Системность и организованность жизни
Проявления жизни на Земле чрезвычайно многообразны. Жизнь на Земле представлена ядерными и доядерными, одно- и многоклеточными существами; многоклеточные, в свою очередь, представлены грибами, растениями и животными. Любое из этих царств объединяет разнообразные типы, классы, отряды, семейства, роды, виды, популяции и индивидуумы.
Во всем, казалось бы, бесконечном многообразии живого можно выделить несколько разных уровней организации живого: молекулярный, клеточный, тканевый, органный, онтогенетический, популяционный, видовой, биогеоценотический, биосферный. Перечисленные уровни выделены по удобству изучения. Если же попытаться выделить основные уровни, отражающие не столько уровни изучения, сколько уровни организации жизни на Земле, то основными критериями такого выделения должны быть признаны наличие специфических элементарных, дискретных структур и элементарных явлений. При этом подходе оказывается необходимым и достаточным выделять молекулярно-генетический, онтогенетический, популяционно-видовой и биогеоценотический уровни (Н.В. Тимофеев-Ресовский и др.).
Молекулярно-генетический уровень. При изучении этого уровня достигнута, видимо, наибольшая ясность в определении основных понятий, а также в выявлении элементарных структур и явлений. Развитие хромосомной теории наследственности, анализ мутационного процесса, изучение строения хромосом, фагов и вирусов вскрыли основные черты организации элементарных генетических структур и связанных с ними явлений. Известно, что основные структуры на этом уровне (коды наследственной информации, передаваемой от поколения к поколению) представляют собой ДНК, дифференцированную по длине на элементы кода – триплеты азотистых оснований, образующих гены.
|
|
|
Гены на этом уровне организации жизни представляют элементарные единицы. Основными элементарными явлениями, связанными с генами, можно считать их локальные структурные изменения (мутации) и передачу хранящейся в них информации внутриклеточным управляющим системам.
Конвариантная редупликация происходит по матричному принципу путем разрыва водородных связей двойной спирали ДНК с участием фермента ДНК-полимеразы (рис. 4.2). Затем каждая из нитей строит себе соответствующую нить, после чего новые нити комплементарно соединяются между собой (см. рис. 4.1). Пиримидиновые и пуриновые основания комплементарных нитей скрепляются водородными связями между собой ДНК-полимеразой. Этот процесс осуществляется очень быстро. Так, на самосборку ДНК кишечной палочки (Escherichia coli), состоящей примерно из 40 тыс. пар нуклеотидов, требуется всего 100 с. Генетическая информация переносится из ядра молекулами иРНК в цитоплазму к рибосомам и там участвует в синтезе белка. Белок, содержащий тысячи аминокислот, в живой клетке синтезируется за 5–6 мин, а у бактерий быстрее.
|
|
|

Рис 4.2. Схема биосинтеза белка (по Л.С. Спирину, 1986): 1 –синтез иРНК (транскрипции). 2–3 – иРНК проходит через поры ядерной мембраны к рибосомам (трансляции), 4 – аминокислоты, соединяясь в определенной последовательности, из РНК образуют фермент, 5–6 – образование белка
Основные управляющие системы как при конвариантной редупликации, так и при внутриклеточной передаче информации используют «матричный принцип», т.е. являются матрицами, рядом с которыми строятся соответствующие специфические макромолекулы. В настоящее время успешно дешифруется заложенный в структуре нуклеиновых кислот код, служащий матрицей при синтезе специфических белковых структур в клетках. Редупликация, основанная на матричном копировании, сохраняет не только генетическую норму, но и отклонения от нее, т.е. мутации (основа процесса эволюции). Достаточно точное знание молекулярно-генетического уровня – необходимая предпосылка для ясного понимания жизненных явлений, происходящих на всех остальных уровнях организации жизни.
|
|
|
Онтогенетический уровень – следующая, более комплексная ступень организации жизни на Земле. Прежде чем охарактеризовать явления, происходящие на этом уровне, надо определить понятие «особь». Жизнь всегда представлена в виде дискретных индивидуумов. Это в равной мере присуще микроорганизмам, растениям, грибам и животным, хотя в указанных царствах индивиды имеют различное морфологическое содержание. Так, одноклеточные состоят из ядра, цитоплазмы, множества органелл и мембран, макромолекул и т.д. Сложность индивидуума у многоклеточных во много раз выше, поскольку он образован из миллионов и миллиардов клеток. Но одноклеточная и многоклеточная особи обладают системной организацией и регуляцией и выступают как единое целое. Индивид (индивидуум, особь) – элементарная неделимая единица жизни на Земле. Разделить особь на части без потери «индивидуальности» невозможно. Конечно, в ряде случаев вопрос об определении границ индивида, особи не столь прост и самоочевиден. Например, не вполне ясно, можно ли считать особью отдельные организмы, составляющие колонии коралловых полипов, мшанок, сифонофор. Каждая из таких частей хотя и существует в виде отдельного самостоятельного индивидуума, но зависит от других частей; между отдельными «особями» колонии существует разделение функций и взаимосвязь (рис. 4.3).
