послужило важным аргументом в пользу точки зрения о единстве их происхождения. Изложенные выше закономерности были установлены в конце XIX в. И. И. Мечниковым и А. О. Ковалевским и легли в основу сформулированного ими «учения о зародышевых листках».
На протяжении эмбрионального периода наблюдается ускорение темпов роста и дифференцировки у развивающегося эмбриона. Только в процессе дробления зиготы роста не происходит и бластула (по своей массе) может даже существенно уступать зиготе, но начиная с процесса гаструляции масса зародыша стремительно увеличивается.
Образование разнотипных клеток начинается еще на этапе дробления и лежит в основе первичной тканевой дифференцировки — возникновения трех зародышевых листков. Дальнейшее развитие зародыша сопровождается все более усиливающимся процессом дифференцировки и морфогенеза. К концу эмбрионального периода у зародыша имеются уже все основные органы и системы, обеспечивающие жизнеспособность во внешней среде.
Завершается эмбриональный период рождением новой особи, способной к самостоятельному существованию.
Половые клетки у рыб формируются в гонадах — половых железах. В соответствии с современными представлениями у рыб зачаток первичных половых клеток — гоноцитов обособляется в конце гаструляции. Их источником служит первичная энтомезодерма, а временным прибежищем перед началом миграции в гонаду — перибласт. Возможно, что в гонадах взрослых рыб присутствуют первичные половые клетки.
Процесс развития женских половых клеток называется оогенезом. Гоноциты вселяются в зачаток женской гонады, и все дальнейшее развитие женских половых клеток происходит в ней. Структура оогенеза в принципе одинакова у всех животных. Попав в яичник, гоноциты становятся оогониями.
Оогоний — это незрелая половая клетка, способная к митозу. Оогонии осуществляют первый период оогенеза — период размножения. В этот период оогонии делятся митотическим путем. Количество делений видоспецифично. У рыб и амфибий периодичность митотических делений оогониев связана с сезонным размножением и повторяется в течение всей жизни.
Следующий период оогенеза — период роста. Половые клетки в этом периоде называются ооцитами первого порядка. Они теряют способность к митотическому делению и вступают в профазу I мейоза. В этот период осуществляется рост половых клеток.
Выделяют стадию малого и стадию большого роста. Главным процессом периода большого роста является процесс образования желтка (вителлогенез, вителлус — желток).
В период малого роста (превителлогенез, цитоплазматический рост) объемы ядра и цитоплазмы увеличиваются пропорционально и незначительно. При этом ядерно-цитоплазматические отношения не нарушаются. В период большого роста (вителлогенез) в высшей степени интенсифицируются синтез и поступление включений в цитоплазму, приводящие к накоплению желтка. Ядерно-цитоплазматическое отношение уменьшается. Часто яйцеклетка в этот период сильно увеличивается и ее размер возрастает в десятки (человек), сотни тысяч раз (лягушки, дрозофилы) и более (акуловые рыбы и птицы).
Различают следующие типы питания яйцевых клеток:
Фагоцитарный тип — встречается у половых клеток животных, не имеющих половых желез (губки, кишечнополостные). При фагоцитарном способе вителлогенеза ооциты, перемещаясь по межклеточному пространству, способны фагоцитировать соматические клетки организма.
Солитарный тип — встречается у колониальных гидроидных полипов, иглокожих, червей, бескрылых насекомых, ланцетника. При солитарном способе питания ооцит получает ингредиенты из целомической жидкости и из половой железы. Желточные белки синтезируются в эндоплазматическом ретикулуме, а формирование желточных гранул происходит в аппарате Гольджи.
Алиментарный тип — осуществляется с помощью вспомогательных клеток; подразделяется на нутриментарный и фолликулярный.
Нутриментарный способ питания встречается у червей и членистоногих. У них ооцит в яичнике окружен трофоцитами (клетка-микормилками), с которыми он связан цитоплазматическими мостиками. Ооцитом становится та клетка, которая контактирует с большим числом сестринских клеток (клеток-кормилок). Фолликулярный способ питания встречается у большинства животных. Вспомогательными клетками при этом способе питания служат гоматические клетки в составе яичника. В оогенез вступает фолликул, т. е. ооцит вместе со вспомогательными фолликулярными клетками. Основная масса желтка образуется за счет поступления веществ извне и овоциты с экзогенным синтезом желтка растут с большой скоростью. В поверхностной зоне ооцита появляется множество пиноцитарных пузырьков, содержащих вителлогенин — предшественник желточных белков, поступаемых из крови.
Вителлогенины у разных животных синтезируются в разных соматических тканях и в процессе эволюции постепенно концентрируются и строго определенном органе. У позвоночных вителлогенин вырабатывается печенью самок. Вителлогенин синтезируется клетками печени и находится под гормональным контролем.
Для начала процесса развития зародыша с момента оплодотворения яйца в самом яйце производится некоторая подготовка к этому событию. Ядро яйца должно быть приведено в соответствующее состояние для того, чтобы соединиться с ядром сперматозоида; при этом часть хромосомного материала удаляется из яйца, переходя в маленькие полярные тельца (процесс этот обычно не завершается к моменту оплодотворения, а временно приостанавливается). Далее цитоплазматическое содержимое яйцеклетки приобретает перед оплодотворением довольно высокий уровень организованности; к этому времени характер ее будущей симметрии, по-видимому, уже в значительной степени определен, хотя дальнейшие события могут ее модифицировать. Количество содержащегося в яйце желтка сильно варьируется; оно служит основным фактором, определяющим размеры яйца и тип дробления. У некоторых животных, в частности у ланцетника и у человека, яйца содержат мало желтка. Такие яйца можно назвать олиголецитальными. Яйца другого типа несколько крупнее и содержат умеренное количество желтка; их называют мезолецитальными. К типичным мезолецитальным яйцам относятся яйца лягушки; к ним принадлежат также яйца хвостатых амфибий, двоякодышащих, низших лучеперых рыб и миног.
