ПОНЯТТЯ ПРО ІНДИВІДУАЛЬНИЙ РОЗВИТОК ЯК ПРОЦЕС ОНТОГЕНЕТИЧНОГО ФОРМОУТВОРЕННЯ
Лекція __. РІСТ І РОЗВИТОК РОСЛИН
РІСТ ЯК ПРОЦЕС НОВОУТВОРЕННЯ ЕЛЕМЕНТІВ СТРУКТУРИ РОСЛИН
Ріст рослин — збільшення розмірів її маси, новоутворення елементів структури на основі розмноження, розтягу і диференціації клітин.
Рослина протягом життя може інтенсивно рости — утворювати велику надземну і підземну біомасу — і може не рости, тобто перебувати в спокої. Час, коли рослина не росте, називають періодом спокою. Період спокою спостерігають у зиготи, яка починає ділитися лише через деякий час після запліднення. Зародок, що сформувався, також проходить період спокою. Тому свіжозібране насіння злаків, бобових та інших культур деякий час не проростає навіть при сприятливих умовах. Це явище називають післязбиральним дозріванням. Тривалість післязбирального дозрівання залежить від погодних умов. У дощову і холодну осінь післязбиральне дозрівання відбувається повільніше, ніж у теплу і суху пору року. Нездатність насіння до проростання в період післязбирального дозрівання пояснюють наявністю непроникної для води і повітря оболонки і фізіологічною незрілістю зародка.
Для того щоб підвищити схожість насіння, що перебуває в спокої, вдаються до скарифікації й стратифікації його.
Скарифікація — видалення або руйнування оболонки насіння. Для цього насіння перетирають з піском, нагрівають до 60° С, проморожують, занурюють в окріп, обливають сірчаною кислотою. У грунті оболонка насіння руйнується під дією фізичних, хімічних і біологічних факторів. Перед висіванням скарифікують насіння конюшини, люцерни, білої акації, гледичії, дуба, плодових культур.
|
|
|
Стратифікація — припинення стану спокою у насіння під дією холоду. Для цього насіння тримають у вологому середовищі при температурі 5 °С протягом осені і зими. Цей прийом використовують для насіння багатьох деревних порід, кормових і квіткових культур.
Під впливом несприятливих умов середовища рослини припиняють ріст, впадають у вимушений спокій. Так, у пустинях Середньої Азії багато рослин через нестачу вологи і високу температуру повітря припиняють ріст влітку і відновлюють його восени.
Періодичне припинення росту спостерігають у паростків деревних рослин. З наближенням осені паросток припиняє ріст, на його верхівці формується брунька. Це явище називають впаданням рослини в глибокий, або органічний, спокій. Якщо гілочку бузку, зрізану у жовтні, поставити в кімнаті у воду, то вона не розпустить квітки, а в грудні не тільки розкриються квітки, а й почнеться ріст пагона. Отже, пагони бузку набули органічного стану спокою, а взимку вони перебувають вже у вимушеному спокої і тому, потрапивши в тепло, починають рости.
|
|
|
Перехід рослин до стану спокою супроводиться утворенням органів розмноження, що перебувають у спокої,— спор, насіння, бруньок, цибулин і бульб, в яких є зародки або зачатки пагона, що містять поживні запаси і захисні покриви від несприятливих умов зовнішнього середовища.
Різновидом спокою можна вважати циклічний ріст пагонів у цитрусових та у деяких плодових на півдні. На Західному Кавказі у цитрусових ріст пагона припиняється навесні, влітку і восени. Вторинний ріст пагонів на півдні спостерігають у абрикоса, вишні, сливи, дуба тільки в теплу погоду. Це явище відбиває ритмічні коливання інтенсивності важливих факторів росту — температури, тривалості дня, вологості та ін.
У рослин помірного клімату ритмічність росту виявляється різкіше, ніж у тропічного. Спокій у тропічних рослин або нетривалий (1-2 тижні), або його зовсім немає. Періодичність росту пов’язана із сезонами дощів і засухи. Тривалий глибокий спокій характерний для бука, клена гостролистого, дуба зимового і нетривалий для бузку, жимолості, барбарису, тополі та ін. Серед кісточкових порід у вишні спокій триваліший, ніж у абрикоса і сливи. У літніх сортів яблуні спокій триваліший, ніж у зимових, а у південних сортів яблуні і груші спокій глибший, ніж у північних. Тривалість органічного спокою залежить і від зовнішніх умов. Волога у вигляді полинів улітку і дощів восени, а також азотні добрива затримують настання спокою. Засуха, літня спека і фосфорно-калійні добрива прискорюють набування рослинами спокою. Молоді рослини впадають у стан спокою пізніше, ніж старі. Коропі взагалі не мають органічного спокою і лише припиняють ріст через несприятливі умови зовнішнього середовища.
|
|
|
Отже, органічний спокій — пристосувальна властивість рослин, яка виникла в ході еволюції під виливом умов зовнішнього середовища і перетворилася на спадкову.
Гальмування росту під час спокою пояснюють нагромадженням інгібіторів росту, які виявлено у більшості рослин. Вони містяться в м’якоті плодів яблуні, груші, у соку плодів помідорів, жимолості, в оболонці насіння каштана, латуку, пшениці, у зародках соняшника, в ендоспермі насіння півників, у цибулинах цибулі й часнику, в коренях моркви, редиски. Інгібітори росту неспецифічні для рослин. Ними можуть бути ціаніди, аміак, етилен, гірчичні олії, органічні кислоти, ненасичені лактони, альдегіди, ефірні олії, алкалоїди, дубильні речовини, антибіотики та іп. Осіннє накопичення інгібіторів змінюється зимовим поступовим руйнуванням їх, і в разі сприятливих весняних умов рослина відновлює ріст.
|
|
|
Проте спокій у рослин має відносний характер, оскільки охоплює не всі тканини рослинного організму. В пагонах дерев під час спокою крохмаль перетворюється на сахар і утворюються ліпіди. У насіння злаків під час післязбирального дозрівання спостерігають ферментативне перетворення речовин, внаслідок якого поліпшуються хлібопекарські якості борошна.