|
|
|
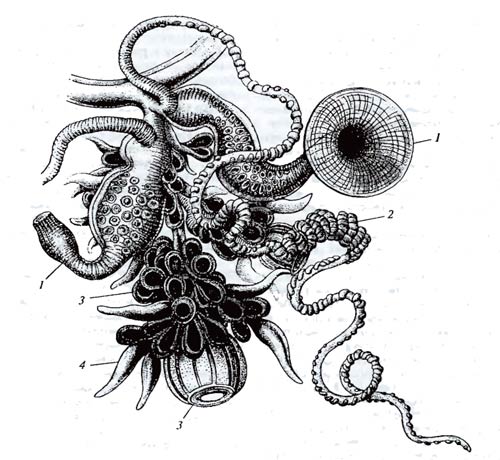
Рис. 4.3. Часть колонии сифонофоры Salacia (из Э. Геккеля, 1896). Единая на первый взгляд особь представляет собой колонию. Исходный индивидуум размножается почкованием; отпочковывающиеся особи могут вести самостоятельную жизнь или стать частями материнской колонии. 1 – питающие, 2 – стрекающие, 3 – половые, 4 – чувствующие группы особей
Не прост вопрос определения особи у таких комплексных организмов, как лишайники, являющиеся сложным симбиотическим сообществом водорослей, грибов и специфических микроорганизмов. Грибок Septobasidium в симбиозе с насекомым-червецом из семейства Coccidae дает новое симбиотическое образование – лакк, которое как единый организм введено в культуру человеком. Вопрос о границах индивида у растений, способных к вегетативному размножению, или у животных, размножающихся фрагментацией (например, гидра), не менее сложен. Тополиная роща, выросшая из черенков с одного родительского дерева, генетически одно и то же существо (пока не произойдет какая-либо соматическая мутация, делающая одно из новых растений генетически отличным от остальных). Во многих подобных неопределенных ситуациях приходится придавать понятию «особь» так называемое операциональное значение, указывая в каждом случае, к какой области биологии она относится. С эволюционной точки зрения особью следует считать все морфофизиологические единицы, происходящие от одной зиготы, гаметы, споры, почки и индивидуально подлежащие действию элементарных эволюционных факторов.
На онтогенетическом уровне единицей жизни служит особь с момента ее возникновения до смерти. По существу, онтогенез – это процесс развертывания, реализации наследственной информации, закодированной в управляющих структурах зародышевой клетки. На онтогенетическом уровне происходит не только реализация наследственной информации, но и апробация ее посредством проверки согласованности в реализации наследственных признаков и работы управляющих систем во времени и пространстве в пределах особи. Через оценку индивидуума в процессе естественного отбора происходит проверка жизнеспособности данного генотипа.
Онтогенез возник после дополнения конвариантной редупликации новыми этапами развития. В ходе эволюции возникает и постепенно усложняется путь от генотипа к фенотипу, от гена до признака. Как будет показано далее, возникновение онтогенетических дифференцировок лежит в основе возникновения всех эволюционных новообразований в развитии всякой группы организмов. В ряде экспериментальных эмбриологических работ установлены отдельные закономерности онтогенеза (см. гл. 14). Но все еще не создана общая теория онтогенеза. Мы до сих пор не знаем, почему в онтогенезе строго определенные процессы происходят в должное время и в должном месте. Пока можно предполагать, что элементарными структурами на онтогенетическом уровне организации жизни служат клетки, а элементарными явлениями – какие-то процессы, связанные с дифференцировкой. В общей форме ясно также, что онтогенез совершается вследствие работы саморегулирующейся иерархической системы, определяющей согласованную реализацию наследственных свойств и работу управляющих систем в пределах особи.
Популяционно-видовой уровень. Объединение особей в популяции, а популяций в виды по степени генетического и экологического единства приводит к появлению новых свойств и особенностей в живой природе, отличных от свойств молекулярно-генетического и онтогенетического уровней.
Популяция – элементарная структура на популяционно-видовом уровне, а элементарное явление на этом уровне – изменение генотипического состава популяции; элементарный материал на этом уровне – мутации.
Выделены элементарные факторы, действующие на этом уровне: мутационный процесс, популяционные волны, изоляция и естественный отбор (см. гл. 9–10). Каждый из этих факторов может оказать то или иное «давление», т.е. степень количественного воздействия на популяцию, и в зависимости от этого вызывать изменения в генотипическом составе популяции.
На популяционно-видовом уровне особую роль приобретают отношения между особями внутри популяции и вида. При этом популяции выступают как элементарные, далее не разложимые эволюционные единицы, представляющие собой генетически открытые системы (особи из разных популяций иногда скрещиваются, и популяции обмениваются генетической информацией). Виды, всегда выступающие как система популяций, являются наименьшими, в природных условиях генетически закрытыми системами (скрещивание особей разных видов в природе в подавляющем большинстве случаев не ведет к появлению плодовитого потомства). Все это приводит к тому, что популяции оказываются элементарными единицами, а виды – качественными этапами процесса эволюции. В целом же на популяционно-видовом уровне реально осуществляется в чреде поколений процесс эволюции.
Популяции и виды как надындивидуальные образования способны к существованию в течение длительного времени и к самостоятельному эволюционному развитию. Жизнь отдельной особи при этом находится в зависимости от процессов, протекающих в популяциях. Популяции и виды, несмотря на то, что состоят из множества особей, целостны. Но их целостность принципиально иная, чем целостность на молекулярно-генетическом и онтогенетическом уровнях. Целостность популяций и видов связана с взаимодействием особей в популяциях и поддерживается обменом генетического материала в процессе полового размножения (в отношении агамных и облигатко-партеногенетических форм этот вопрос требует дальнейшей разработки).
Популяции и виды всегда существуют в определенной среде, включающей как биотические, так и абиотические компоненты. Конкретная среда протекании процесса эволюции, идущего в отдельных популяциях,– биогеоценоз. В то же время биогеоценоз – элементарная единица следующего уровня организации жизни на Земле.
Биогеоценотический (экосистемный) уровень. Популяции разных видов всегда образуют в биосфере Земли сложные сообщества – биоценозы. Биоценоз – совокупность растений, животных, грибов и прокариот, населяющих участок суши или водоема и находящихся в определенных отношениях между собой. Вместе с конкретными участками земной поверхности (педосферы), занимаемыми биоценозами, и прилежащей атмосферой называются экосистемами.
Такие экологические системы (экосистемы) могут быть разного масштаба – от капли воды или муравьиной кучи до экосистемы острова, реки, континента и всей биосферы в целом. Экосистема – взаимообусловленный комплекс живых и косных компонентов, связанных между собой обменом веществ и энергией (А. Тенсли, 1935). Экосистема – «безразмерное» понятие, но есть один класс экосистем, имеющий определенные размеры и принципиальное значение как «кирпичики» организации всей биосферы,– биогеоценозы (В.Н. Сукачев, Н.В. Тимофеев-Ресовский). Биогеоценоз – это такая экосистема, внутри которой не проходит биоценотических, микроклиматических, почвенных и гидрологических границ; биогеоценоз – одна из наиболее сложных природных систем. Внешне заметные границы биогеоценозов чаще всего совпадают с границами растительных сообществ (фитоценозов).