Мезолецитальные яйца так широко распространены среди примитивных водных форм, что они, по-видимому, были характерны для предковых позвоночных. У акул и скатов, с одной стороны, и у рептилий и птиц — с другой, яйца крупные; их называют полилецитальными, потому что большую часть клетки занимает желток, а цитоплазма, которой относительно мало, сконцентрирована на одном полюсе.
Яйца также классифицируются на основе распределения в них желтка. В некоторых яйцах, главным образом олиголецитальных, желток распределен по клетке довольно равномерно; такие яйца называют изолецитальными. В мезо- и полилецитальных яйцах желток в большинстве случаев сосредоточен в одной половине яйца; у яиц, плавающих в воде, — в нижней половине. Такие яйца называют телолецитальными. У современных костистых рыб яйца также очень богаты желтком, но размеры их варьируют.
Концентрация желтка в одном полушарии ясно свидетельствует о наличии в яйце определенной организации, или полярности: на верхнем его конце находится анимальный полюс, а на нижнем — вегетативный; верхняя половина яйца заполнена относительно прозрачной цитоплазмой, а нижняя переполнена желтком.
Яйца рыб, как и яйца позвоночных, чрезвычайно разнообразны по величине, представляют собой, как правило, сферические клетки, содержащие помимо ядра и некоторого количества прозрачной цитоплазмы желток, служащий пищей развивающемуся зародышу. Икринки у рыб обычно шаровидные, хотя есть и другие формы. Строение икринок является характерным признаком не только для рода, семейства, но и более крупных категорий.
Икринки рыб различаются не только формой, но и размером, цветом, наличием или отсутствием жировых капель, строением оболочки. Величина икринок, как и другие морфологические признаки, является стабильным признаком вида. Крупные рыбы откладывают икру большего диаметра, чем мелкие, но амплитуда колебаний размера икринок остается постоянной для вида даже в разных водоемах, хотя средние значения их могут смещаться в ту или иную сторону.
Размеры икринок зависят от содержания в них питательного вещества — желтка и значительно колеблются (в мм): тюлька — 0,8-1,05, сазан — 1,4-1,5, белый амур — 2,0-2,5, русский осетр — 3,0-3,5, семга — 5,0-6,0, кета — 6,5-9,1, полярная акула — 80 (без капсулы), китовая акула — 670 (в длину с капсулой).
Среди многочисленных костистых рыб наиболее мелкие икринки характерны для камбалы-лиманды, самые крупные — для лососевых, особенно для кеты. Большой объем желтка в икринках лососевых, в отличие от других рыб, обеспечивает более длительный период развития, появление более крупных личинок, способных на первом этапе активного питания потреблять более крупные кормовые организмы. Самые крупные яйца наблюдаются у хрящевых рыб. Развитие эмбрионов у некоторых из них (катран) длится почти 2 года.
Окраска икринок специфична для каждого вида. У ряпушки они желтые, у лососей оранжевые, у щуки темно-серые, у сазана зеленоватые, у терпугов изумрудно-зеленые, голубые, розовые и фиолетовые. Желтоватые и красноватые тона обусловлены наличием дыхательных пигментов — каротиноидов. Икра, развивающаяся в менее благоприятных кислородных условиях, обычно окрашена интенсивнее. Из лососевых у нерки наиболее яркая малиново-красная икра, развивающаяся в воде, относительно бедной кислородом. Пелагические икринки, развивающиеся при достаточном содержании кислорода, пигментированы слабо.
Икринки многих рыб содержат одну или несколько жировых капель, которые наряду с другими способами, например обводнением, обеспечивают плавучесть икринок. Икринки снаружи покрыты оболочками, которые могут быть первичными, вторичными и третичными.
Первичная — желточная, или лучистая, оболочка, образованная самим яйцом, пронизана многочисленными порами, по которым в яйцо поступают питательные вещества во время его развития в яичнике. Эта оболочка достаточно прочная, причем у осетровых двухслойная.
Над первичной оболочкой у большинства рыб развивается вторичная оболочка, студенистая, липкая, с разнообразными выростами для прикрепления яиц к субстрату.
На анимальном полюсе обеих оболочек расположен особый канал— микропиле, по которому сперматозоид проникает в яйцо. У костистых имеется один канал, у осетровых их может быть несколько. Существуют также третичные оболочки — белковая и роговая. Роговая оболочка развивается у хрящевых рыб и миксин, белковая — только у хрящевых. Роговая оболочка хрящевых рыб значительно больше самого яйца, не соответствует ему по форме, сплюснута и слегка сжимает яйцо. Часто от нее отходят роговые нити, с помощью которых яйцо прикрепляется к водным растениям. У яйцеживородящих и живородящих видов роговая оболочка очень тонкая, исчезающая вскоре после начала развития.
Партеногенез. Развитие яйцеклетки возможно и без участия сперматозоида и в таком случае оно называется партеногенезом (от греч. «партеноз» — девственница, «генез» — возникновение).
Известны случаи, когда организмы нормально развиваются из отложенных неоплодотворенных яиц.
Когда говорят о партеногенезе, то имеют в виду развитие на основе женского пронуклеуса. Однако в некоторых случаях возможно развитие на базе мужского пронуклеуса, и тогда говорят об андрогенезе, противопоставляя ему гиногенез. Гиногенез — это форма однополого развития, при которой сперматозоид активирует яйцеклетку, побуждая ее к развитию, но его ядро (мужской пронуклеус) не сливается с женским и в разпитии не участвует. Естественный гиногенез известен у одного вида карася, икра которого осеменяется спермой другого вида, активирует икру, но ядро сперматозоида не участвует в образовании зиготы. Андрогенез — явление гораздо более редкое, и когда он происходит (естественный или искусственный), развитие идет без женского пронуклеуса на базе мужского ядра и мужского пронуклеуса.