Спокій має велике народногосподарське значення, оскільки запобігає проростанню зерна у дощову погоду. Крім того, збільшується тривалість зберігання бульб і цибулин, а квіткові бруньки у плодових і декоративних рослин не розвиваються передчасно. Проте спокій часто і заважає господарському використанню рослин. Тривале післязбиральне дозрівання насіння пшениці в умовах Сибіру і районів Півночі знижує схожість посівного матеріалу. Спокій бульб картоплі заважає використовувати свіжозібрані бульби для літнього вирощування другого врожаю. Те, що квіткові бруньки не проростають восени і на початку зими, скорочує асортимент квітів у зимову пору року. В зв’язку з цим було винайдено способи керування спокоєм рослин.
Для розпукування квіткових бруньок широко використовується спосіб теплових ванн (рис. 23). При температурі води 30-350 С гілки занурюють у ванну на 9-12 год, а при температурі 42° С — тільки на 1-2 год. Стан спокою припиняють, обробляючи рослини хімічними сполуками: ацетиленом, етиленхлоргідрином, тіосечовиною, тіаміном
|
|

та нікотиновою кислотою (вітамінами В) і РР). Для того щоб подовжити період спокою, бульби картоплі обпилюють дустом, який містить 1,7-3,5 % метилового ефіру α-нафтилоцтової кислоти. Найбільш ефективним є спосіб опромінювання бульб картоплі γ-променями радіоактивного кобальту. Розпукування бруньок плодових гальмується від обприскування дерев влітку попереднього року розчинами а-нафтилоцтової і гідрозидмалеїнової кислот.
Після закінчення органічного спокою насіння здатне до проростання. Проте схожість його коливається у великих межах. Насіння бур’янів зберігає схожість протягом десятиліть, а насіння культурних рослин — нетривалий час: насіння пшениці, ячменю і вівса протягом 10 років; жита і кукурудзи — 5 років; овочевих культур — 2-3 роки; капусти і помідорів —до 5 років, а гарбузових — понад 5 років. Швидко втрачає схожість насіння проса.
Насіння дуба, бука, верби та ін. втрачає схожість протягом 2-3 тижнів.
Нормальне проростання насіння зумовлюється наявністю тепла, води і кисню. Сухе насіння містить тільки зв’язану воду і для проростання має набубнявіти, тобто поглинути певну кількість води, необхідну для активації ферментів і створення середовища для біохімічних реакцій. Напочатку насіння бубнявіє завдяки силам гідратації, а наприкінці — осмотичним енлам завдяки нагромадженню продуктів гідролізу білків, ліпідів і вуглеводів. Кількість води, яку поглинуло насіння при набубнявінні, залежить від хімічного складу зародка, ендосперму і покривів насіння. Насіння бобових, багате на білки, поглинає більше води, ніж крохмалисте насіння злаків. Дуже багато води поглинає насіння льону і цукрових буряків, оболонки яких містять слизуваті гідрофільні полісахари (табл. 6).
6. Кількість води, яку поглинає насіння при набубнявінні
(в % до повітряносухої маси) (Гребинський, 1961)
| Насіння | Кількість води | Насіння | Кількість води |
| Пшениця | 47,7 | Сочевиця | 93,3 |
| Жито | 64,7 | Вика | 75,4 |
| Овес | 76,3 | Горох, боби, квасоля | 114,0 |
| Ячмінь | 517,4 | Льон | 160,6 |
| Просо | 38,2 | Буряки кормові | 65,2 |
| Кукурудза | 44,0 | « цукрові | 168,0 |
| Коноплі | 43,9 | Конюшина лучна | 143,0 |
| Соняшник | 56,5 | Тимофіївка | 80,0 |
| Рапс | 51,0 | Тонконіг | 90,0 |
| Люцерна | 56,0 | Мак | 91,0 |
Швидкість поглинання води при набубнявінні у різного насіння неоднакова. Так, насіння гірчиці, редиски, капусти поглинає воду протягом кількох годин, а люпину — кількох днів.
Насіння білої акації може лежати у воді 30 років, перш ніж почне проростати. Якщо набубнявіле насіння добре провітрюється, то воно швидше поглинає воду, ніж повністю занурене в неї.
Це свідчить про тісний зв’язок процесів поглинання води і дихання.
Насіння проростає при температурі вищій за 0° С. Мінімальна температура для проростання насіння жита, пшениці, ячменю, вівса, вики, сочевиці, гороху, горошку, моркви, брукви, рижію, конопель, гірчиці 1-2° С, кукурудзи, проса, сої — 8-10° С, квасолі, рицини і сорго 10-120 С, рису, огірка, гарбуза, кавуна 12-14° С. Для польових умов середньодобова температура має бути на 2-3° С вищою. При цьому з’являються дружні сходи культури.
Крім води і температури для проростання насіння потрібний кисень, споживання якого рідко посилюється з початком росту зародка. Дрібному насінню, що має більшу питому поверхню, ніж велике, менше загрожує нестача кисню у спокійній воді. Навіть насіння водяних рослин не проростає у воді, бідній на кисень. Проростання насіння дуже гальмується, якщо вміст вуглекислоти близько 17 % і більше, а при концентрації її близько 35 % насіння гине.
Насіння культурних рослин однаково добре проростає як у темряві, так і на світлі. Насіння дикорослих рослин краще проростає одне на світлі, а інше в темряві. Світло потрібне для проростання злакових трав, тютюну, салату, селери, а темрява — для насіння бука, дуба, каштана, обліпихи.
Проростання насіння починається з появи корінця, потім витягується гіпокотиль, розкриваються сім’ядолі, останньою починає рости брунечка, з якої формується пагін. Проросток складається з пагона або облистяного стебла і кореня, який росте швидше, ніж пагін.
Ріст кореня. В частині кореня, що росте, є чотири зони. На кінчику кореня міститься ембріональна зона, вкрита кореневим чохликом. За нею йде зона розтягу, далі — зона кореневих волосків і зона галуження кореня. У кукурудзи зона росту кореня становить 4 мм. В першому міліметрі від кінчика кореня відбувається тільки поділ клітин, у другому міліметрі клітини збільшуються в об’ємі. В останніх двох міліметрах містяться зони розтягу клітин і галуження. Такий самий розподіл зон спостерігають у інших культурних рослин (горох, овес, пшениця, ячмінь).
Кожна клітина в процесі росту закономірно проходить окремі фази, що відрізняються певним обміном речовин і структурою. Спочатку клітина перебуває в ембріональній фазі, потім у фазі розтягу, і завершується ріст фазою диференціювання.
