ГЛАВА III . Основы популяционной генетики
В генетике человека особое значение имеет популяционный метод, который позволяет изучать гены и генотипы без постановки скрещиваний. В основе этого метода лежит закон, сформулированный в 1908 году независимо английским математиком Г. Харди и немецким врачом В. Вайнбергом (закон Харди-Вайнберга). Условия для выполнения этого закона следующие:
- популяция должна иметь неограниченный размер (быть достаточно многочисленной по меркам статистики);
- генотип по изучаемым генам не должен влиять на выбор брачного партнера (скрещивание должно быть свободным, то есть не ассортативным);
- миграция не должна существенно изменять генотип популяции;
- должен отсутствовать отбор по аллелям изучаемых генов.
В большинстве популяций человека для большинства признаков эти условия соблюдаются. Исключения, когда закон Харди-Вайнберга не может выполняться:
- островные, отдаленные и высокогорные популяции, где из-за небольшого числа особей случайные факторы могут повлиять на частоты аллелей;
- избирательность (ассортативность) связей, приводящих к рождению детей. Например, в США браки белых мужчин с белыми женщинами и черных мужчин с черными женщинами встречаются намного чаще, чем смешанные.
- иммиграция большого числа носителей редких в популяции генотипов;
- гены, аллели которых по-разному влияют на жизнеспособность и репродуктивную функцию.
Если частота в популяции доминантного аллеля А составляет p, то частота рецессивного аллеля а будет q = 1 - p. Согласно первому положению закона Харди-Вайнберга эти значения будут неизменны из поколения в поколение (при условии выполнения требований, изложенных выше) - это состояние генетического равновесия в популяции. Соотношение равновесных частот генотипов будет определяться возведением соотношения частот аллелей в квадрат - это второе положение закона. И третье положение закона Харди-Вайнберга говорит о том, что равновесие частот генотипов достигается за одно поколение и остается неизменным.
|
|
|
(p + q)2 = p2 + 2pq + q2
p - частота доминантного аллеля А
q - частота рецессивного аллеля а
p2 - частота генотипа АА (доминантные гомозиготы)
2pq - частота генотипа Аа (гетерозиготы)
q2 - частота генотипа аа (гомозиготных рецессивов)
Пример:
Одна из форм альбинизма (отсутствия пигментации кожи, радужной и пигментной оболочек глаза) у человека обусловлено редким рецессивным аллелем а (мутация в гене тирозиназы). В некоторой популяции частота альбиносов равна 0,0001. Тогда,
q - частота рецессивного аллеля а - 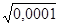 = 0,01
= 0,01
p - частота доминантного аллеля А - 1 - 0,01 = 0,99
p2 - частота генотипа АА (доминантные гомозиготы) - 0,992 = 0,98
|
|
|
2pq - частота генотипа Аа (гетерозиготы) - 2 х 0,99 х 0,01 = 0,02
Из примера видно, что гетерозигот по гену альбинизма в популяции в 200 раз больше чем альбиносов.
В случае множественного аллелизма используют аналогичные расчеты.
Пример:
В популяции индусов I группа крови встречается с частотой 0,314, II - 0,189, III - 0,410, IV - 0,087.
Пусть частота аллеля I0 - r, аллеля IA - p, аллеля IB - q.
Тогда, носителей генотипа I0 I0 (I группа) будет r2. Таким образом,
r2 = 0,314
r =  = 0,560
= 0,560
Общая частота аллелей IA и IB (p + q) = 1 - r = 1 - 0,560 = 0,440
Суммарная частота групп крови I и III равна (q + r)2. Таким образом,
(q + r) 2 = 0,314 + 0,410 = 0,724
(q + r) = 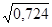 = 0,851
= 0,851
q = 0,851 - 0,560 = 0,291
p = 1 - q - r = 1 - 0,291 - 0,560 = 0,149
Итак, частоты аллелей групп крови системы ABO в популяции индусов следующие: I0 - 0,560, IA - 0,149, IB - 0,291.
В большинстве популяций наблюдается дрейф генов - изменение частот аллелей под влиянием случайных факторов. Эффект бутылочного горлышка - случайной гибели носителей того или иного генотипа при существенном снижении размера популяции - является наиболее частой причиной дрейфа генов. В небольших популяциях можно встретить эффект основателя - когда одна особь (почти всегда мужчина, например, Чингизхан) оставляет огромное число потомков, вследствие чего изменяется соотношение частот аллелей и генотипов.