Половые клетки самцов — сперматозоиды в противоположность ооцитам — мелкие, многочисленные и подвижные. Каждая группа сперматозоидов — производное одной инициальной клетки и развивается в виде клона синцитиально связанных клеток, а по численности и некоторым особенностям строения дает группу отдельных подвижных клеток. Само развитие сперматозоидов у разных животных схоже. Сперматогенез всегда тесно связан со вспомогательными обслуживающими клетками соматического происхождения. Взаиморасположение половых и соматических обслуживающих клеток в достаточной мере специфично характеризует сперматогенез и представляет наибольший интерес. Развитие сперматозоида правильнее рассматривать не как «биографию» отдельной половой клетки самца, а как историю жизни клона.
Мужские половые клетки никогда не развиваются в одиночку, а растут в виде клонов синцитиально связанных клеток, где все клетки оказывают друг на друга влияние.
У большинства животных в процессе сперматогенеза принимают участие вспомогательные соматические клетки фолликулярного эпителия («опорные», «питающие»).
Половые клетки и связанные с ними вспомогательные клетки на ранней стадии развития отделяются от клеток сомы слоем пограничных клеток, выполняющих барьерную функцию. Внутри самой гонады происходит дальнейшее структурное обособление в виде цист или канальцев, где вспомогательными фолликулярными клетками создается специфическая среда сперматогенеза.
Первичные половые клетки, в том числе мужские, у многих животных могут быть идентифицированы задолго до образования гонады и часто вообще на очень ранних стадиях развития. Половые клетки появляются на ранних этапах развития у эмбрионов в генитальных складках, которые тянутся вдоль полости тела. У молоди лососей (горбуша, кета, нерка, сима, кижуч и атлантический лосось) первичные половые клетки обнаруживаются на стадии формирования первично-почечных протоков. У зародыша атлантического лосося первичные половые клетки были выявлены в возрасте 26 cyт. У мальков рыб уже можно найти половые железы в виде волосовидных тяжей.
Отличительной особенностью процесса развития сперматозоидов – сперматогенеза – является многократное уменьшение клеток. Каждый исходный сперматогоний делится несколько раз, в результате чего возникает скопление сперматогониев под одной оболочкой, называемое цистой (стадия размножения). Образовавшийся при последнем делении сперматогоний несколько увеличивается, в его ядре происходят мейотические преобразования и сперматогоний превращается в сперматоцит I порядка (стадия роста). Затем наступают два последовательных деления (стадия созревания): сперматоцит I порядка делится на два сперматоцита II порядка, вследствие деления которых образуются две сперматиды. В следующей – завершающей – стадии формирования сперматиды превращаются в сперматозоиды. Таким образом, из каждого сперматоцита образуются четыре сперматиды с половинным (гаплоидным)набором хромосом. Оболочка цисты лопается, и сперматозоиды наполняют семенной каналец. Через семяпроводы созревшие сперматозоиды выходят из семенника, а затем по протоку – наружу.
Сперматозоид вносит в яйцо ядерный материал, играющий важную роль в наследственности и в преобразованиях на более поздних стадиях развития, но не оказывающий существенного влияния на ранние стадии. Яйцо содержит все необходимое для полного развития взрослой особи. Зрелое яйцо готово к развитию; оно лишь ожидает соответствующего стимула, чтобы приступить к дроблению на клеточные единицы, что составляет первый шаг, необходимый для развития тканей и органов сложного взрослого организма. Во многих случаях этот процесс может быть запущен физическими или химическими стимулами. Однако в нормальных условиях начало процесса развития стимулируется проникновением в яйцо сперматозоида.
Вопрос 9
Особенности размножения рыб
Можно выделить следующие особенности размножения рыб:
· раздельнополость (есть самцы и самки);
· наличие у рыб только полового размножения (у рыб не встречается бесполое размножение);
· наружнее (внешнее) оплодотворение и развитие личинки вне организма самки (так происходит у подавляющего большинства рыб);
· очень большая плодовитость.
Особенности размножения подавляющего большинства рыб таковы. В яичниках самок рыб развивается икра, состоящая из отдельных икринок. В парных семенниках самцов образуется семенная жидкость (молоки), содержащая сперматозоиды. В период нереста самки через половое (которое находится около анального) отверстие выбрасывают (выметывают) в воду икру. Находящиеся рядом самцы рым поливают икру семенной жидкостью. Подвижные сперматозоиды достигают икринок (яйцеклеток) и происходит оплодотворение, в результате которого образуется зигота.
Нерестом называют поведение рыб в период размножения. В основе нереста лежат инстинкты.
Деление зиготы приводит к формированию личинки рыбы. Сначала она развивается в икринке, позже выходит из нее. При этом личинка продолжает питаться за счет запаса питательных веществ, находящихся в желточном мешке, которые находится на брюшной стороне личинки.
Когда личинка начинается питаться самостоятельно (обычно мелкими взвешенными в воде организмами — планктоном) и становится похожа на взрослую рыбу (но маленькую), она называется мальком.
Важной особенностью размножения рыб является большая плодовитость. Количество икринок, которая выметывает самка различных видов рыб, обычно колеблется от нескольких сотен тысяч до нескольких миллионов. Такая плодовитость связана с тем, что рыбы почти не проявляют заботу о потомстве, и большое количество икры, личинок и мальков гибнет, так и не достигнув взрослого состояния. Это происходит из-за того, что не вся икра оплодотворяется, ее поедают различные водные животные. Личинки и мальки также часто служат пищей различным животным.
Из вышеописанных особенностей размножения рыб есть исключения.