Все группы экосистемы – продукт совместного исторического развития видов, различающихся по систематическому положению; виды при этом приспосабливаются друг к другу (подробнее см. гл. 19). Первичной основой для сложения биогеоценозов служат растения и прокариоты – продуценты органического вещества (автотрофы).
В ходе эволюции до заселения растениями и микроорганизмами определенного пространства биосферы не могло быть и речи о заселении его животными. Растения и прокариоты представляют жизненную среду для животных – гетеротрофов.
Биогеоценозы – среда для эволюции входящих в них популяций. Популяции разных видов в биогеоценозах воздействуют друг на друга по принципу прямой и обратной связи. В целом жизнь биогеоценоза регулируется в основном силами, действующими внутри самой системы, т.е. можно говорить о саморегуляции биогеоценоза. Автономность и саморегуляция биогеоценоза определяют его ключевое положение в биосфере нашей планеты как элементарной единицы на биогеоценотическом уровне.
Биогеоценозы, образующие в совокупности биосферу нашей планеты, взаимосвязаны круговоротом веществ и энергии. В этом круговороте жизнь на Земле выступает как ведущий компонент биосферы (В.И. Вернадский, В.Н. Сукачев). Биогеоценоз представляет собой незамкнутую систему, имеющую энергетические «входы» и «выходы», связывающие соседние биогеоценозы (рис. 4.4). Обмен веществ между соседними биогеоценозами может осуществляться в газообразной, жидкой и твердой фазах, а также в форме живого вещества (миграции расселения).
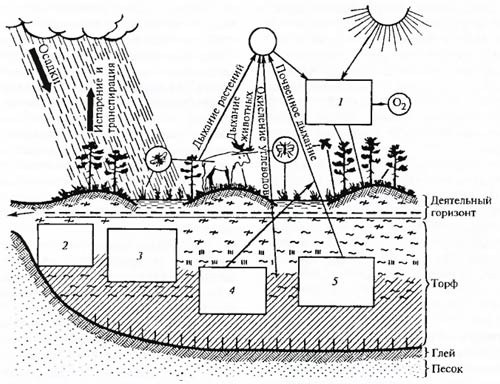
Рис. 4.4. Схема взаимоотношений компонентов в биогеоценозе участка верхового болота (по Н.И. Пьявченко. 1972): 1 – расход энергии (альбедо, транспирация, испарение, фотосинтез); 2 – подземная фитомасса 5 т/га; 3 – неорганические; вещества в метровом слое, т/га: азота – 2,8, магния – 0,6. фосфора – 0,7, калия – 0,8; 4 – в 1 г сухого торфа бактерий более 4 млрд экз., грибов более 175 тыс. экз.; 5 – на 1 м2 поверхности энхитренд, около 270 экз., коллембол около 4000 экз., акарин около 700 экз., нематод около 240 экз.
При эволюционном подходе проявляется тесная связь и взаимообусловленность всех уровней организации жизни. Хотя развитие представлений о системной организации жизни принято относить к 40-м годам XX в., основы их были заложены еще Ч. Дарвином. Именно он четко определял вид как систему, а естественный отбор – как фактор, упорядочивающий организацию жизни (А.А. Малиновский). Представления о биологических системах оказали мощное влияние и на развитие теории систем и кибернетики (К. Шеннон, У. Эшби), а последняя, в свою очередь, способствовала углублению представлений о системной организации жизни (И.И. Шмальгаузен).
14. Мутационный процесс – как элементарный фактор эволюции.
Как показано выше, мутации — элементарный эволюционный материал. Сам же процесс возникновения мутаций — постоянно действующий элементарный эволюционный фактор, оказывающий давление на популяции.
Значительная часть (от нескольких до нескольких десятков процентов) особей в популяции — носители вновь возникших мутаций. Постоянно идущий в природе мутационный процесс ведет к изменению в популяции частоты одного аллеля по отношению к другому. Хотя по каждому отдельному гену давление мутационного процесса обычно невелико, при наличии же большого числа генов в организме оно оказывает заметное действие на генетическую структуру популяции (в сочетании с генетической комбинаторикой).
Генетическая комбинаторика. В результате постоянных скрещиваний в популяции возникает множество новых сочетаний аллелей. Эта генетическая комбинаторика многократно изменяет значение мутаций: они входят в новые геномы, оказываются в разной генотипической среде (см. гл. 11). Потенциально число таких комбинаций имеющегося генетического материала в любой популяции невообразимо велико, но реализуется лишь ничтожная часть из этого теоретически возможного числа вариантов. Реально осуществленная часть комбинаций определяет то обстоятельство, что практически любая особь оказывается генетически уникальной. Это важно для действия естественного отбора (см. гл. 8, 10).
Комбинативная изменчивость, связанная с механизмом распределения хромосом в мейозе, случайной встречей гамет при оплодотворении и процессом кроссинговера,— мощный фактор, повышающий гетерогенность популяций. Подсчитано, что около 98% всех наследственных изменений в популяции обязано своим распространением процессу генетической комбинаторики первично сравнительно редких мутаций.
Безграничная изменчивость, однако, была бы вредна для организма, так как не позволяла бы закрепляться полезным комбинациям генов. Поэтому в эволюции выработались механизмы, не только определяющие увеличение изменчивости, но и ведущие к понижению генотипической изменчивости. На уровне особи генетическая стабильность поддерживается механизмом митоза и распределением генов в геноме по группам сцепления в определенных хромосомах, а на уровне ДНК — механизмами репарации.
На уровне же популяции такое ограничение изменчивости связано с тем или иным нарушением панмиксии (панмиксия никогда не бывает абсолютной) и гибелью части особей в процессе борьбы за существование.
Обычно по любому признаку или свойству в природных популяциях наблюдается достаточная степень изменчивости, обеспечивающая потенциальную возможность изменения данного признака или свойства в чреде поколений под давлением эволюционных факторов.