В ембріональній фазі клітини заповнені протоплазмою без видимої у мікроскоп вакуолі, у них збільшується ядро, і клітини всі діляться на дочірні мітозом або каріокінезом. Ембріональна фаза росту клітини відбувається в конусах наростання, у твірній меристемі.
У фазі розтягу клітини меристеми перестають ділитися, проте збільшуються в об’ємі, з’являються вакуолі з клітинним соком — спочатку багато дрібних вакуолей, потім вони зливаються в одну. Оболонки клітини збільшуються завдяки новим молекулам целюлози, проте помітного потовщення немає. Дуже збільшується кількість цитоплазми.
Фаза диференціювання клітини настає після досягнення нею певного розміру. Прн цьому вона може перетворюватися на спеціалізовану клітину: палісадну, паренхімну, ситоподібну трубку, членик судини, луб’яне волокно, флоему, тобто в будь-яку клітину, потрібну рослинному організмові.
З ростом клітини вміст води, сахаріи і білка збільшується. На кінець росту вміст білка зменшуються внаслідок диференціації клітини. Інтенсивність дихання н різних зонах ростучої частини кореня змінюється мало, збільшуючись у зоні розтягу. Дихальний коефіцієнт коливається близько одиниці внаслідок дихання завдяки вуглеводам.
У ростучому корені квасолі вміст ДНК. становив в ембріональній зоні 0,78 % сухої маси, в зоні розтягу — 0,42 і в зоні кореневих волосків — 0,34%, а вміст РНК відповідно 7,78; 2,83 і 2,06 %. У коренів сочевиці активність ауксиноксидази збільшується від кінчика кореня до зони галуження, активність пероксидази — навпаки. Аналогічне явище спостерігають у коріння щодо таких ферментів, як поліфенолоксидаза, цитохромоксидаза, дегідрогеназа і дезоксирибонуклеаза. У зоні розтягу кореня цибулі міститься також максимальна кількість вітамінів; аневрину, рибофлавіну і нікотинової кислоти.
Ріст кореня залежить від віку рослини. У пшениці у фазі двох листків коріння проникає у грунт на глибину 45 см, у вівса — 80 см. У ярої пшениці у фазі кущення загальна поверхня коріння становить 9,6 м2, під час трубкування — 29,4, на початку цвітіння — 36,7, наприкінці цвітіння — 44,1, а під час воскової стиглості зерна 30,9 м2. Коріння дерев росте повільніше. У яблуні і молодої сосни на піщаному грунті приріст за добу становив 0,3 см. Велике значення для росту коріння має вологість, аерація грунту і температура (табл. 7).
7. Оптимальні температури росту коренів
| Рослина | Температура, °С | Рослина | Температура, °С |
| Тонконіг луговий, гря | Кукурудза | 24 | |
| стиця збірна, типчак | Соя, бавовник | 27 | |
| лучний, кострець | 25 | Тютюн | 28 |
| Тимофіївка, мітлиця | 21 | Квасоля | 26 |
| Конюшина червона | 21 | Картопля | 20 |
| Суданська трава | 29 | Цибуля | 22 |
| Пшениця | 16 | Помідори | 20 |
| Ячмінь | 20 | Яблуня, персики | 18 |
З віком чутливість коренів до температури змінюється. У молодих рослин помідорів корені краще ростуть при 30° С, ніж при 20° С, а у дорослих — навпаки. При 10° С ріст коренів пригнічується на всіх фазах розвитку рослин.
Добрий ріст коренів спостерігають на грунтах з достатнім ступенем аерації. Серед культурних рослин мінімальна потреба в кисні у рису і гречки, а найбільша у помідорів, гороху і кукурудзи. Коріння рису та інших затоплюваних культур поглинає кисень із спеціальних повітропровідних тканин. Вміст кисню в повітропровідних порожнинах і болотяних рослин досягає 15 %. Потреба коренів у кисні тим більша, чим вища температура грунту. Критичний мінімум кисню для зберігання коренів живими близько 3 %. Певною мірою листки трав’янистих рослин можуть постачати кореням кисень навіть без спеціальних повітропровідних тканин. Так, якщо рослини озимої пшениці або жита затоплені талими водами, але кінчики листочків знаходяться у повітрі, то рослини від вимокання гинуть значно менше, ніж при повному зануренні листків у воду.
Корені потребують пухкого і вологого грунту. При розпушуванні грунту на 35-40 см основна маса коренів пшениці розміщується в горизонті 10-27 см, а при оранці на 20-22 см — в горизонті 0-10 см. В разі висихання грунту до вологості в’янення корені не ростуть. При зрошенні помірними дозами корені пшениці розміщуються у верхніх шарах грунту, а без поливання проникають глибоко в грунт.
Корені як гетеротрофні органи рослини потребують для росту органічних речовин, що надходять з листя. Тому умови середовища, сприятливі для фотосинтезу, сприяють росту коренів і навпаки. Підвищена температура в нічні години, яка спричинює посилену витрату вуглеводів на дихання, призводить до зменшення маси коренів. Скошування надземної частини рослин не тільки гальмує ріст коренів, а й призводить до зменшення їхньої маси. Багатий урожай плодів затримує ріст коріння дерева, а видалення суцвіття стимулює його.
Вода і іони солей інтенсивно поглинаються ростучою частиною кореня, зокрема зоною кореневих волосків. Тому підживлення рослин найефективніше тоді, коли поживні речовини, які вносять у грунт, вступають у контакт з кореневими волосками, чому сприяє багате галуження коренів. У плодових дерев маса коріння становить 25-30 % маси дерева, причому галуження коренів набагато більше, ніж крони. У однорічного сіянця яблуні налічують близько 45 000 коренів. Загальна довжина коренів і кореневих волосків у озимого жита становить 600 км, а довжина надземної частини пагонів і листя ледве досягає 20 м. Іони, які поглинаються з грунту, специфічно впливають на ріст коріння. Дуже стимулюють ріст калі.цій, молібден та ін.