|
|
|
Исходя из закона Харди-Вайнберга нетрудно убедиться, что отбор против гомозиготных рецессивов не эффективен - элиминация (устранение) q2 носителей генотипа аа не влияет существенно на частоты аллелей. Большинство носителей рецессивного аллеля являются гетерозиготами. В этом причина генетического груза в популяциях человека - значительного числа гетерозиготных носителей летальных (приводящих к смерти) аллелей и аллелей, связанных со снижением жизнеспособности и репродуктивной функции. Понятие генетического груза является фундаментальным в популяционной генетике, его ввел Г. Меллер в 1950 году в своей книге «Наш груз мутаций». Для расчета порядкового номера поколения (t), в котором начальная частота рецессивного аллеля (q0) примет ожидаемое значение qt при отборе против гомозиготных рецессивов, используют формулу:
t = 1/ qt - 1/ q0
Пример:
Частота рецессивного летального аллеля 0,01. Требуется установить, сколько потребуется поколений для ее уменьшения в 10 раз при условии отсутствия новых мутаций?
q0 = 0,01
qt = 0,001
t = 1/ 0,001 - 1/ 0,01 = 1000 - 100 = 900
Итак, для уменьшения частоты рецессивного летального аллеля с 0,01 до 0,001 потребуется целых 900 поколений.
|
|
|
Контрольные вопросы и задания к главе III
1. Рассчитайте частоты аллелей групп крови системы AB0 в популяции англичан, где I группа крови встречается с частотой 0,462, II - 0,436, III - 0,074, IV - 0,028.
2. Изменяется ли генетический груз в популяциях человека со временем? Если да, то благодаря действию каких факторов?
3. На одном острове дикари приносили в жертву всех альбиносов до достижения ими половозрелого возраста. Изначальная частота встречаемости носителей этого фенотипа была 0,0001. Насколько она изменилась через 180 поколений?
Дополнительная литература к главе III
Айала Ф., Кайгер Дж. Современная генетика. Т. 3 // М.: Мир. 1988. 332 С.
ГЛАВА IV . Близнецовый метод
В предыдущих главах речь шла о наследовании дискретных (качественных) признаков. Большинство морфологических (рост, телосложение, форма отдельных частей тела), физиологических (интенсивность обмена веществ, скорость роста) и психологических (темперамент) признаков являются количественными, их можно выражать числовыми значениями и сравнивать между собой. Одним из подходов генетического анализа количественных признаков у человека является близнецовый метод.
Следует различать два принципиальных типа близнецов - разнояйцовые (дизиготные), которые возникают при оплодотворении двух разных яйцеклеток двумя различными сперматозоидами, и однояйцовые (монозиготных), которые появляются в результате дробления уже оплодотворенной яйцеклетки (зиготы). Дизиготные близнецы ничем не отличаются от обычных сибсов. Монозиготные близнецы являются клонами - генетически идентичными организмами.
Известны случаи, когда дизиготные близнецы имели разных отцов. В этом случае с точки зрения генетики они являются полусибсами. Рождение однополых дизиготных близнецов случается несколько чаще, чем разнополых, но это связано, скорее всего с разной подвижностью сперматозоидов, несущих Х- и Y-хромосомы. Разнояйцовые близнецы появляются в результате одновременного созревания двух и более яйцеклеток - полиовуляции, которая обычна для многих млекопитающих, но у человека происходит с небольшой частотой. Склонность к полиовуляции наследуется (если у женщины уже были близнецы, то частота их рождения во второй раз в четыре раза выше, чем в среднем в популяции).
Монозиготные близнецы иногда разделены не полностью, у них могут быть даже общие органы. В таких случаях производят хирургическое разделение, зачастую сопряженное с риском для жизни обоих близнецов.
Средняя частота рождения монозиготных близнецов - около 40 на 10000 и мало варьирует в разных популяциях человека. Дизиготные близнецы чаще рождаются у представителей негроидной расы (около 100 на 10000), у европеоидов - около 80 на 10000, у монголоидов - около 20 на 10000. Этот факт можно объяснить наследственным характером склонности к полиовуляции и негенетическими причинами дробления оплодотворенной яйцеклетки.
Влияние на признак наследственности и факторов среды можно определить исходя из степени сходства (конкордантности) дизиготных (имеющих разный генотип, но выросших в близких условиях) и монозиготных (имеющих один генотип и выросших в близких условиях) близнецов. Особое значение имеет изучение разлученных в младенческом возрасте монозиготных близнецов (Рисунок IV, 1), так как они выросли в разных условиях, но являются генетически идентичными организмами. Существенным ограничением метода является то, что он позволяет установить только факт наследования изучаемого признака, но не дает возможности ответить на вопросы о типе наследования, количестве участвующих генов и их возможном взаимодействии.

Рисунок IV, 1 . Разлученные в раннем возрасте близнецы, выросшие в разных социальных условиях.
Контрольные вопросы и задания к главе IV
1. У отца вторая группа крови, у матери - первая. Родились близнецы - мальчик с первой группой крови и девочка - с четвертой. Может ли формальный отец считаться биологическим отцом этих детей?
2. Зависит ли вероятность рождения близнецов у супружеской пары от наличия или отсутствия близнецов среди кровных родственников жены и мужа?
Дополнительная литература к главе IV
Фогель Ф., Мотульски А. Генетика человека. Т. 1 // М: Мир. 1989. 308 С.
Дата добавления: 2021-07-19; просмотров: 157; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!