Значение мутационного процесса как элементарного эволюционного фактора. Эволюционное значение мутационного процесса определяется тем, что он постоянно поддерживает высокую степень гетерогенности природных популяций — основу для действия других факторов эволюции, прежде всего естественного отбора. Можно сказать, что мутационный процесс — это фактор- поставщик элементарного эволюционного материала.
Мутационный процесс ведет к возникновению части того «резерва» наследственной изменчивости, который определит в будущем возможность приспособления популяций к тем или иным изменениям условий среды. Образно говоря, груз мутаций, непрерывно пополняемый мутационным процессом,— это цена, которую «платит» популяция сегодня за возможность сохранения в измененных условиях завтра, приобретения новых признаков и свойств и освоения ранее недоступных условий существования.
15. Основные этапы и пути эволюции растений.
ОСНОВНЫЕ ПУТИ.
Число видов ныне существующих растений достигает более 500 тыс., из них цветковых примерно 300 тыс. видов. Остатки зеленых водорослей находят в породах архейского возраста (около 3 млрд лет назад). В протерозое в морях обитало много разных представителей зеленых и золотистых водорослей. В это же время, видимо, появились первые прикрепленные ко дну водоросли. Первичные почвообразовательные процессы в протерозое подготовили условия для выхода настоящих растений на сушу. В силуре (435–400 млн лет назад) в царстве растений происходит крупное эволюционное событие: растения (риниофиты) выходят на сушу. В первые периоды палеозоя растения населяют в основном моря. Среди прикрепленных ко дну встречаются зеленые и бурые водоросли, a в толще воды – диатомовые, золотистые, красные и другие водоросли.
С самого начала эволюции параллельно с настоящими растениями существовали и развивались группы с автотрофным и гетеротрофным питанием, взаимодополняющие друг друга в круговороте веществ. Это способствовало усилению целостности растительного мира и его относительной автономности. Первичные фототрофные низшие растения были также разнообразны по составу, среди них были группы с содержанием хлорофилла «a» и «b», с высоким содержанием каротиноидов и хлорофилла «c» и, наконец, группы с преобладанием фикобилинов. Вероятно, между этими группами организмов не было генетического единства. Разнообразие состава первичных фототрофов было вызвано, несомненно, достаточно разнородными условиями существования и позволило полнее использовать особенности среды.
В конце силура отмечено появление первых наземных растений – псилофитов, которые покрывали сплошным зеленым ковром прибрежные участки суши. Это было важным эволюционным шагом. Происходит перестройка в проводящей системе и покровных тканях: у псилофитов появляются проводящая сосудистая система со слабо дифференцированной ксилемой и флоэмой, кутикула и устьица. Псилофиты оказались и более надежно прикрепленными к субстрату с помощью дихотомически разветвленных нижних осей: у некоторых из них обнаружены примитивные «листья». Псилофиты занимали промежуточное положение между наземными сосудистыми растениями и водорослями: внешне были похожи на водоросли, тело не было дифференцировано на вегетативные органы и имело большую испаряющую поверхность.
Дальнейшая эволюция растений в наземных условиях привела к усилению компактности тела, появлению корней, развитию эпидермальной ткани с толстостенными, пропитанными восковидным веществом клетками, замене трахеид сосудами, изменению способов размножения, распространения и т.д. Самая примитивная сосудистая система состояла из трахеид. Переход от трахеид к сосудам – приспособление к засушливым условиям; с помощью сосудов создается возможность быстрого поднятия воды на большие высоты. Переход к сосудам начался в корнях, стеблях, затем в листьях.
Начальные этапы эволюции наземных растений связаны с возникновением архегониальных форм – мохообразных, папоротникообразных и голосеменных. У всех этих групп женский половой орган представлен архегонием, а мужской – антеридием. Предполагают, что архегониальные произошли от бурых или зеленых водорослей. При выходе на сушу защита гаметангиев водорослей от иссушения обеспечивалась преобразованием их в архегоний и антеридий. Этому способствовали изменение формы гаметангиев и образование у них многослойных стенок.
С момента выхода на сушу растения развиваются в двух основных направлениях: гаметофитном и спорофитном. Гаметофитное направление было представлено мохообразными, а спорофитное – остальными высшими растениями, включая цветковые. Спорофитная ветвь оказалась более приспособленной к наземным условиям: особого развития достигает корневая система, усложняется и совершенствуется проводящая система, заметно совершенствуются покровные и механические ткани, а также способы размножения (см. ниже) и создаются возможности для снижения частоты проявления возникающих летальных и других мутаций (в результате диплоидизации организма), В наземных условиях оказались непригодными для размножения свободно плавающие незащищенные половые клетки, здесь для целей размножения формируются споры, разносимые ветром, или семена.
Уже в девоне встречаются пышно развитые леса из прогимноспермов, папоротников и плаунов (рис. 5.5). Эти леса еще более распространяются в карбоне, характеризующемся увлажненным и равномерно теплым климатом в течение всего года. Мощные споровые растения – лепидодендроны и сигиллярии – достигали 40 м высоты. В карбоне же получили развитие первые семенные – голосеменные: птеридоспермы, древесные кордаиты и гинкговые, часть из которых вымирает в перми, около 280 млн лет назад.
Генеральная линия эволюции папоротникообразных на суше шла по пути преобразования спорофита (бесполое поколение). Он достиг совершенства как по разнообразию форм (деревья и травы), так и по строению. В засушливых условиях гаметофит (половое поколение) стал уже помехой из-за необходимости капельно-жидкой воды для переноса гамет. Поэтому не удивительны редукция гаметофита и значительное развитие спорофита в ходе дальнейшей эволюции наземных растений.