Корені за своєю функцією є пе тільки провідним і опорним органом рослини, а й місцем біосинтезу необхідних для рослини фізіологічно активних сполук. У коренях біосинтезуються 22 амінокислоти, глутатіоп, уреїд та різні фосфорорганічні сполуки. Проте для нормального росту корінь має поглинати вітаміни, що надходять до нього з тканин інших органів. Для коренів гороху, люцерни і редиски необхідні тіамін та нікотинова кислота (вітаміни В1 і РР), а для моркви, бавовнику, конюшини, помідорів, соняшнику і дурману, крім зазначених, потрібний ще піридоксин (вітамін В6).
Ріст стебла. При проростанні насіння стебло формується з гіпокотиля. Стебло виконує опорну, провідну функцію і може бути місцем відкладання запасних речовин, молоді зелені стебла здійснюють фотосинтез. За своєю будовою і функціями до стебла близький пагін, що виростає з бруньки, яка утворюється в пазусі листка. Завдяки утворенню бічних пагонів стебло гілкується.
Ріст стебла у висоту зумовлений діяльністю верхівкової меристеми, а ріст у товщину — латеральними меристемами. У злаків стебло росте завдяки інтеркалярній меристемі, розміщеній біля основи міжвузля. Внаслідок поділу меристематичних клітин виникають клітини, які не діляться і дають початок окремим тканинам. У межах конуса наростання (верхівкова меристема) розрізняють протодерму, з клітин якої утворюється епідерміс, прокамбій, який формує провідну систему і основну меристему, що утворює основну тканину — паренхіму. Поділ клітин конуса наростання зумовлений біосинтезом ДНК і ауксинів — стимуляторів росту — диференціації клітин. Звичайно стебло росте верхівкою, тому нові вузли на ньому формуються згори доти, поки верхівка стебла не перетвориться на суцвіття. У злаків диференціація колоса відбувається дуже рано, у фазі кущення, і при видовженні стебла нові вузли не формуються. У пагонів, що ростуть із бруньок відновлення, вузли сформовані в бруньці. Якщо квітка формується на пагоні після виходу з бруньки, спостерігається формування нових вузлів. Міжвузля подовжуються в основному завдяки подовженню клітин і тільки у злаків завдяки утворенню нових клітин.
Ріст стебла в товщину здійснюється камбієм, розміщеним між флоемою лубу і ксилемою деревини. Витягнуті клітини камбію діляться вздовж довгої осі і в напрямі до периферії стебла з наступною диференціацією дочірніх клітин або к елементи ксилеми, або — флоеми. В однорічних рослин клітини камбію припиняють ділитися до початку цвітіння, у дерев і кущів камбій переходить до стану спокою із середини осені і до весни.
Настання спокою у камбію спочатку виявляється в гілках, потім у стовбурі і в останню чергу — у коренях. Періодичність діяльності камбію спричинює утворення річних кілець у стовбурі дерева. Добре виражені річні кільця у дуба і слабо у верби і тополі. Весняне пробудження камбію пов’язане з дією ростових гормонів, що опускаються з розпуклих бруньок, в яких до весни різко збільшується вміст ауксинів. Після того як клітини камбію починають ділитися, в них синтезуються ауксини та інші активатори росту, які підтримують активність камбію незалежно від ауксинів листя.
Отже, ріст стебла в довжину і товщину відбувається за типом S-подібної кривої, тобто спочатку повільно, потім інтенсивно, загасаючи наприкінці росту. Внаслідок такого характеру росту довжина міжвузлів неоднакова. Найдовші міжвузля бувають звичайно в середній частині пагона. Виняток становлять міжвузля злаків, які збільшуються знизу догори.
У стебла добре виражені відмінності властивостей у верхній і нижній частинах його. Це явище називають полярністю. Ростуче стебло неоднорідне за довжиною: основа стебла складається з тканини, клітини яких перестали рости, а вершина утворена ростучими клітинами і тканинами. Це зумовлює відмінність морфологічних і фізіологічних властивостей різних частин стебла. Полярність виявляється в закономірному зростанні або зменшенні концентрації речовин, інтенсивності фізіологічних процесів, що, в свою чергу, виявляється в ярусності листя, утворенні біля основи пагона коренів, а на верхівці бруньки і взаємозалежності різних органів рослини.
Поки листок залишається життєдіяльним, він гальмує ріст своєї пазушної бруньки. Тому знищення листя або пошкодження його стимулює швидкий ріст бруньок і появу нового листя. Верхівка пагона гальмує ріст бічних бруньок, що залишилися після опадання листя. Знищення або пошкодження її стимулює проростання сплячих бруньок, а отже, й галуження. Гальмування росту бічних бруньок може бути різним. У дерев воно виявляється більше, тому дерева гілкуються менше, ніж кущі, а в помідорів і тютюну гальмування послаблене, і тому у них з пазушних бруньок часто ростуть нові нагони. Гальмування росту бічних бруньок непостійне в онтогенезі. Воно послаблюється з віком, і бічні пагони у яблуні, що добро розрослися, починають пригнічувати ріст головного пагона. У ялини знищення верхівки приводить до того, що найближча До верхівки гілка вигинається догори, ніби замінюючи верхню бруньку, і активно гальмує ріст бічних пагонів.
Ріст стебла безпосередньо залежить від зовнішніх умов. Так, температура значною мірою визначаю довжину пагона. Для рослин помірного клімату оптимальна температура буває 25-30° С, причому інтенсивний ріст рослин спостерігають тільки при коливанні температури протягом доби і різниці температури повітря і ґрунту.
Для нормального росту льону, гречки і пшениці при температурі повітря 20 °С температура ґрунту має бути 10 °С.
Значною мірою висоту рослини визначають ґрунтове живлення і водопостачання.
Затримання росту в разі засухи зумовлене втратою тургору клітинами, зменшенням вмісту ауксинів і нуклеїнових кислот.
Ріст листя. Листок розвивається на конусі наростання пагона і являє собою групу меристематичних клітин, які диференціюються в зародковий листок. Диференціація зародкового листка відбувається від основи до верхівки. З верхівки утворюється листкова пластинка і черешок, а з нижньої — піхва листка і основа черешка в місці прикріплення його до стебла. Формування листкової пластинки в брунці може починатися від основи до верхівки, від верхівки до основи, від середини листка вгору і донизу або рівномірно вздовж всієї пластинки. Для більшості листкових пластинок характерною є наявність кількох точок росту, внаслідок чого на них утворюються зубці, лопаті, листочки. У дводольних ріст листкової пластинки здійснюється рівномірним ростом вздовж всієї площі листка. Цей ріст зумовлений розтягом клітин і тільки почасти їхнім поділом. У однодольних листкова пластинка подовжується завдяки утворенню нових клітин в основі листка. Ріст пластинки в товщину здійснюється розтягом клітин палісадної тканини і поділом клітин лизофілу.
