Цитопатический эффект вирусов
Вирусная цитопатология
Структура и классификация вирусов
Вирусы – это неклеточная форма жизни, которая существует благодаря тому, что они являются облигатными внутриклеточными паразитами. Геном вирусов представлен молекулами ДНК или РНК, которые упакованы в оболочку из белков. Размножение вирусов обеспечивается биосинтетическими системами эукариотических и прокариотических клеток. Полная частица вируса называется вирион. Основной функцией вириона является доставка ДНК или РНК вируса в клетку, где вирусный геном экспрессируется и контролирует воспроизведение вируса.
Вирусы относятся к самым примитивным формам жизни. Об этом убедительно свидетельствуют размеры их генома (табл. 1).
Размеры генома живых организмов
| Группа | Вид | Число генов |
| Вирусы | ||
| РНК-вирусы | Вирус иммунодефицита человека | 9 |
| ДНК-вирусы | Вирус оспы | 199 |
| Прокариоты | ||
| Микоплазмы | Mycoplasma genitalium | 470 |
| Риккетсии | Rickettsia prowazekii | 834 |
| Археобактерии | Archaeoglobus fulgidus | 2436 |
| Цианобактерии | Synechocystis sp. | 3168 |
| Эубактерии | Escherichia coli | 4288 |
| Эукариоты | ||
| Грибы | Saccharomyces cerevisiae | 6241 |
| Простейшие | Dictyostelium discoideum | 11000 |
| Высшие растения | Arabidopsis thaliana | 27540 |
| Беспозвоночные | Drosophila melanogaster | 13600 |
| Позвоночные | Homo sapiens | 28000 |
Следует отметить, что в последние годы обнаружены вирусы, величина генома которых достигает нескольких тысяч, что ставит их на один уровень с бактериями. Однако по сравнению с большинством вирусов они выглядят экзотически.
|
|
|
Вирусы классифицируют на основе таких критериев как форма и размер вириона, химический состав и структура генома, место и особенности репродукции в клетке.
Большинство вирусов имеют два типа симметрии вириона – кубический и спиральный. Кубический тип симметрии представлен правильным многогранником – двадцатигранником, или икосаэдром. Он состоит из большого числа белковых глобул –капсомеров (рис. 1А). Вирион спирального типа симметрии представляет собой трубку, которая образована спирально закрученной фибриллой из капсомеров (рис 1Б). Образованная капсомерами оболочка носит название капсид, или нуклеокапсид.

|

|
| А | Б |
Рис. 1. Два типа симметрии вирионов: А – кубический тип симметрии (икосаэдр); Б – спиральный тип симметрии (полая трубка).
Нуклеокапсид в форме икосаэдра характерен для большинства ДНК-содержащих вирусов и части РНК-содержащих вирусов. Спиральный тип симметрии обнаруживается у вируса табачной мозаики (ВТМ). Вирионы ВТМ представляют собой белковую трубку со спирально завитой молекулой РНК внутри. Спиральный тип симметрии обнаруживается также у многих нитевидных и плеоморфных вирусов. Вирионы ряда РНК-содержащих вирусов имеют шаровидную или вытянутую форму, представляя собой дериват плазматической мембраны клетки со встроенными вирусными белками. Вне зависимости от типа симметрии нуклеокапсид может быть дополнен мембраной или другими структурами.
|
|
|
Примером вириона комбинированной структуры может служить вирион вируса иммунодефицита человека (ВИЧ). Нуклеокапсид ВИЧ имеет кубический тип симметрии, он покрыт снаружи мембраной клеточного происхождения (рис. 2).
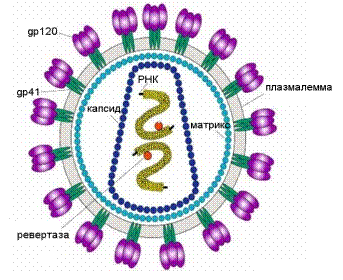
Рис. 2. Структура вириона вируса иммунодефицита человека: gp41 и gp120 являются
белками оболочки, под мембраной локализуется вирусный М-белок. Молекулы РНК
упакованы в белковый нуклеокапсид.