Так у некоторых рыб (чаще всего акул) встречается живорождение. При этом у них происходит внутреннее оплодотворение (семенная жидкость самцов попадает в половые пути самки рыбы). Икра не выметывается, а задерживается внутри самки, и там происходит развитие личинки. Самка рождает уже способных к самостоятельной жизни мальков. При этом их количество небольшое. Например, у акул максимум несколько десятков.
Также небольшое количество икринок наблюдается у рыб, проявляющих заботу о потомстве (колюшка, морской конек), даже при том, что у них наружнее оплодотворение. Родители защищают свое потомство от неблагоприятных факторов внешней среды и большинство мальков выживает. Поэтому нет необходимости в большом количестве икры.
В период размножения для рыб характерно определенное инстинктивное поведение. Они скапливаются в определенных местах (одни на мелководье, другие на песчаном дне, третьи на каменистом дне и т. п.), характерных для конкретного вида. Существует ряд видов рыб, которые называются проходными. Эти виды (например, лососи, кета, горбуша, осетр) живут в морях, а ходят на нерест в реки. Именно там есть необходимые условия для развития их икры и личинок. Существуют виды, совершающие миграции для размножения из рек в моря и океаны. Так угорь живет в реках, а на нерест уходит в море.
Стадии эмбриогенеза:
бластула, гаструла, закладка осевых органов
После того, как спермий принес в яйцеклетку центросому и недостающий гаплоидный набор хромосом, начинается первый этап эмбриогенеза – дробление.
Дробление – последовательное митотическое деление зиготы на клетки (бластомеры) без роста дочерних клеток до размеров материнской. Зигота в течение нескольких дней остается окруженной блестящей зоной, поэтому дочерние клетки с увеличением их количества уменьшаются в размерах и тесно прилегают друг к другу.
Каждому виду животных свойственен определенный тип дробления, обусловленный количеством и характером распределения желтка. Желток тормозит дробление. Часть зиготы, перегруженная желтком, дробится медленнее или не дробится вовсе. Поэтому дробление бывает полным или неполным.
Полное дробление может быть равномерным, если образующиеся клетки (бластомеры) равной величины, и неравномерным, если бластомеры разной величины. В зависимости от типа дробления у разных типов животных образуются разные типы бластул: у ланцетника – целобластула, у амфибий – амфибластула, у птиц – дискобластула и у млекопитающих – бластоциста.
Дробление зиготы человека начинается к концу первых суток и характеризуется как полное неравномерное асинхронное. В течение первых суток оно происходит медленно. Первое дробление (деление) зиготы завершается через 30 ч, в результате образуются два бластомера, покрытых общей оболочкой оплодотворения. За стадией двух бластомеров следует стадия трех бластомеров. Через 40 ч образуются 4 клетки (рис. 8.1).
С первых же дроблений зиготы формируются два вида бластомеров –темные и светлые. Светлые, более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг темных, которые оказываются в середине зародыша. Из поверхностных светлых бластомеров в дальнейшем развивается трофобласт, связывающий зародыш с материнским организмом и обеспечивающий его питание. Внутренние, темные, бластомеры формируют эмбриобласт, из которого образуется тело зародыша и некоторые внезародышевые органы (амнион, желточный мешок, аллантоис).
Число клеток эмбриобласта значительно меньше, чем в периферических отделах бластоцисты (в среднем 1 клетка на 10-12 клеток трофобласта).
После трех делений через 3 сутки после оплодотворения дробление идет быстрее и образуется морула (плотное скопление клеток), состоящая из 12-16 клеток, а к 4 дню – поздняя морула. На 4 сутки у эмбриона, состоящего из 32 бластомеров, между клетками появляется полость бластулы и начинает формироваться бластоциста – полый пузырек, заполненный жидкостью.
Морула, а затем бластоциста в течение 3 суток перемещается по яйцеводу к матке и через 5 сутки попадает в матку. Бластоциста находится в полости матки в свободном виде в течение двух дней (5-6 сутки) и эта стадия обозначается, как свободная бластоциста. К этому времени бластоциста увеличивается благодаря росту числа бластомеров (клеток эмбриобласта и трофобласта) до 100 клеток и более вследствие усиленного всасывания трофобластом секрета маточных желез, а также активной выработки жидкости самим трофобластом. Эмбриобласт располагается в виде узелка зародышевых клеток, который прикреплен изнутри к трофобласту на одном из полюсов бластоцисты.После попадания бластоцисты в матку на 6-7 сутки происходит имплантация – внедрение зародыша в слизистую оболочку матки. Имплантация продолжается около 40 ч. Период имплантации является первым критическим периодом развития зародыша. Одновременно с имплантацией начинается следующий этап эмбриогенеза – гаструляция.
Гаструляция – (от лат. gaster - желудок) - сложный процесс химических и морфогенетических изменений, сопровождающихся делением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки: наружный (эктодерма), внутренний (энтодерма), средний (мезодерма). Эти зародышевые листки вследствие дальнейшего развития дают начало эмбриональным зачаткам, из которых формируются ткани и органы. Эктодермаиэнтодерма образуются в результате деламинации (расслоения) зародышевого узелка, а мезодерма – путем иммиграции (выселения) клеток.
В процессе дальнейшего развития зародыша путем миграции клеток происходит образование мезенхимы – эмбриональной соединительной ткани, заполняющей пространства между зародышевыми листками и зачатками осевых органов.
Из зародышевых листков и мезенхимы происходит формирование всех тканей и органов развивающегося организма. Вначале закладываются осевые зачатки органов: нервная трубка, хорда, кишечная трубка.
Нервная трубка формируется из первичной эктодермы, хорда – из хордомезодермального зачатка, а эпителий кишечной трубки – из энтодермы.