У листкових зародках багато ауксинів, які впливають на морфогенез листка. Підвищені дози стимуляторів росту спричинюють різні зміни в будові листка (ненормальне жилкування, зростання листочків у перистих листків та ін.). Аналогічно діють гербіциди. З обміном пов’язаний листопад. Так, утворення окремого шару спричинене зменшенням притоку ауксинів з листкової пластинки в черешок. При цьому перед опаданням листя в ньому порушується співвідношення етилену і ауксинів.
Будова листкової пластинки визначається тривалістю освітлення. Світло стимулює поділ клітин, проте гальмує їхній розтяг. Тому у затінку листки виростуть більшими і тоншими, ніж на світлі.
Знижена температура гальмує ріст листка в довжину, проте стимулює ріст його в товщину. При цьому у морозостійких сортів пшениці фаза розтягу клітин послаблюється більше, ніж у нестійких.
Ріст листя залежить від вмісту елементів живлення В: рослині. На цьому ґрунтується листкова діагностика, яка дає змогу за хімічним аналізом листка прогнозувати врожай, своєчасно вносити підживлення тощо. Так, наявність кобальту стимулює ріст листя, відсутність молібдену гальмує розростання листкової пластинки в обидва боки від центральної жилки.
В зв’язку з особливістю росту листя у рослин встановлено чітко виражені градієнти вмісту води, різних хімічних сполук, активності ферментів, інтенсивності транспірації, фотосинтезу, дихання в листках, розміщених на різній висоті по стеблу. Такій самій закономірності підлягають і білки, яких у верхніх листках завжди більше, ніж у нижніх.
Старіння листка супроводжується припиненням росту його, в основі цього явища лежить зниження вмісту білка і нагромадження не використаних на інші біосинтези вуглеводів (сахарів, крохмалю, інуліну, клітковини), інгібірування ферментів і біосинтез речовин вторинного походження (алкалоїдів, ліпідів, терпенів, глікозидів, пігментів), послаблення фотосинтезу, транспірація, збільшення вмісту вільної води і зменшення колоїдно-зв’язної.
ТРОПІЗМИ, НАСТИ, НУТАЦІЇ
Завдяки росту рослини здатні рухатися в просторі і часі. Ці рухи надзвичайно різноманітні і мають велике значення в пристосуванні рослин до умов середовища. Багато які з слабкоздерев’янілих, похилоростучих пагонів просуваються вперед завдяки приросту в довжину, періодично утворюючи корені і на нових ділянках стебла (гарбуз, ожина). У деяких рослин верхівки стебел знаходять опору, здійснюючи колові рухи: листки і стебла повертаються до світла; корені рухаються у грунті в напрямі до вологи і поживних речовин. Особливості ростових рухів зумовлюють поділ їх на тропізми, настії й нутації.
Тропізми — вигини органів рослин під впливом факторів, що діють однобічно. В разі однобічного освітлення виявляється фототропізм — вигини органів внаслідок різної інтенсивності росту освітленого і тіньового боків (рис. 24), геотропізм — вигин органів під дією сили тяжіння Землі. Якщо покласти рослину горизонтально, то через деякий час стебло вигинається вгору, а корені — донизу, тобто для стебла властивий негативний геотропізм, а для коренів — позитивний (рис. 25, 26). Під дією сили тяжіння значно змінюється розподіл осмотичного тиску і сисної сили в клітинах горизонтально орієнтованого органа з різних боків. Накопичення осмотично активних речовин на нижньому боці збільшує сисну силу її клітин і призводить до стимуляції розтягу їх, внаслідок чого нижній бік стає випуклим і визначає утворення негативного геотропічного вигину. Крім перелічених тропізмів, у рослин спостерігається хемотропізм (вигини від дії хімічних речовин), травмотропізм (вигини від уражень), термотропізм (вигини від дії температури), гідротропізм (рис. 27) (вигини від дії води) та ін.
|
|
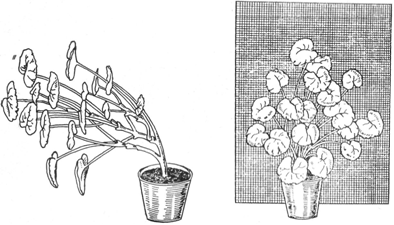
Рис. 24. Фототропізм листків і стебел.
Тропізми потрібні рослинам для орієнтації в просторі, кращого пристосування до умов зовнішнього середовища.
|
|

Рис. 25. Геотропічний вигін кореня бобів у вологому повітрі при спостереженні крізь скло:
А — вихідне горизонтальне положення; В — через 7 год; С — через 24 год. Трикутником позначено положення кінчика кореня.
|
|

Рис. 26. Підняття соломини злака завдяки утворенню геотропічних вигинів у вузлах.
|
|


|
|
Рис. 27. Гідротропізм коренів. Корені притискуються до тирси, замість того, щоб рости донизу.
В житті рослин найчастіше виявляється головний тропізм, що переважає над іншими. Якщо дерево росте на відкритому місці, то у стовбура спостерігають негативний геотропізм, тобто воно росте вертикально вгору. У нього не виявляються ні фототропізм, ні травмотропізм, оскільки немає сусідніх дерев, ні термотропізм, оскільки температура однакова навколо крони. Якщо грунтові води неглибокі і незасолені, то корені у дерева ростимуть вертикально вниз до води, тобто позитивний геотропізм і гідротропізм збігаються за напрямом росту. Проте якщо біля дерева висипати мінеральні добрива, то корені ростимуть горизонтально і в протилежний бік від них, тобто спостерігається негативний хемотропізм, що переважає над геотропізмом. І лише через деякий час, після сильних дощів, коли концентрація добрив стане мінімальною, короні дерева ростимуть до звичайного місця перебування їх, тобто у них виявиться позитивний хемотропізм.