Геном ВИЧ содержит всего три структурных гена – gag, pol и env. Гены ВИЧ кодируют, однако, девять белков, что обеспечивается сплайсингом.
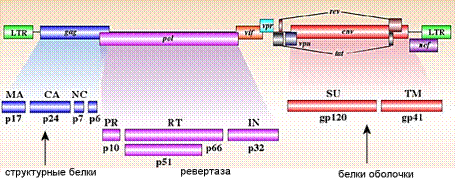
Рис. 3. Структура генома вируса иммунодефицита человека.
Вирусы человека и животных, как уже отмечалось, подразделяются на ДНК-содержащие и РНК-содержащие. Классификация ДНК-содержащих вирусов представлена в таблице 2.
|
|
|
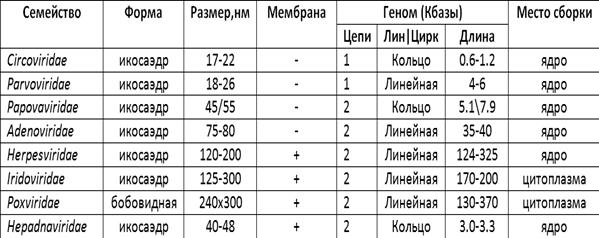
Из представленных в таблице данных следует, что вирионы большинства ДНК-содержащих вирусов имеют икосаэдра. Размеры вирионов варьируют в пределах от 17 до 300 нм. ДНК почти у всех двойная. Исключение составляют вирусы семейств Circ a viridae и Parvoviridae, у которых ДНК одинарная. Размеры генома колеблются в широких пределах: от 0.6 до 370 килобаз. Многие ДНК-содержащие вирусы имеют мембрану, причем у семейства оспы и вирусов радужности она собственного происхождения. Сборка всех ДНК-содержащие вирусов кроме последних двух семейств происходит в клеточном ядре. Вирусы семейства Papovaviridae способны вызывать опухолевую трансформацию клеток.
Классификация РНК-содержащих вирусов представлена в таблице 3.
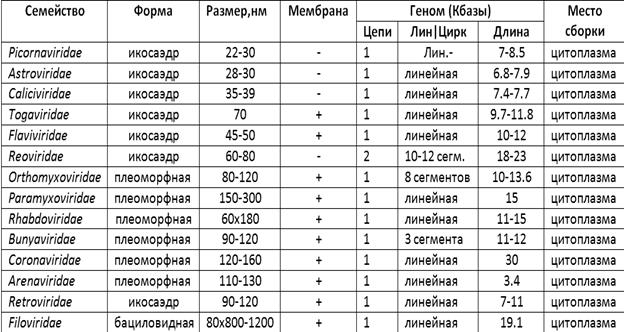
В отличие от ДНК-содержащих вирусов РНК-содержащие вирусы более многочисленны и разнообразны. У половины семейств форма вириона в виде икосаэдра, вирионы остальных семейств отличаются большой изменчивостью, варьируя от шаровидной до нитевидной. Размеры вирионов колеблются от 22 до 300 нм аналогично ДНК-содержащим вирусам. Размеры генома у РНК-содержащих вирусов, однако, значительно меньше, составляя от 3.4 до 30 килобаз. У всех вирусов, кроме Reoviridae, РНК одинарная, но может быть представлена как "+"-, так и "–"- цепью. У некоторых вирусов (семейства Reoviridae, Orthomyxoviridae, Bunyaviridae) геном сегментирован, т.е. отдельные гены не связаны между собой. Репликация всех РНК-содержащих вирусов происходит в цитоплазме.
|
|
|
Цитопатический эффект вирусов
Несмотря на большое разнообразие клеток и вирусов выделяют три основные формы их взаимодействия: цитопатическая, вирусопатическая аллобиофорическая.
Цитопатическая форма взаимодействия вируса и клетки всегда заканчивается ее деструкцией и гибелью. Вирусный геном может функционировать в клетке автономно, реплицируясь независимо от клеточного генома. В этом случае сформируется новое поколение полноценных вирусных частиц. Возможна, однако, и абортивная вирусная инфекция, когда цикл размножения вируса прерывается на какой-либо стадии и полноценного вирусного потомства не образуется. В основе патологических изменений клеток, инфицированных вирусом, лежат особенности его внутриклеточного онтогенеза.
