Классификация вирусов по Балтимору
| класс | геном и репликация | примеры |
| I | dsDNA | Adenoviridae , Herpesviridae , Poxviridae, Iridoviridae |
| II | ssDNA | Parvoviridae |
| III | dsRNA | Reoviridae |
| IV | ssRNA + | Picornaviridae, Togaviridae |
| V | ssRNA - | Orthomyxoviridae, Rhabdoviridae |
| VI | ssDNA +, DNA-intermediate | Retroviridae |
| VII | dsDNA, RNA-intermediate | Hepadnaviridae |
Ценность системы Балтимора состоит в том, что она позволяет связать структуру генома вируса с процессом его размножения в клетке. В другой, более наглядной форме эта система представлена на рисунке 4.
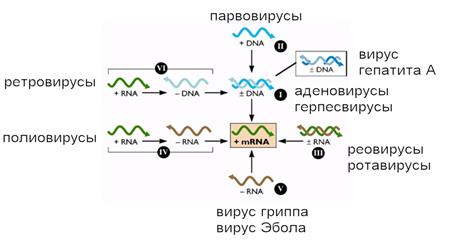
Рис. 4. Система Балтимора показывает, что центральной молекулой в репликации вирусов является мРНК.
Система Балтимора является важным дополнением к основной классификации вирусов, которая учитывает форму и размеры вириона, особенности вирусного генома и место сборки вирионов в клетке. Главным следствием из этой системы является заключение, что все вирусы для того, чтобы обеспечить репликацию своей нуклеиновой кислоты и синтез вирусных белков, должны на основе своего генома генерировать +цепь мРНК.
Конкретные механизмы репликации нуклеиновых кислот могут сильно отличаться у различных семейств. Например, большинство ДНК-содержащих вирусов кодируют полимеразы в своем геноме. Исключение составляют только Parvoviridae и Papovaviridae . Зависимость от клеточных полимераз накладывает значительные ограничения на их распространение, поскольку они могут реплицироваться только в пролиферирующих клетках.
|
|
|
Репликация у ДНК-содержащих вирусов осуществляется одним из трех способов:
· Двунаправленная репликация кольцевой ДНК (Papovaviridae).
· Последовательная репликация линейной ДНК (Adenoviridae).
· Репликация с помощью РНК-посредника (Hepadnaviridae).
У РНК-содержащих вирусов способы репликации более разнообразны. Наиболее характерные для них особенности заключаются в следующем:
· +РНК-геном может функционировать как мРНК, для –РНК-генома для этого требуется синтезировать комплементарную цепь;
· Все РНК-содержащие вирусы кроме Retroviridae кодируют собственную РНК-зависимую РНК-полимеразу. Если у вируса –РНК-геном, ему надо обеспечить вход в клетку не только нуклеиновой кислоты, но и полимеразы.
· Все РНК-содержащие вирусы реплицируются в цитоплазме. Исключение составляют Retroviridae и некоторые другие.
· Семейство Retroviridae уникально. Эти вирусы имеют +РНК-геном, однако он конвертируется в двуцепочечный ДНК-геном содержащейся в вирионе обратной транскриптазой (РНК-зависимой ДНК-полимеразой). ДНК-геном встраивается затем в геном клетки с помощью вирусной интегразы и реплицируется клеточной РНК-полимеразой II.
|
|
|
Созревание новых вирионов. Специфические вирусные вещества и структуры часто синтезируются в цитоплазме или/и ядре клетки в составе вирусорепликативных комплексов, или виропластов. Особенно крупные и сложные виропласты формируются в цитоплазме клетки при инфицировании ДНК-содержащими вирусами семейств Poxviridae и Iridoviridae. В электронном микроскопе они выглядят как обширные скопления мелкодисперсного материала, по периферии которых располагаются вирионы на различных стадиях морфогенеза, а также рибосомы и линейные полисомы.
Морфогенез РНК-содержащих вирусов семейства Reoviridae также происходит в цитоплазматических виропластах. У ДНК-содержащих вирусов семейства Herpesviridae нуклеокапсиды образуются в виропластах, локализованных в ядре клетки, тогда как оболочки вирионов – из модифицированных вирусными белками клеточных мембран.
Многие РНК-содержащие вирусы размножаются в клетке без образования виропластов. К ним относятся вирусы семейств Ortomyxoviridae, Paramyxoviridae, Rhabdoviridae и др.
Выход вирионов нового поколения из клетки. Вирионы нового поколения выходят из клетки путем ее лизиса или почкованием на плазмалемме. Оба процесса приводят к изменениям в плазмалемме, которые можно наблюдать в микроскоп как проявление ЦПЭ.
|
|
|
Вирус при его выходе из клетки сталкивается с проблемой связывания новых частиц со специфическими для них клеточными рецепторами. Вирусы семейств Paramyxoviridae и Orthomyxoviridae решают эту проблему с помощью нейраминидазы (сиалидазы), которая отщепляет остатки сиаловой кислоты при выходе вириона. Ингибирование этого фермента лежит в основе фармакологического действия таких противовирусных препаратов как занамивир и озельтамивир. Эти вещества подавляют выход вирионов из клетки, что приводит к их агрегации на клеточной поверхности.
Многие вирусы выходят из клеток не полностью зрелыми и не способными инфицировать другие клетки. Примером может служить ВИЧ-1, созревание которого вне клетки требует участия вирусной протеазы, ремоделирующей ряд его компонентов. Если ингибировать эту протеазу, вирионы ВИЧ-1 не приобретают инфекционность.