Нейруляция – процесс образования нервной трубки – протекает по времени неодинаково в различных частях зародыша (рис. 8.3). Замыкание нервной трубки начинается в шейном отделе, а затем распространяется кзади и несколько замедленнее протекает в краниальном направлении, где формируются мозговые пузыри. Примерно на 25-е сутки нервная рубка полностью замыкается, с внешней средой сообщаются только два незамкнувшихся отверстия на переднем и заднем концах – передний и задний нейропоры. Задний нейропор соответствует нейрокишечному каналу. Через 5-6 суток нейропоры зарастают. Из нервной трубки образуются нейроны и нейроглия головного и спинного мозга, сетчатки глаза и органа обоняния.
При смыкании боковых стенок нервных валиков и образовании нервной трубки появляется группа нейроэктодермальных клеток, образующихся в области соединения нейральной и остальной (кожной) эктодермы. Эти клетки, сначала располагающиеся в виде продольных рядов по обе стороны между нервной трубкой и поверхностной эктодермой, образуют нервный гребень. Клетки нервного гребня способны к миграциям. В туловище одни клетки мигрируют в поверхностном слое дермы, другие – в вентральном направлении, образуя нейроны и нейроглию парасимпатических и симпатических ганглиев, мозговое вещество надпочечников. Часть клеток остается в области нервного гребня, формируя ганглиозные пластинки, которые сегментируются и дают начало спинномозговым узлам.
Из вторичной эктодермы формируется кожный эпителий. Из кишечной трубки развивается эпителий органов пищеварения, дыхания и некоторых органов эндокринной системы.
Мезодерма дифференцируется на сомиты, нефрогонотомы и спланхнотомы. Из дерматомов сомитов формируется соединительнотканная часть кожи, из миотомов – скелетные мышцы, из склеротомов – хрящи и скелет. Из нефрогонотомов развиваются эпителиальные зачатки почек и половых желез. Из листков спланхнотома образуется эпителий серозных оболочек внутренних органов, плевры, перикарда. Из мезенхимы развиваются кровь и лимфа, кроветворные органы, сосуды, соединительные ткани, гладкая мышечная ткань.
В стадии гаструляции и образования зачатков осевых органов происходит формирование внезародышевых органов, которые обеспечивают условия для нормального развития эмбриона. Они развиваются из внезародышевой части зародышевых листков и функционируют только в период эмбрионального развития. К ним относятся желточный мешок, амнион, аллантоис, серозная оболочка (у птиц), хорион и плацента (у млекопитающих).Характеристика основных морфологических процессов в различные периоды внутриутробного (пренатального) развития человека представлена в схеме:
| Срок развития, нед. | Морфогенетические процессы |
| Начальный период (ранний эмбриогенез) | |
| 1-я | Оплодотворение. Дробление зиготы. Образование морулы и бластулы. Первая стадия гаструляции (деламинация), образование эпибласта и гипобласта. Начало имплантации. |
| Зародышевый период (эмбриональный) | |
| 2-я | Завершение имплантации. Формирование зародышевого диска. Вторая стадия гаструляции (иммиграция), образование первичной полоски, прехордальной пластинки. Образование амниотического и зародышевого пузырьков, внезародышевой мезодермы. Дифференцировка трофобласта на цитотрофобласт и симпластотрофобласт, первичных ворсин хориона. Развитие первичного и вторичного (дефинитивного) желточного мешка. |
| 3-я | Продолжение 2-й стадии гаструляции, образование трех зародышевых листков, хорды, прехордальной пластинки, нервной трубки, нервного гребня. Начало сегментации дорсальной мезодермы (сомиты, сегментные ножки), образование париетального и висцерального листков спланхнотомов и эмбрионального целома, который далее разделяется на три полости тела - перикардиальную, плевральную, перитонеальную. Закладка сердца, кровеносных сосудов, предпочки - пронефроса. Формирование внезародышевых органов - аллантоиса, вторичных и третичных ворсин хориона. Образование туловищной складки и отделение первичной кишки зародыша от вторичного желточного мешка. |
| 4-я | Углубление желточной складки, образование желточного стебля и приподнятие зародыша в полости амниона. Продолжение сегментации дорсальной мезодермы до 30 сомитов и дифференцировка на миотом, склеротом и дерматом. Замыкание нервной трубки и формирование переднего невропора (к 25 сут.) и заднего невропора (к 27 сут.), образование нервных ганглиев; закладка легкого, желудка, печени, поджелудочной железы, эндокринных желез (аденогипофиза, щитовидной и околощитовидных желез). Образование ушной и хрусталиковой плакод, первичной почки — мезонефроса. Начало формирования плаценты. Образование зачатков верхних и нижних конечностей, 4-х пар жаберных дуг. |
| 5-я | Расширение головного конца нервной трубки. Окончание сегментации мезодермы (образование 42-44-х пар сомитов), образование несегментированной мезодермы (нефрогенная ткань) в каудальном отделе. Развитие бронхов и долей легкого. Закладка окончательной почки (метанефрос), урогенитального синуса, прямой кишки, мочевого пузыря. Образование половых валиков. |
| 6-я | Формирование лица, пальцев рук. Начало образования наружного уха и глазного яблока. Образование зачатков отделов головного мозга - моста, мозжечка. Формирование печени, поджелудочной железы, легких. Закладка грудных желез. Отделение гонад от мезонефроса, формирование половых различий гонад. |
| 7-я | Формирование верхних и нижних конечностей. Разрыв клоакальной мембраны. |
| 8-я | Формирование пальцев верхней и нижней конечностей. Значительное увеличение размеров головы (до 1/2 длины туловища). Пуповина. |
| Плодный период | |
| 9-я | Завершение формирования плаценты (12 — 13 нед.). Образование гладкого и ворсинчатого хориона. Разрастание симпластотрофобласта и редукция цитотрофобласта в ворсинах плаценты. Значительное увеличение размеров и массы плода. Продолжение процессов формирования тканей и органов. Формирование системы мать — плод. Кровообращение плода. |
Вопрос 10
· Эмбриональный период развития рыб не заканчивается выходом зародыша из оболочки. Он продолжается в течение некоторого времени после выклева, пока предличинка, или свободный эмбрион, обладая ещё рядом эмбриональных особенностей строения органов дыхания, кровооб-ращения, пищеварения и других систем, проходит заключительные этапы эмбрионального разви-тия. После того как начинают функционировать жаберная, пищеварительная и другие системы, деятельность эмбриональных органов прекращается и соответственно кончается период эмбрио-нального развития.