Вирусопатическая форма взаимодействия вируса и клетки отличается сохранением клетки и деструкцией вируса. Атакующие клетку вирионы могут инактивироваться на клеточной поверхности и в процессе эндоцитоза. Одним из факторов, обеспечивающих устойчивость клеток к вирусу является продукция ими интерферона.
Аллобиофорическая форма взаимодействия клетки с вирусом представляет собой их эндосимбиоз с различной степенью интеграции геномов. Крайними вариантами этой формы являются интеграция вирусного генома или его ДНК-транскрипта с ядерным геномом или геномами митохондрий и хлоропластов, с одной стороны, или персистенция вируса в популяции клеток, с другой. Аллобиофорическая форма взаимодействия обеспечивает длительное совместное сохранение и размножение вирусов и клеток в природе.
Существует множество переходных форм взаимодействия вируса и клетки. В частности, онкогенные вирусы могут интегрировать в ядерный геном отдельные гены, что приводит к опухолевой трансформации клеточного клона.
Цитопатический эффект вируса (ЦПЭ) обозначает повреждение и разрушение популяций чувствительных к вирусу клеток. ЦПЭ рассматривается как завершающий этап сложного и многоступенчатого процесса деструкции клеток вследствие размножения в них цитопатогенного вируса
Выделяют следующие морфологические типы ЦПЭ:
1. Равномерная мелкозернистая деструкция клеточного монослоя. Такая картина поражения чувствительных культур клеток характерна для вирусов полиомиелита, Коксаки, ECHO, ECMO, оспы, гриппа.
2. Очаговая мелкозернистая деструкция с тяжами сохранившихся клеток. Этот тип ЦПЭ свойственен вирусам клещевого энцефалита и псевдобешенства, пенящему вирусу.
3. Гроздевидные скопления пораженных клеток, характерные для аденовирусной инфекции. Клетки при этом типе ЦПЭ могут долго не отпадать от стекла.
4. Равномерная крупнозернистая деструкция клеточного монослоя. Характерна для инфекции чувствительных культур клеток герпесвирусами.
5. Формирование гигантских многоядерных клеток – симпластов. Такой тип ЦПЭ наблюдается, в частности, при инфекции вирусами кори и осповакцины.
В основе ЦПЭ лежат молекулярно-генетические механизмы взаимодействия вируса с клеткой. При продуктивной вирусной инфекции выделяют следующие стадии взаимодействия вируса и клетки:
1. Адсорбция вириона на плазмалемме;
2. Проникновение вириона внутрь клетки;
3. Освобождение вирусной нуклеиновой кислоты;
4. Синтез вирусных нуклеиновых кислот и белков;
5. Созревание новых вирионов;
6. Выход вирионов нового поколения из клетки.
Адсорбция вириона на плазмалемме. Вирусы обычно прикрепляются к клеткам, используя рецепторы на ее плазмалемме. Эти рецепторы часто выступают над поверхностью клетки, облегчая процесс адсорбции. Среди рецепторов, которые используются вирусами, встречаются самые различные молекулы – поверхностные гликопротеины, компоненты сигнальных путей, внеклеточного матрикса и т.п. Некоторые рецепторы, в частности, сиаловые кислоты или протеогликаны, могут использоваться вирусами, относящимися к разным семействам. Для ряда вирусов кроме основного рецептора для прикрепления к клетке дополнительно необходим также корецептор, роль которого состоит в завершении процесса адсорбции. Одним из наиболее исследованных случаев такого типа является адсорбция ВИЧ-1 на Т-лимфоцитах "хелперах". Сначала рецептор CD4 на поверхности клеток контактирует с гликопротеином вирусной оболочки gp120, затем у последнего открывается центр связывания для корецептора, в качестве которого выступают CCR5, CXCR4 и другие рецепторы к лимфокинам. Примеры рецепторов плазмалеммы, которые используются вирусами для прикрепления к клетке, представлены в таблице 4.
Дата добавления: 2022-06-11; просмотров: 13; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!