Настії — рухи рослин, зумовлені змінами фізіологічного стану тканин, які по-різному діють па ріст органів в окремих частинах їх. Поширеними ппстичіпіми рухами є епінастії — вигини органів донизу і гіпонастії вигини органів догори. Настії зумовлені різною швидкістю розтягу клітин тканини, розміщених протилежно на органі. Від зміни дня і ночі бувають фотонастії, при яких квіти розкриваються і закриваються (кульбаба, нічна фіалка); зміна водопостачання спричинює гідронастії.
Крім ростових пастичиих рухів, у рослин спостерігають рухи, зумовлені створенням нерівномірного тургорного натягу у клітин різних боків органа. Такі рухи називають сейсмонастичними.
У мімози при механічній дії різко опускаються пагони і листочки, у бальзаміну від доторкання до плодів стулки негайно розкриваються і насіння висипається.
Нутації — періодично повторюваний ріст протилежних сторін органа. При цьому верхівка стебла або листковий вусик бобових здійснюють колові рухи для зіткнення з опорою, навколо якої закручуються рослими. Ці рухи характерні для кручених рослин.
Ростові рухи рослин визначають орієнтацію стебел і листя в просторі, зумовлюють форму пагона. Послаблення негативного геотропізму у стебел спричинює вилягання у злаків. Орієнтація пагона у просторі впливає на інтенсивність росту і формоутворювальні процеси. Прискорення дозрівання гарбуза, кавуна, огірка, дині пов’язане з пришпилюванням верхівки пагонів до землі, а у плодових — з переведенням гілок у горизонтальне положення.
Взаємозв’язок між орієнтацією пагонів й інтенсивністю їхнього росту характеризується законом Фехтінга: інтенсивність росту пагона прямо пропорційна висоті розташування його на материнській осі і обернено пропорційна нахилу до вертикалі. Доповненням до закону є вікові зміни листків, розміщених на різних ярусах. У пагонів, що ростуть похило, виявляється анізофілія, при якій інтенсивність росту більша у листків, що з’явилися на нижньому боці пагона, ніж у листків верхнього боку. У таких пагонів ізоелектрична точка (ІЕТ) білків зміщена в кислий бік.
Ростові рухи листя, квітів та інших органів рослин характеризуються добовою ритмічністю, яка визначається змінами фотоперіодичних умов. Основою періодичності рухів є ендогешішї ритм, тобто внутрішні фізіологічні зміни, для яких характерна певна розмірність, пристосована до добової зміни умов освітлення, акцептором чого є пігментна система листя, чутлива щодо червоного і віддаленого червоного світла.
Сучасні уявлення про рух живих організмів грунтуються на визначенні В. О. Енгельгарда: «єдність у безлічі». Єдність процесу полягає в підвищенні активності аденозинтрифосфатази, в перетворенні АТФ і скорочувальних білків. Множинність процесу характеризується різноманітними формами руху від плазмодія міксоміцетів рослин до висо- коспеціалізованої м’язової тканини.
ПОНЯТТЯ ПРО ІНДИВІДУАЛЬНИЙ РОЗВИТОК ЯК ПРОЦЕС ОНТОГЕНЕТИЧНОГО ФОРМОУТВОРЕННЯ
Живий і неживий світи планети Земля єдині за своєю матеріальністю. Матерія існує у вигляді різних конкретних систем, що перебувають у постійному розвитку. Органічний світ є однією з таких систем, яка зародилася в надрах геологічної системи і яку певною мірою вважають продуктом розвитку її. Тому органічний світ не може розвиватися без постійного зв’язку з матеріальними системами неживої природи, що оточують його. Розвиток органічного світу — це розгортання системи у просторі з послідовною зміною в часі якісних особливостей.
Система органічного світу у просторі обмежена зовнішньою природою, зовнішнім середовищем і зовнішніми умовами. Зовнішня природа — все те, що оточує органічний світ в цілому. Вона існує самостійно, незалежно від нього і розвивається своїм шляхом. Зовнішнє середовище — матеріальні системи, що впливають на органічний світ або змінюються під його дією. Зовнішні умови — елементи навколишнього середовища, які входять до складу органічної системи і забезпечують існування і розвиток її. Таке обмеження забезпечує органічному світові під час розвитку його вибіркове ставлення до елементів зовнішнього середовища, його аналітичну діяльність, зумовлену спадковістю, згідно з чим здійснюється специфічний обмін речовин. Загальний обмін речовин відрізняється від біологічного тим, що він здійснюється незалежно від життєвих потреб організмів і нерідко призводить до руйнування їх.
Еволюція — природно-історичний процес, ніякого плану, умисності, свідомості у нього ніколи не було і немає. Виникнення будь-яких нових структур і функцій, посилення і послаблення тих, що вже є, – все здійснюється в постійній взаємодії оранізмів із зовнішнім середовищем і між самими організмами.
Кожна рослина і тварина в будь яких відношеннях є досить недосконалим організмом і протягом свого життя зазнає якісних змін, характерних для окремих етапів життєвого циклу, або індивідуального розвитку (онтогенезу). Зміна якісних особливостей організму — процес, в якому запліднена яйцеклітина або вегетативний зародок внаслідок поділу клітин набуває форми, характерної для дорослого організму. Онтогенез організмів характеризується: морфологічними змінами, внаслідок яких утворюються тканини, органи і весь організм в цілому; фізіолого-біохімічними процесами, що відбуваються в клітинах, тканинах і органах організму; реалізацією генетичної інформації, закладеної в будові молекул ДНК; екологічною пристосованістю до умов зовнішнього середовища і еволюційною мінливістю, яка відбувається при переході від предків до потемків на різних етапах філогенезу.
У рослин розрізняють вегетативний розвиток — процес формування вегетативних органів і генеративний розвиток— перехід до цвітіння і наступного плодоношення. Більшості видів вищих рослин властиве статеве розмноження — а нижчим — безстатеве. У деяких груп рослин статевого процесу зовсім не буває. При безстатевому і вегетативному розмноженні успадковуються властивості тільки одного організму, що характеризуються віковим станом. Статеве розмноження має переваги щодо збереження виду: по-перше, поєднання спадкових властивостей материнського і батьківського організмів приводить до утворення нових ознак; по- друге, зростає пристосовуваність потомства до умов існування, збільшуються адаптивні властивості організму; по-третє, утворюється численне потомство; по-четверте, створюється можливість заселення потомством великої території і потрапляння насіння в нові умови, які сприяють збільшенню мінливості.