Вирусные включения
Вирусные включения представляют собой комплексы структур, которые наблюдаются в инфицированной вирусом клетке в световой микроскоп. По реакции на красители они подразделяются на базофильные и оксифильные (эозинофильные), по структуре – на гомогенные и зернистые, по локализации – на цитоплазматические и внутриядерные.
|
|
|
Включения встречаются при многих вирусных инфекциях, имея в отдельных случаях диагностическое значение. Обычно они представляют собой округлые образования диаметром 1-10 мкм, окруженные светлой каймой. Количество, размеры, структура и другие свойства включений могут изменяться по мере развития вирусной инфекции. В пораженной клетке может быть несколько включений различных размеров и формы. Обычно они прилегают к ядру, отодвигая его к периферии или окружая его. Для каждого тельца-включения характерна внутренняя структура.
Большинство включений, образующихся при инфицировании клетки ДНК-содержащими вирусами, также содержит ДНК. Хорошо изучены включения при таких вирусных болезнях животных как оспа птиц (тельца Боллингера), болезнь Борна (тельца Дегена и Оста), чума собак (тельца Лектура), инфекционном ларинготрахеите птиц (тельца Зейфрида) и т.п.
Цитоплазматические включения при оспе разных видов животных и птиц имеют много общего. При окраске гематоксилином и эозином включения обнаруживаются в виде овальных образований величиной, примерно равной величине ядрышка клетки. Центральная их часть эозинофильна, с неровным контуром. Каждое включение по периферии окружено светлой зоной, или мантией. Впервые эти включения наблюдал итальянский врач Гварньери в 1882 г. В 1896 г. Боллингер обнаружил их при оспе кур, а в 1903 г. Боррель – при оспе овец. Они обнаруживаются в световом микроскопе, главным образом, в пораженных участках кожи..
Внутриядерные включения встречаются при инфекциях, вызываемых как относительно крупными вирусами (герпес, болезнь Ауески, ринопневмония лошадей), так и вирусами, имеющими очень мелкие размеры (ящур, гепатит собак). Ядерные включения лучше выявляются в гистологических препаратах, причем их форма и размеры зависят от штамма вируса. Различают внутриядерные включения типа А и В. Включения типа А представляют собой гомогенные округлой формы оксифильные образования, четко отграниченные от базофильного хроматина. Включения типа А встречаются при ринопневмонии лошадей, ларинготрахеите птиц, болезни Ауески и др. Включения типа В имеют вид оксифильных гиалиновых зерен, структура ядра при этом изменяется мало. Они наблюдаются, в частности, при болезни Тешена и аденовирусной инфекции.
При одной и той же болезни характер вирусных включений может отличаться. Так при кори формирование цитоплазматических включений сопровождается накоплением нового поколения вирионов, тогда как развитие внутриядерных включений рассматривают как реакцию клетки на вирус.
В таблице 6 представлены примеры вирусных включений, обладающих патогномоничностью (т.е. специфичностью по отношению к системе клетка-вирус).
| Инфекция | семейство | Включени z |
| цитоплазматические | ||
| Бешенство | Rhabdoviridae | Тельца Бабеша-Негри |
| Натуральная оспа | Poxviridae | Тельца Гварниери |
| Птичья оспа | Poxviridae | Тельца Боллингера |
| внутриядерные | ||
| Герпесвирусы | Herpesviridae | Включения типа A |
| Аденовирусы | Adenoviridae | Включения типа В |
| Желтая лихорадка | Phlaviviridae | Тельца Торреса |
| цитоплазматические и внутриядерные | ||
| Корь | Paramyxoviridae | Индуцируют синцитий |
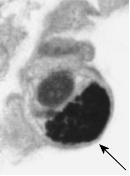
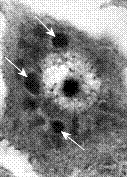
| 
|
| Включение Гварниери при оспе | Включения Бабеша-Негри при бешенстве |
|
| |
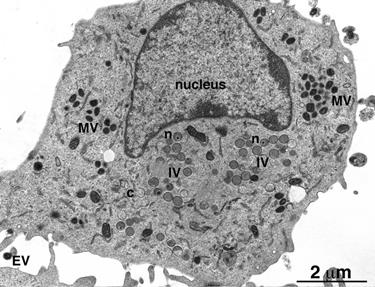
Рис. 5. Световая микроскопия включений Гварниери при оспе и включений Негри при бешенстве. Внизу показана электронная микрофотография клетки с формирующимися в зоне виропласта вирионами вируса оспы: IV – незерелые вирионы, MV – зрелые внутриклеточные вирионы, EV – внеклеточный вирион, который вышел из клетки путем почкования на плазмалемме.
Природу вирусных включений исследуют с помощью электронного микроскопа. Так, было установлено, что во многих случаях включения представляют собой места образования и накопления вирусных частиц. Примером могут служить тельца Гварниери в цитоплазме клеток, зараженных вирусами семейства Poxviridae. Эти структуры обозначаются как виропласты, или вирусные фабрики. Они характерны в основном для ДНК-содержащих вирусов, которые размножаются в цитоплазме. В то же время тельца Бабеша-Негри представляют собой скопления нуклеокапсидов вируса бешенства.. Включения типа А, формирующиеся в ядрах клеток, зараженных вирусом герпеса, состоят из плотных глыбок хроматина. Внутриядерные включения типа В, которые встречаются при аденовирусной инфекции, представляют собой паракристаллические скопления вирионов, имеющих форму икосаэдра.
При ряде вирусных инфекций обнаружение включений имеет диагностическое значение. В частности, обнаружение их стало одним из методов диагностики бешенства, оспы, ринопневмонии лошадей, аденовирусной инфекции и т.п. В диагностике гриппа животных, болезни Ауески, ларинготрахеита птиц и других болезней выявление телец включений играет вспомогательную роль. На частоту выявления телец – включений, кроме различий штаммов влияет возраст организма и его физиологическое состояние.
Дата добавления: 2022-06-11; просмотров: 48; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!