· Следующий период – личиночный – начинается с момента перехода молоди на активное питание внешней пищей. Сначала питание смешанное – остатками желточного мешка и частично внешней пищей, затем полностью экзогенное. Имеются временные личиночные органы (непарная плавниковая кайма, наружные жабры и т. д.), отсутствуют многие органы взрослой рыбы.
· При переходе в следующий период развития – мальковый молодь приобретает форму взрослой рыбы; появляется чешуя, характерные для взрослого органы и функции (например, брюшные плавники и жаберное дыхание через рот), но некоторые органы могут ещё отсутство-вать, например каналы боковой линии. Личиночные органы исчезают.
· Для примера рассмотрим развитие карпа в нерестовом пруду (при температуре воды 20–22°С рис. 38).
· В течение первых суток проходят этапы, предшествующие оформлению тела зародыша.
· Образование бластодиска (1-й этап). Начинается сразу после оплодотворения. Примерно через 30 мин в икринках между желтком и наружной оболочкой возникает перивителлиновое про-странство, занимающее 3,4–15,4 % диаметра икринки. На анимальном полюсе икринки формиру-ется бластодиск в виде возвышающегося над желтком светлого бугорка.
· Дробление бластодиска (2-й этап). Бластодиск разделяется бороздами дробления на бла-стомеры.
· Сначала наблюдается морула крупных клеток, но по мере того, как возрастает число бла-стомеров, размеры их уменьшаются. Примерно через 5 ч после оплодотворения наблюдается мо-рула мелких клеток.
· Бластула (3-й этап). Бластомеры уплотняются и отодвигаются к периферии. Образуется бластула, внутри которой имеется полость – бластоцель; желток образует впячивание навстречу накрывающей его бластодерме.
· Гаструла (4-й этап). При дальнейшем размножении клеток анимального полюса проис-ходит обрастание желтка: бластомеры как бы сползают в сторону вегетативного полюса, посте-пенно накрывая его; образуется зародышевый узелок; формируются зародышевые пласты, а из них зачатки органов.
· К концу первого дня после оплодотворения в икринке имеется зародыш в виде прозрач-ной зародышевой полоски, лежащей на желтке. Произошла закладка головного и туловищного зачатков, причём головной конец заметен резче, хвосто 74;ой конец утончается постепенно, огра-ничиваясь едва заметно; выявляются участки эмбрионального материала, которые дадут начало хорде, миотомам, кишечной энтодерме, нервной и другим системам.
· В течение вторых суток проходят следующие три этапа.
· Органогенез (5-й этап). Зародыш увеличивается в размерах: тело утолщается, хвостовой отдел оканчивается перед головным, немного не доходя до него. Формируются головной, туло-вищный, хвостовой отделы тела и основные органы и системы органов: нервная, мышечная, ки-шечник и т. д. Примерно через 28 ч после оплодотворения в головном отделе хорошо виден мозг, причем заметно разделение его на передний и задний отделы, четко различимы слуховые пузырь-ки, глаза продолговатой формы, ещё не имеющие пигмента. В туловищном отделе происходит сегментация хорды. Примерно через 32 ч. после оплодотворения хорошо заметна плавниковая кайма, начинающаяся на спинной стороне тела в задней его трети. Кайма огибает хвостовой отдел и подходит к желтку. Видны также плавниковые складочки на желтке.
· Появляется нервно-мышечная моторика (6-й этап). Зародыш начинает временами подер-гиваться, а затем периодически поворачивается в оболочке. Так как зародыш в это время дышит поверхностью тела (специальных органов дыхания нет), то перемешивание перивителлиновой жидкости при таких поворотах способствует улучшению газового обмена.
· Зародыш настолько увеличивается, что хвостовой отдел начинает заворачиваться по по-верхности желтка, образуя спираль. В головном отделе просматриваются обонятельные ямки, глазные бокалы, хрусталики, отолиты. В глазах появляется точечный меланин. Сердечная трубка сокращается, но форменных элементов крови ещё нет. Хорошо видна кишечная трубка. Продол-жается сегментация тела (в хвостовом отделе). Желточный мешок становится грушевидным.
· Начинает функционировать эмбриональная дыхательная система (7-й этап). Так как де-финитивные органы дыхания ещё не сформированы, то дыхательную функцию выполняет сеть кровеносных сосудов: Кювьеровы протоки (лежащие на передней части желточного мешка), ниж-няя хвостовая вена (в хвостовом отделе тела), сеть сегментальных сосудов в плавниковой кайме (в анальной ее части). В токе плазмы крови появляются форменные элементы. Заканчивается сегмен-тация тела. Появляются грудные плавнички. Усиливается пигментация глаз. Примерно через 52 ч после оплодотворения появляются пигментные клетки над кишечной трубкой, вскоре покрываю-щие головку зародыша, спинной и хвостовой отделы и желточный мешок. Пигментные клетки (меланофоры) крупные, лежат близко друг к другу (группами). На голове видны зачатки жаберных крышек. На голове и желтке появляются железки вылупления.