Хоч зовнішні середовище і умови впливають на індивідуальний розвиток організмів, організми розвиваються в основному за певною успадкованою програмою, закладеною в ядрі клітини, а точніш у молекулах ДНК, що містяться у хромосомах. Молекули ДНК як початкова ланка у біосинтезі білка визначають кількісну і якісну сторони його, а отже, й процеси формування органів і властивостей організмів. Тому онтогенез визначається послідовним утворенням у різних частинах організму конституційних і ферментних білків, специфічних для цих органів, — морфоструктур. Виникнення морфоструктур залежить від взаємодії ядра і цитоплазми, пов’язаної з іншими клітинами організму, що розвивається, і умовами зовнішнього середовища. Цей фактор послаблює або посилює взаємозв’язок між білками, гістонами і ДНК. Гістони, з’єднані з фосфатними групами ДІІК з обох боків спіралі, блокують певні ділянки молекули ДНК, тримаючи їх у недіяльному стані. В певний період розвитку організму взаємодія ядра і цитоплазми знімає гіс- тонну блокаду з ДНК, і, згідно з спадковою інформацією, організм від вегетативного розвитку переходить до генеративного з утворенням нової морфоструктури — квітки. Далі в онтогенезі організму з’являються і зникають інші морфоструктури, характерні для цього виду.
Онтогенез в еволюційному розвитку постійно змінюється, що можна простежити на прикладах онтогенезів нижчих і вищих рослин.
У одноклітинних, колоніальних та інших примітивних водоростей, грибів і подібних до них організмів онтогенез постає у вигляді різних морфоструктур — біонтів, які можуть існувати самостійно. Протягом року розвиток нижчих організмів обмежений періодом від проростання спори до утворення спори. Навесні, коли грунт вологий, спора проростає в слань, найбільш пристосовану для життя в цих умовах. Як тільки на грунті нагромаджується вода, в слані утворюються зооспори, які виходять з неї і ведуть незалежний спосіб життя. При підсиханні грунту зооспори проростають, утворюючи нову слань.
Під час літньої засухи слань перетворюється на цисту, здатну вигримувати будь-яке зневоднення. Такі взаємоперетворення біонтів у нижчих спостерігають до осені, і всі вони відбивають пристосовуваність організму до зовнішнього середовища. Паралельно з утворенням біонтів нижчі організми розвиваються і врешті-решт набувають здатності утворювати нові спори, тобто розмножуватися. З настанням осені у водоростей і грибів утворюються статеві органи — гаметангії з гаметами, що дають спочиваючі зиготи, пристосовані до зимових умов. Отже, у процесі індивідуального розвитку нижчого організму немає внутрішньо закріпленого, послідовно змінюваного порядку якісних змін.
У високоорганізованих водоростей (харових, бурих, червоних) і вищих рослин за умови правильного чергування безстатевого і статевого поколінь сформувався єдиний цикл онтогенезу. В еволюційному розвитку у цих організмів один з біонтів поступово домінував щодо інших морфоструктур.
Ця домінантність біонта збіглася з тривалістю періоду сприятливих умов для організму. У такого біонта утворювані морфоструктури не відокремлювались, а допомагали йому пережити несприятливі умови. Так, з часом у одного біонта утворились органи, що зумовили краще пристосування до зовнішніх умов і певний порядок якісних змін організму. Отже, еволюція онтогенезу відбувалася від водорості— до псилотових і папоротей — з трьома і врешті до насіннєвих рослин з одним біонтом. Проте еволюція онтогенезу триває. Вивчення С.С. Хохловим численних форм апоміксису стало основою для теорії появи нового еволюційно більш досконалого типу безстатевонасіинєвих рослин. Перехід до цього типу здійснюється через редукцію статевих органів.
ЕТАПИ ОНТОГЕНЕЗУ РОСЛИН
Розвиток органічного світу немислимий без розвитку неорганічних систем, вони єдині, як єдиний онтогенез індивідуума з внутрішніми і зовнішніми факторами, що зумовлюють якісні зміни організму, які ведуть його до розмноження. В роботах І. В. Мічуріна накреслена тенденція до розгляду життя рослини не як однорідного процесу, а як такого, що складається з внутрішньо обумовлених, якісно різних етапів, протягом яких рослина має різні властивості і реакцію на зовнішнє середовище.
За І. В. Мічуріним, онтогенез деревних складається з таких етапів: ембріонального — від першого поділу зиготи до проростання насіння; молодості — від проростання насіння до першого цвітіння; змужніння — від цвітіння до перших років плодоношення; дорослого стану — від початку плодоношення до інтенсивного росту і багатого плодоношення; старіння — від багатого плодоношення до послаблення росту, плодоношення і смерті. Отже, етапи онтогенезу характеризуються не тільки кількісними, а й якісними змінами організму. Якщо культурна рослина на початку онтогенезу має певну схожість із своїми дикими предками, то до стадії змужніння вона формується як культурний сорт. Особливості етапів онтогенезу виявляються у відмінності пагонів дерева за ярусами. Пагони, що йдуть від кореневої шийки, за фізіологічними особливостями наближаються до диких форм, а пагони, які утворюються далі, вище по стовбуру, мають властивості сорту.
Цю закономірність у розвитку плодових рослин використовують у роботах з селекції рослин.
Основними регуляторами ритмів росту і розвитку рослин є процеси, які здійснюються під дією кількісного сприйняття точкою росту тепла й світла. Відомо, що озимі зернові культури, які було висіяно восени, зимують у фазі кущення, а навесні колосяться і дають урожай зерна.
Досліди показали, що при весняній сівбі озимі не колосяться, оскільки не підлягали дії зниженої температури, якої вони зазнають в умовах зими. Це явище називають післядією знижених температур.
У багаторічних трав’янистих рослин таких, як багаторічне жито, ячмінь цибульний, грястиця збірна, типчак лучний та інші протягом онтогенезу зберігається властивість ози- мості, що забезпечує щорічну пристосовуваність їх до перенесення морозів і засухи. Нові вкорочені пагони мають високу стійкість як проти морозів, так і проти спеки, тому вони з’являються після відмирання пагонів, що відплодо- носили, і тим самим забезпечують рослину в цілому високою стійкістю проти несприятливих факторів середовища Проте дія знижених температур на рослину не є абсолютною, вона може замінюватися дією інших факторів, які також забезпечують нормальний розвиток організмові.