Одним из важных эволюционных приобретений считается появление разноспоровых папоротников, предвестников семенных растений. Начиная с лепидодендрона у некоторых плауновидных (селагинелла), папоротников и семенных папоротников закрепляется разноспоровость; в пазухах спорофитов развиваются мега- и микроспорангии. Такое событие отмечено в силуре – девоне, т.е. примерно 400 млн лет назад. Мегаспорангии имели 4 мегаспоры, а микроспорангии – множество микроспор. Дифференциация спорангиев и спор привела к появлению разных размеров гаметофитов (включая и очень мелких) и разобщению мужского и женского гаметофитов, что в конечном итоге оказало влияние на редукцию гаметофита (гаплоидного тела). Редукция гаметофита способствовала удлинению диплоидной фазы развития организма, удлинению и усложнению процессов дифференциации и онтогенеза. Не случайно, что первые разноспоровые достигали гигантских размеров; это сигиллярии, лепидодендроны, гигантские папоротники, каламиты.
Важнейшее событие в жизни голосеменных – превращение мегаспорангия в семязачаток, семяпочку с защитными покровами – интегументами и полное освобождение у всех семенных процесса полового размножения от воды. Микроспорангии у голосеменных превращаются в гнезда пыльника. Спермии большинства голосеменных неподвижны, и перенос их к архегониям осуществляется пыльцевой трубкой. Потеря мужским гаметофитом самостоятельности привела к редукции его до пыльцевой трубки с вегетативным ядром и двумя сперматозоидами. Опыление у голосеменных осуществляется ветром и нередко насекомыми, после оплодотворения семязачаток превращается в семя. Заметим, что семя появляется у семенных папоротников еще в девоне, т.е. задолго до развития цветка. Переход к семенному размножению связан с рядом эволюционных преимуществ; диплоидный зародыш в семенах защищен от неблагоприятных условий наличием покровов и обеспечен пищей, а семена имеют приспособления для распространения животными и др. Эти и другие преимущества способствовали широкому распространению семенных растений.
Непосредственные предки покрытосеменных пока не найдены в ископаемом виде. Считается, что покрытосеменные происходят от беннетитовых (С.В. Мейен). С последними примитивные покрытосеменные сходны наличием общих черт в строении древесины, устьица, пыльцы, энтомофилии и т.п. Прародиной покрытосеменных считают районы с семиаридным или сезонно сухим климатом, где они имели наибольшие шансы обживать нарушенные экосистемы благодаря высоким темпам генеративного развития и формирования зародыша (Г. Стеббинс). Аналогичными признаками как раз обладали раннемеловые беннетитовые и цикадовые. Есть предположение о появлении признаков, характеризующих цветковые (сосуды в древесине, сетчатое жилкование, завязь, рыльце, двойное оплодотворение), параллельно и порознь у разных родственных и неродственных групп. Цветковые возникают, когда все эти признаки концентрируются в одной группе. Такой путь становления характерен и для других таксонов (см. гл. 17, 20). Филогенетические взаимоотношения основных групп растений представлены на рис. 5.6.
В процессе эволюции покрытосеменных цветок (основной отличающий их орган) претерпевает значительные изменения. Ось цветка – цветоложе – постепенно укорачивается, междоузлия сближаются, спиральное расположение частей цветка переходит в циклическое, происходит процесс уменьшения числа гомологичных частей (олигомеризация).
Первые примитивные энтомофильные цветки привлекали насекомых обилием пыльцы, что одновременно способствовало перекрестному опылению. Преимущество получили те растения, у которых высокая наследственная пластичность потомства, большая вероятность опыления и завязываемости семян. В дальнейшем отбор растений пошел по пути привлечения опылителей с помощью нектара, аромата, окраски и специализации цветков на опыление определенными видами насекомых. Таким путем происходило взаимоприспособление растений и животных по соответствующим признакам. При опылении насекомыми повышается возможность свободного скрещивания растений одного вида, что и служит одной из причин высокой эволюционной пластичности цветковых растений. У цветковых (в отличие от голосеменных) даже деревья представлены множеством разнообразных форм. Цветковые также были приспособлены использовать среду путем быстрого развития и накопления органического вещества.
В кайнозое (начало – 66 млн лет назад) вся Европа была покрыта пышными лесами теплого и умеренного климатов, включающими дуб, березу, сосну, каштан, бук, виноград, орех и др. В это время леса достигали наибольшего распространения на Земле. В тропической флоре этого периода были представлены фикусы, лавровые, гвоздичные, эвкалипты, виноград и др.
В четвертичном периоде кайнозойской эры (2 млн лет назад) увеличилось количество осадков и наступило оледенение значительной части Земли, вызвавшее отступление теплолюбивой третичной растительности на юг (а местами полное ее вымирание), возникновение холодоустойчивых травянистых и кустарниковых растений. На огромных территориях завершается начатая в миоцене смена лесов степью, формируется ксерофитная и эфемерная растительность с выраженной сезонностью в цикле развития, складываются современные фитоценозы.
Таким образом, растительность нашей планеты постоянно менялась, приобретая все более современные черты.
Основные черты эволюции царства растений следующие:
1. Переход от гаплоидности к диплоидности. С диплоидизацией организма снижался эффект проявления неблагоприятных мутаций, усиливались морфогенетические потенции организма. У многих водорослей все клетки (кроме зиготы) гаплоидны. У более высокоорганизованных водорослей (бурые и др.) наряду с гаплоидными существуют и диплоидные особи. У мхов преобладает гаплоидное поколение при сравнительно слабом развитии диплоидного. У папоротников преобладает диплоидное поколение, однако и у них гаплоидное поколение (гаметофит) еще представлено самостоятельным образованием, у голосеменных и покрытосеменных наблюдается почти полная редукция гаметофита и переход к диплоидной фазе (рис. 5.7).
2. Утрата связи процесса полового размножения с наличием капельно-жидкой воды, потери подвижности мужских гамет, заметная редукция гаметофита и сильное развитие спорофита, переход от наружного оплодотворения к внутреннему, возникновение цветка и двойного оплодотворения.
3. Дифференциация тела с переходом к наземным условиям: деление на корень, стебель и лист, развитие сети проводящей системы, совершенствование покровных, механических и других тканей.
4. Специализация опыления (с помощью насекомых) и распространение семян и плодов животными. Усиление защиты зародыша от неблагоприятных условий: обеспечение пищей, образование покровов и др.
16. Эмбрионизация и автономизация онтогенеза.