· К концу вторых – началу третьих суток после оплодотворения начинается последний – 8-й этап развития зародыша в оболочке. Увеличиваются все части тела и просвечивающие сквозь прозрачные покровы органы. Головка зародыша частично обособляется от желтка. В слуховых пузырьках видны полукружные каналы. Отчетливо видна ротовая ямка (рот неподвижный, откры-тый). Оформляется жаберно-челюстной аппарат. В передней части головы видны клетки, образу-ющие железки приклеивания. Основания грудных плавников расположены наклонно по отноше-нию к оси тела. В плавниковой складке обособляются спинной, хвостовой и анальный участки. Усиливается пигментация тела.
· Примерно через 78 ч после оплодотворения начинается массовый выклев молоди. Вы-клюнувшиеся зародыши, или предличинки (этап развития А, или последний зародышевый), имеют около 5,0–5,2 мм длины (рис. 39). Обращает на себя внимание большой желточный мешок груше-видной формы и прямая (не изогнутая) хорда. Голова немного пригнута вниз. В передней части ее, ближе к глазам, имеются углубления – обонятельные ямки. Хорошо видны сегменты (их насчиты-вается 38), не одинаковые по величине, они постепенно уменьшаются к заднему концу тела. По спине зародыша, начиная с 9-го сегмента, тянется вдоль тела плавниковая кайма, переходящая на хвост, далее на брюшную сторону и оканчивающаяся на желточном мешке. В хвостовой части плавниковая кайма разделяется задним концом хорды на две равные половины. Плавниковая кай-ма узкая, недифференцированная, без выемок, расширяется только в хвостовой части, прозрачная, чуть-чуть уплотненная с прилегающей к телу стороны; в спинной и анальной частях пронизана кровеносными сосудами. Грудные плавнички подвижны. Глаза сильно пигментированы. По телу разбросаны пигментные клетки; больше всего их на голове и вдоль спинного и брюшного краев тела, лежат они и на желточном мешке. На голове и спине имеется также желтоватый пигмент.
· На переднем краю головы зародыши имеют железу приклеивания, позволяющую им прикрепляться к подводным растениям. Сквозь прозрачное тело просвечивают внутренние орга-ны: сердце в околосердечной сумке, кишка, прямая, ещё без просвета, не вполне сформированный жаберный аппарат – только начальные жаберные дужки прикрыты намечающейся жаберной крышкой, два отолита в слуховой капсуле. Рот открыт, имеет форму ямки.
· В течение 1-х суток жизни после выклева зародыши движутся периодически; время от времени, приклеившись к растениям, они висят неподвижно, покойно; затем, оторвавшись от суб-страта, проделывают несколько червеобразных движений, после чего опять приклеиваются. Таким образом чередуются состояния движения и покоя.
· При указанных температурах преобразования зародышей протекают быстро. Уже к кон-цу первого дня их жизни (длина 6 мм) желточный мешок оказывается сильно втянутым. На 2-е сутки жизни (длина 5,9–6,7 мм) зародыши имеют сравнительно небольшой желточный мешок. Уменьшение желточного мешка происходит по всей площади соединения его с зародышем, но быстрее в передней расширенной части. В плавниковой кайме, особенно в нижней части хвостово-
го отдела, уплотнённые участки (скопления мезенхимных клеток) становятся более значительны-ми. Зародыши больше не приклеиваются к растениям, они постоянно плавают.
· На 3-й сутки жизни (этап развития В, или первый личиночный) при длине тела 6,2–7,8 мм у молоди остается совсем мало желтка. Хорда по-прежнему оканчивается прямо – она не изо-гнута. У особей длиной около 7 мм дифференциации плавниковой каймы ещё нет, но в хвостовой части в нижней половине намечаются мезенхимные тяжики. Пигментных клеток становится больше. Жаберная крышка прикрывает не все жаберные дужки. Линия основания грудных плав-ников становится вертикальной. Кровь начинает окрашиваться, приобретает очень слабый жёлто-розовый оттенок.
· Кишечник представляет собой едва изогнутую трубку, но уже с просветом. Молодь за-глатывает воздух, плавательный пузырь (задняя камера) наполняется им и становится хорошо видным. Наполнение плавательного пузыря воздухом облегчает передвижение рыбок. Части рото-вого аппарата могут двигаться. Рот перемещается на конец рыла.
· Молодь переходит к активному питанию (внешней пищей). Таким образом, в это время у личинок питание смешанное: как внешней пищей, так и за счет не совсем израсходованного жел-точного мешка. Вследствие прозрачности тела хорошо видно содержимое кишечника.
· На 4-е сутки жизни (этап развития С1; или второй личиночный)длина личинок достигает 5,5–9,0 мм. Самые мелкие из них имеют ещё остатки желтка. Рот приобретает способность закры-ваться полностью.
· У личинок, достигших длины около 8,3 мм, задний конец хорды – уростиль – начинает загибаться кверху. Зачатки лучей в нижней половине хвостовой части плавниковой каймы увели-чиваются. Плавниковая кайма в передней части (на спине) становится более высокой, здесь появ-ляется сгущение мезенхимных клеток. Такое же сгущение мезенхимы наблюдается в анальной ча-сти каймы, на месте будущего анального плавника. Перед хвостом плавниковая кайма становится немного уже, тем самым намечаются границы хвоста.
· Пигментных клеток становится очень много, они крупные, разбросаны по всему телу. Особенно крупны они на спинной стороне головы. Жаберные крышки увеличиваются. Личинки уже заглатывают циклопов, босмий и других мелких ветвистоусых и веслоногих рачков.
· На 5-е сутки жизни (этап развития С2, или третий личиночный)при длине 7,0–10,1 мм личинки отличаются от предыдущих в основном тем, что у них сильнее загнут уростиль, хвост стал гетероцеркальным, в плавниковой кайме резче выделяется хвостовой отдел, в котором лучи уже сформировались; в спинном и анальном отделах плавниковой каймы сгущения мезенхимы стали плотнее. На челюстях появляются роговые зубы. В пищевом комке кроме коловраток, вет-вистоусых и веслоногих рачков начинают встречаться планктонные личинки хирономид.