Застосування цих факторів має практичне значення для сільського господарства, квітництва, селекції. Вміщуючи насіння озимих культур у набубнявілому стані в умови з низькими температурами, можна мати 2-3 покоління на рік, що дуже важливо для прискорення селекції. Цей прийом називається яровізацією, оскільки озимі стають ніби ярими і можуть колоситися в літній період. Потребу коренеплодів і капусти першого року життя в зниженій температурі використовують при організації насінництва їх. Теплі ванни, ефіризація дають змогу мати квіти бузку і конвалії в зимовий період.
Наступним регулятором розвитку рослин є фотоперіодизм, або тривалість дня. Прикладом цього є листопад у дерев помірної зони восени і у тропічних рослин перед літньою посухою. Зміна тривалості дня активізує поділ клітин, І утворюється окремий шар біля основи черешка. Листопад може затримуватися додатковим освітленням, проте при цьому різко знижується морозостійкість рослин. Отже, реагуючи на світло (фотоперіодична реакція), рослина заздалегідь «готує» організм для перенесення несприятливого сезону року із збереженням нормального онтогенезу. Це особливо важливо враховувати при інтродукції й акліматизації рослин, а також створенні нових сортів, пристосованих до фотоперіоду певної географічної широти.
Довгі дні у весняно-літній період сприяють швидкому переходові довгоденних рослин :« вегетативного до генеративного стану і тим самим визначають скоростиглість сортів пшениці, жита, ячменю. У короткоденних рослин (соя, просо, сорго) в цих умовах розвиток затримується, накопичується біомаса, а врожай зерна утворюється в період скорочення дня, тобто у другій половині літа. Тому скоростиглі сорти проса висівають як післяжнивні культури, а редиску і редьку вирощують навесні і восени при короткому дні.
Отже, теоретичне і практичне значення яровизації і фотоперіодизму полягає в тому, що вони відбивають загальну істотну закономірність індивідуального розвитку рослини, зумовлену еволюцією.
ПрПри розробці проблеми розвитку висувались і припущення про значення для цвітіння речовин гормональної природи. М.X. Чайлахян висунув гормональну теорію розвитку, згідно з якою для переходу рослини до цвітіння їй необхідно накопичити достатню кількість гормона цвітіння — флоригену. Цвітіння рослини, що росла в несприятливих умовах, прискорювалось, якщо до неї прищеплювали пагін з рослини, яка цвіла або не цвіла, проте накопичила в листках флориген.
У рослини є фізіологічно зв’язана система: листок — точка росту. Листок сприймає фотоперіодичний стимул, внаслідок якого утворюється флоригеп, а в точці росту він реалізується у вигляді квітки.
Флориген рухається тільки по живих клітинах флоеми черешків листків і стебла. Хімічну природу флоригену поки що не з’ясовано. Великого значення М.X. Чайлахян надає гіберелінам — стимуляторам, що виявляють сильну дію на розвиток багатьох видів рослин, його праці з вивчення природи дії гіберелінів дали змогу зробити висновок, що вони набагато більше діють на цвітіння рослин довгого дня, ніж на розвиток кроткоденних форм. Тому М.X. Чайлахян припускає двокомпонентну будову флоригену: гіберелін і антезин. Перший потрібний для росту пагона, а другий – для формування квітки (рис. 28).
Ефективність гібереліну у прискоренні цвітіння виявляється у тих рослин, квітування яких затримується через гальмування стеблування, наприклад у злаків. Під дією гібереліну прискорюється не тільки ріст стебла у злаків, а й розвиток листків, що збільшує кількість їх, а отже, і акцептування фотоперіодичного стимулу для розвитку організму. Аналогічний вплив виявляє гіберелін на дворічні рослини, в яких па першому році життя за нормальних умов стебло залишається вкороченим і витягується тільки на друге літо після нерезимування. Оброблені гібереліном рослини зацвітають першого року. Крім гібереліну, аналогічну дію мають кінетин, що прискорює цвітіння перили черво- нолисткової при ізольованій культурі верхівок пагонів, і стероїди, які стимулюють цвітіння у потреби (короткоденна рослина) в умовах довгого дня. Встановлено наявність зв’язаних систем метаболічних факторів: співвідношення вуглеводів і азотних сполук, співвідношення активності метало- вмісних оксидаз (фенолази, цитохромоксидази, пероксидази, аскорбіноксидази) і оксидаз залишкового дихання (жовтий флавіновий фермент); співвідношення ауксинів і нуклеїнового обміну; утворення гібереліну — фактора, що стимулює формування стебла, і антезину — фактора формування квітки.
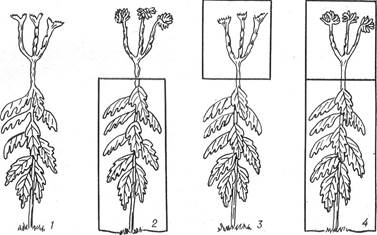
Рис. 28. Роль листків у фотоперіодичній реакції рослин (схема):
1 — рослину вирощено під час довгого дня; 2 — листки перебували на короткому дні, верхівки нагонів — на довгому; 3 — листки перебували на довгому дні, верхівки нагонів — на короткому; 4 — усі частини рослини вирощено під час короткого дня (у рамках — частини рослин, що перебували на короткому дні).
Зазначені метаболіти утворюються у всіх рослин, проте перехід до цвітіння здійснюється тільки при певному співвідношенні їх. У довгоденних рослин цвітіння відбувається при підвищеному накопиченні вуглеводів, посиленні активності металовмісних оксидаз, збільшенні вмісту ауксинів і накопиченні гіберелінів.
Для короткоденних рослин потрібні підвищений вміст азотистих сполук, висока активність оксидаз залишкового дихання, збільшення метаболітів нуклеїнового обміну і утворення антезинів.
Отже, внутрішній регулятор важко визнати однокомпонентною речовиною- «гормоном цвітіння». Швидше можна вважати його комплексом хімічних стимуляторів і інгібіторів, серед яких істотна роль належить компонентам нуклеїнового обміну.
Дата добавления: 2022-01-22; просмотров: 54; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!