Эмбрионизация онтогенеза — это возникновение в процессе эволюции способности к прохождению части стадий развития под защитой материнского тела или специальных (семенных или яйцевых) оболочек. Эмбриональное развитие — не изначальное свойство живого, а результат эволюции. Эмбрионизация при этом отражает тенденцию развития все усложняющегося зародыша в более защищенной и постоянной внутренней среде (табл. 14.2). При этом имеет место конвергенция функций в онтогенезе животных и растений — результат сходно направленного действия отбора (А.А. Захваткин)
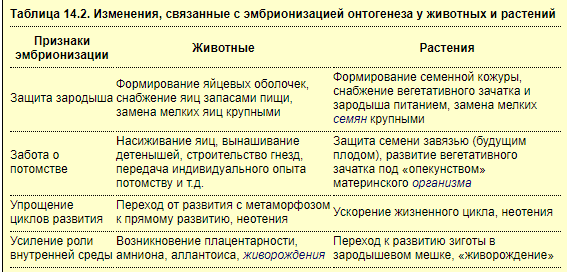
В приспособительном значении эмбрионизации можно убедиться на примере эволюции разных типов эмбрионального развития у животных: первично-личиночного, неличиночного и вторично-личиночного. Исходным в эволюции является первично-личиночный тип развития, характерный для животных, откладывающих мелкие яйца с небольшим запасом желтка (кишечнополостные, губки, полихеты, ракообразные, пресноводные костистые рыбы, амфибии). Личинка у этих животных свободная и может существовать самостоятельно. При переходе к образованию крупных яиц, содержащих больше желтка, личиночный тип сменяется неличиночным развитием (головоногие моллюски, акулы, миксины, некоторые земноводные, рептилии, птицы, яйцекладущие млекопитающие и др.). При неличиночном развитии зародыш долгое время находится под защитой яйцевых оболочек, потребляя запасы пищи из яйца. В приспособлении позвоночных к наземным условиям большое значение имел переход к неличиночному развитию. У рептилий и птиц по сравнению с амфибиями эмбрионизация онтогенеза усиливается за счет добавления плодного периода и приводит к освобождению начальных стадий развития от водной среды путем создания необходимых условий для развития зародыша в яйце. Запасы пищи в яйце (белки и жиры) увеличиваются, формируются амнион, желточный мешочек, аллантоис, хорион-сероза, изменяются способ дробления и тип эмбриогенеза. Все это определяет возможность развития яиц на суше и выход из них особи, сразу способной к активной самостоятельной жизни.
В условиях, когда зародыш развивается в теле другого вида (паразитизм) или матери, отбор идет в направлении измельчения яиц и возникновения вторично-личиночного типа развития (пиявки, дождевые черви, жуки, муравьи, скорпионы, сальпы, сумчатые и плацентарные млекопитающие). Вторично-личиночный тип отличается от первично-личиночного большей независимостью от среды по способу питания, в том числе и возможностью утилизации разнообразных продуктов питания. У цветковых паразитов наблюдается своеобразная редукция зародыша и семян (дезэмбрионизация). Из-за узкой специализации зародыш у них выполняет только функцию присоски-внедрения в тело хозяина.
В целом эмбрионизация онтогенеза сопровождается у растений и животных многими изменениями и приводит к усилению роли внутренней среды в развитии зародыша и эмансипации его от внешней среды. Одно из важных результатов эмбрионизации — снабжение зародыша необходимой пищей и достижение быстрого его развития. В связи с защищенностью зародыша при усилении эмбрионизации отбор идет на уменьшение числа яиц и зародышей, повышение выживаемости зародышей. Высшим этапом эмбрионизации является живорождение, связанное с плацентацией и вскармливанием детенышей молоком. В силу возрастания защищенности зародыша и усиления системы морфогенетических корреляций эмбриональное развитие у млекопитающих делается особенно консервативным по сравнению с таковым других животных (включая рептилий и птиц). Консервативность эмбриональных стадий, видимо, обезвреживает эффект малых мутаций, а при возникновении мутаций, выходящих за пределы допустимых пороговых уровней, способствует элиминации их носителей. Так, эмбрионизация способствует усилению целостности онтогенеза в эволюции, явлениям неотении и фетализации.
Автономизация — главное направление эволюции онтогенеза
В процессе эволюции онтогенеза происходит возникновение разнообразных регуляторных механизмов и повышение устойчивости онтогенетического развития в целом. Наблюдается постепенное сокращение детерминирующей роли физико-химических факторов внешней среды в индивидуальном развитии. Постоянно повторяющиеся факторы среды из факторов, детерминирующих развитие, превращаются в условия нормального развития. Организм постепенно «освобождается» от влияния случайных и кратковременных изменений среды; в процессе эволюции он становится все более автономным, внешние факторы его развития заменяются внутренними.
Повышение устойчивости индивидуального развития делает его независимым от разрушающих влияний внешней среды. Процесс сокращения детерминирующего значения физико-химических факторов внешней среды, ведущий к возникновению относительной устойчивости развития, называется автономизацией онтогенеза в ходе эволюции (И.И. Шмальгаузен).
Автономизация онтогенеза выявляется и в сохранении способности к нормальному развитию у теплокровных животных при широких колебаниях температуры. Нечто сходное есть и в развитии растений. У птицемлечника Воронова (Ornithogalum woronowii) и безвременника веселого (Colchicum laetum) закладка зачатков цветков в естественных условиях обычно наступает при температуре почвы 25—20°C. Однако этот процесс может протекать и при значительных отклонениях температуры от оптимума.
Часто для реализации в онтогенезе унаследованной программы требуется наличие лишь минимума внешних условий. Так, образование хлорофилла у растений идет при коротких световых вспышках, а индукция цветения наступает при наличии минимума благоприятных условий (фотопериод, низкие температуры, минеральное питание).
Механизм индивидуального развития, зависимый от минимума интенсивности внешнего фактора, дающего лишь первый толчок к развертыванию формообразования, называется авторегуляторным механизмом развития (И.И. Шмальгаузен).