· На 6-е сутки жизни (длина 8,2–11,3мм) личинки своим общим видом напоминают уже больше рыбку, чем личинку. Головка из закругленной становится вытянутой. Жаберные крышки закрывают все жаберные дужки. Хвостовой отдел на плавниковой кайме ограничивается четче, мезенхимные сгущения в спинном и анальном участках каймы уплотняются. Тело личинок стано-вится менее прозрачным, сегменты видны плохо, только в задней части.
· На 8-е сутки жизни личинки достигают длины 10–12,8 мм. Меньшие из них прошли этап развития D1, , или четвертый личиночный, у наиболее крупных развитие продвинулось до этапа D2 –пятого личиночного.
· Сегменты в теле видны совсем плохо. Уростиль сильно загибается кверху, образуя почти прямой угол с плавниковыми лучами. В плавательном пузыре обе камеры наполнены воздухом. На месте брюшных плавников появляются кожистые выросты. Плавниковая кайма ясно диффе-ренцирована, спинной отдел ее имеет лучи, в анальном ее отделе также появляются зачатки лучей. Хорошо видны кости черепа. У наиболее крупных рыбок хвостовой отдел представляет собой сформированный хвостовой плавник (гомоцеркальный); появляется хвостовая выемка, раздваива-ющая плавник на верхнюю и нижнюю лопасти. Спинной плавник также вполне сформирован. Грудные и брюшные плавники ещё не имеют лучей. Все тело очень сильно пигментировано. Рот становится выдвижным. Кишечник слабо изогнут, намечается первая петля.
· На 11-е сутки жизни (длина 11,1 – 16,0 мм) у личинок спинной и анальный участки плав-никовой каймы приобретают форму плавников. С хвостовым плавником они соединяются совсем узенькими перетяжками. Лопасти брюшных плавников становятся крупнее, но лучей в них ещё нет. Пигментные клетки очень крупные. В кишечнике образуется первая петля.
· На 13-е сутки жизни (длина 12–13 мм) остатки плавниковой каймы между плавниками становятся еле заметными. Тело почти непрозрачно, лишь слабо просвечивает кишечник.
· На 14-е сутки жизни (этап развития Е, или шестой личиночный)при длине тела 15–20 мм никаких следов плавниковой каймы между плавниками нет. В брюшных и грудных плавниках по-
явились лучи. Тело непрозрачно, его почти сплошь покрывают пигментные клетки. В кишечнике стало две петли. Чешуи ещё нет. Дальнейшее развитие происходит в выростном пруду.
· В течение развития у зародыша чередуются периоды усиленного роста тканей и 87;ериоды усиленной дифференцировки их и образования новых зачатков органов. При этом ме-няется характер обмена веществ, в частности интенсивность водного обмена, интенсивность усво-ения биогенных элементов (фосфора, кальция, углерода), аминокислотный состав тела (уменьша-ется число свободных аминокислот, увеличивается количество связанных), интенсивность потреб-ления кислорода. Наиболее интенсивен обмен во время формирования органов и тканей. Чувстви-тельность зародышей к внешним воздействиям – тряске, колебаниям температуры, содержанию кислорода – на разных стадиях развития различна. Наименее устойчивы зародыши во время уси-ленного формирования тканей и органов, когда обмен наиболее интенсивен. Это начало дробле-ния, гаструляция, закрытие бластопора, начало формирования зародыша и т. д. Это обстоятельство учитывается при работах с икрой, особенно при ее перевозках.
· Инкубация икры рыб каждого вида проходит при определённых условиях внешней сре-ды (температура, содержание кислорода и углекислоты, рН, освещенность, соленость и т. д. ).
· Исход инкубации определяется также качеством икры. Оно связано с видом рыбы, ее возрастом, условиями содержания и в предыдущий год, и особенно перед нерестом, временем взя-тия икры от самок, а при искусственном осеменении – с техникой проведения всех операций.
· Длительная задержка икры в полости тела самки вызывает перезревание. Перезревшая икра характеризуется пониженной оплодотворяемостью, повышенным отходом в период инкуба-ции, увеличенным числом уродов и самцов.
· Невыметанные зрелые половые продукты рассасываются. Но это длительный процесс. Поэтому если нерест почему-либо не прошел (например, из-за похолодания), то очередной нерест на следующий год также может не состояться, так как в гонадах не успеют пройти процессы ре-зорбции зрелых невыметанных клеток и образования клеток новых генераций.
· Продолжительность инкубации при прочих равных условиях зависит от температуры: чем она выше, тем развитие происходит быстрее.
· У рыб, выметывающих икру весной и летом, при высоких температурах, развитие длится несколько дней; у рыб с осенне-зимним нерестом – несколько месяцев.
· Успешнее всего инкубация проходит при оптимальной температуре. При повышенной она хотя и заканчивается быстрее, но молодь выклевывается мелкой и недоразвитой, а при пони-жении температуры зародыши более крупные, но нарушается процесс выклева. При отклонении температуры повышается количество уродов –особей с укороченным туловищем, искривлением позвоночника, водянкой околосердечной и брюшной полости, а также двухголовых экземпляров, срастающихся разными участками ту 83;овища, особей с аномалиями челюстного аппарата и т. д.
· Для учета длительности развития существует понятие ‛градусо-дни‛. Это произведение средней температуры инкубации на число дней развития икры. Оно дает общ 77;е представление о сумме тепла, необходимого для развития молоди до выклева. Но это не постоянная величина, она имеет разные значения при разных температурах. У карпа развитие длится 54–126 градусо-дней, у радужной форели – 330–400. При неблагоприятных условиях, например при недостатке кислоро-да, продолжительность развития удлиняется.
Дата добавления: 2018-02-28; просмотров: 3317; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!