При авторегуляторном развитии роль большинства изменений внешних условий сводится к «пуску» внутренних механизмов морфогенеза. Это происходит в тех случаях, когда отбор эволюционно-длительное время бывает направлен на установление связи развития организма с частыми колебаниями физических условий среды. С появлением авторегуляции устойчивость онтогенеза в целом возрастает и он может протекать даже при относительно неблагоприятных условиях.
В идеальном случае автономизация онтогенеза сопровождается полной заменой внешних факторов развития внутренними. Примером может служить процесс развития легких у позвоночных (рис. 14.4). У аксолотля респираторная часть легкого развивается после растяжения легочных мешочков воздухом, т. е. после начала атмосферного дыхания. У личинок лягушек до начала воздушного дыхания имеет место первичная фрагментация ткани легкого. У более приспособленных к наземным условиям жаб до периода воздушного дыхания респираторная структура развивается еще сильнее, чем у лягушек. У рептилий и млекопитающих легочные структуры формируются еще на более ранних стадиях эмбриогенеза задолго до их функционирования.
17. Основные этапы и пути эволюции животных.
Основные пути эволюции животных. Царство животных не менее разнообразно, чем царство растений, а по числу видов животные превосходят растения. Описано около 1 млн 200 тыс. видов животных (из них около 900 тыс. видов – членистоногих, 110 тыс. – моллюсков, 42 тыс. – хордовых животных) и считается, что это может быть лишь половина существующих видов.
Возникновение животных в ископаемых остатках не прослеживается. Первые останки животных находят в морских отложениях протерозоя, возраст которых превышает 1 млрд лет. Первые многоклеточные животные представлены сразу несколькими типами: губки, кишечнополостные, плеченогие, членистоногие.
В морях кембрийского периода уже существовали все основные типы животных. Облик фауны определили многочисленные хелицеровые (похожие на современных мечехвостов), губки, кораллы, иглокожие, разнообразные моллюски, плеченогие, трилобиты (рис. 5.8). После кембрия эволюция животных характеризовалась лишь специализацией и совершенствованием основных типов. Исключение составляют позвоночные, останки которых обнаружены в ордовике. Это были так называемые щитковые – существа, отдаленно сходные с современными круглоротыми (миноги, миксины), но покрытые со спинной стороны мощно развитыми костными пластинами. Предполагают, что они защищали первых мелких (около 10 см длиной) позвоночных от огромных хищных ракообразных.
В теплых и мелководных морях ордовика обитали многочисленные кораллы, значительного развития достигали головоногие моллюски – существа, похожие на современных кальмаров, длиной в несколько метров.
Силурийский период ознаменовался важными событиями не только для растений, но и для животных. Появились животные, дышащие воздухом. Первыми обитателями суши были паукообразные, напоминавшие по строению современных скорпионов. Тем временем в водоемах происходило бурное развитие разнообразных низших позвоночных, прежде всего панцирных рыб. Предполагается, что первые позвоночные возникли в мелководных пресных водоемах. Постепенно, в течение девона, эти пресноводные формы завоевывают моря и океаны. В девоне же возникают двоякодышащие, кистеперые и лучеперые рыбы. Все они были приспособлены к дыханию в воде. До наших дней дожили некоторые виды двоякодышащих (рис. 5.9), лучеперые дали начало современным костистым рыбам, а кистеперые – первичным земноводным (стегоцефалам). Стегоцефалы появились в верхнем девоне; примерно в это же время возникает другая чрезвычайно прогрессивная группа животных – насекомые.
В развитии линий позвоночных и беспозвоночных проявились две разные тенденции в решении одних и тех же задач. Переход в воздушную среду из водной потребовал укрепления основных несущих органов и всего тела в целом. У позвоночных роль каркаса играет внутренний скелет, у высших форм беспозвоночных – членистоногих – наружный скелет. Развитие в среде, требовавшей все более сложных поведенческих реакций, решалось в этих двух ветвях древа жизни двумя принципиально разными способами. У насекомых чрезвычайно сложная нервная система, с разбросанными по всему телу огромными и относительно самостоятельными нервными центрами, преобладание врожденных реакций над приобретенными. У позвоночных – развитие огромного головного мозга и преобладание условных рефлексов над безусловными.
В каменноугольном периоде появляются первые пресмыкающиеся, что определило начало активного завоевания суши позвоночными. Рептилии благодаря сухим прочным покровам, яйцам, покрытым твердой скорлупой и не боящимся высыхания, были мало связаны с водоемами. В этом периоде возникают и достигают значительного развития такие древнейшие группы насекомых, как стрекозы и тараканы.
В пермском периоде начинают исчезать стегоцефалы и широко распространяются различные рептилии. От примитивных рептилий из группы цельночерепных в это время развивается ветвь пеликозавров, приведшая несколько позже – через терапсид – к возникновению млекопитающих.
В конце палеозоя происходит значительное иссушение климата. Поэтому бурное развитие претерпевают разнообразные рептилии; до наших дней из триасовых рептилий дожили гаттерия и черепахи. Некоторые рептилии становятся хищниками, другие – растительноядными, третьи – вторично возвращаются в водную среду (рис. 5.10), обеспечивающую им пищу в виде многочисленных форм костистых рыб и головоногих моллюсков. Однако особенно сильного развития достигают морские рептилии в юре (ихтиозавры, плезиозавры). Тогда же пресмыкающиеся осваивают и воздушную среду – возникают птерозавры, видимо, охотившиеся на многочисленных и крупных насекомых. В триасе от одной из ветвей рептилий возникают птицы; первые птицы причудливо сочетали признаки рептилий и птиц (см. рис. 6.3).
В меловом периоде продолжается специализация рептилий: возникают гигантские растительноядные динозавры, встречаются летающие ящеры с размахом крыльев до 20 м. Знаменательные события происходят и в мире насекомых – начинается активная сопряженная эволюция энтомофильных растений и насекомых-опылителей. Происходит процесс вымирания аммонитов, белемнитов, морских ящеров. В связи с сокращением пространств занятых богатой прибрежной растительностью, вымирают растительноядные динозавры, а следом – и охотившиеся на них хищные динозавры. Лишь в тропическом поясе сохраняются крупные рептилии (крокодилы). В условиях похолодания исключительные преимущества получают теплокровные животные – птицы и млекопитающие, которые пышно расцветают лишь в следующем периоде – кайнозое.
Кайнозой – время расцвета насекомых, птиц и млекопитающих. В конце мезозоя возникают плацентарные млекопитающие. В палеоцене и эоцене от насекомоядных происходят первые хищники. В это же время или несколько позже первые млекопитающие начинают завоевывать море (китообразные, ластоногие, сиреновые). От древних хищных происходят копытные, от насекомоядных обособляется отряд приматов. К концу неогена встречаются уже все современные семейства млекопитающих, на обширных открытых пространствах саванн Африки появляются многочисленные формы обезьян, многие из которых переходят к прямохождению. Одна из групп таких обезьян – австралопитеки – дала ветви, ведущие к роду Homo (см. гл. 18).
В кайнозое особенно четко проявляются тенденции в развитии самых прогрессивных ветвей древа жизни животных, ведущих к возникновению стайного, стадного образа жизни (что стало ступенькой к возникновению социальной формы движения материи).
В четвертичном, или антропогеновом, периоде кайнозоя наблюдались резкие изменения климата нашей планеты, в основном связанные с постепенным похолоданием. На этом общем фоне неоднократно повторялись фазы особенно резкого похолодания, при которых в средних широтах Северного полушария возникали значительные оледенения суши.
Максимального распространения материковые оледенения достигали во время среднего плейстоцена – около 250 тыс. лет назад. На территории Европы в плейстоцене насчитывается по крайней мере пять таких ледниковых периодов
Огромное значение для эволюции современной фауны имело то обстоятельство, что одновременно с наступлением ледниковых периодов происходили значительные колебания уровня Мирового океана: в разные периоды этот уровень понижался или повышался на сотни метров сравнительно с современным. При таких колебаниях уровня океана могла обнажаться большая часть материковой отмели Северной Америки и Северной Евразии. Это, в свою очередь, вело к появлению сухопутных «мостов» типа Берингийской суши, соединявшей Северную Америку и Северную Евразию, соединению Британских островов с европейским материком и т.п. В Европе 5–6 тыс. лет назад климат был заметно теплее современного. Однако эти изменения климата уже не играли столь значительной роли в изменении видового состава животного мира, какую стал играть Человек, не только уничтоживший многие виды животных и растений (по некоторым подсчетам, человек к середине XX в. уничтожил более 200 видов животных), но и создавший новых домашних животных и ставящий сейчас грандиозную задачу управления эволюционным процессом.
В эволюции животных можно наметить несколько магистральных направлений развития адаптации:
1. Возникновение многоклеточности и все большее дифференцирование всех систем органов.
2. Возникновение твердого скелета (наружного – у членистоногих, внутреннего – у позвоночных).
3. Развитие центральной нервной системы. Два разных и чрезвычайно эффективных эволюционных «решения»: у позвоночных развитие головного мозга, основанного на обучении и условных рефлексах, и возрастание ценности отдельных особей; у насекомых – развитие нервной системы, связанной с наследственным закреплением любого типа реакций по типу инстинктов.
4. 4. Развитие социальности в ряде ветвей древа животных, с разных сторон подходящих к рубежу, отделяющему биологическую форму движения материи от социальной формы движения. Перешагнуть этот рубеж смогла лишь одна ветвь приматов – род Человек.
18. Недарвиновская эволюция или теория нейтральности.
Нейтрализм
В связи с успехами молекулярной биологии в анализе генетического кода возникла концепция так называемой недарвиновской эволюции (Д. Кинг, Т. Юкес, 1969). Эта концепция основана на предположении о селективной нейтральности многих мутаций. В свою очередь, предположение о нейтральности мутаций опирается на факты вырожденности генетического кода: не один, а несколько (до шести) нуклеотидных триплетов могут кодировать синтез ряда аминокислот. Например, синтез аминокислоты серина кодируется триплетами УЦУ, УЦЦ, УЦА, УЦГ, АГУ, АГЦ и изменения (мутация) в одном из триплетов не скажутся на результате синтеза. Таких нейтральных мутаций должно быть много — около 30% от возникающих на молекулярном уровне, и их распространение в популяции, не контролируемое естественным отбором, позволяет предполагать большую роль стохастических, ненаправленных процессов на молекулярном уровне. Исходя из нейтральности этих мутаций предполагается также, что их накопление в геноме может идти с постоянной скоростью. Таким образом, обнаруживая различия в строении белков разных организмов, можно вычислить время расхождения их от общего предка (концепция молекулярных часов) (см. 6.2).
Общая концепция молекулярной эволюции сформулирована в виде следующих эмпирических правил (М. Кимура): скорость эволюции любого белка (в полингах) постоянна и одинакова в разных филогенетических ветвях (1), функционально менее важные молекулы или их части эволюционируют быстрее важных (2), мутационные замены аминокислот с меньшими нарушениями структуры и функции белков являются наиболее обычными (3), функционально новый ген появляется в результате дупликации (4) и случайная фиксация нейтральных (или очень слабо вредных) мутаций в ходе эволюции происходит чаще благоприятных мутаций (5).
Как видно, концепция нейтрализма не отказывается полностью от приспособительного характера эволюции и на молекулярном уровне. Так, в одних белках (фибринопептиды) аминокислотные замены наблюдаются чаще других (гистоны). У первых скорость эволюционных замен аминокислот совпадает со скоростью возникновения мутаций, так как замены не влияют на свойства белка. В гемоглобине ввиду чрезвычайной важности его функции замены наблюдаются реже, чем в молекуле фибринопептидов. У цитохрома C (взаимодействует с цитохромоксидазой и редуктазой) скорость эволюционных изменений еще ниже гемоглобина. Гистон H4 как белок, участвующий в регуляции генетической информации, эволюционно оказывается еще консервативнее цитохрома C.
В последние годы идеи нейтрализма часто рассматриваются в свете гипотезы молекулярного дрейфа — влияния на эволюционный процесс выпадающей из-под действия естественного отбора так называемой «эгоистической» ДНК. Эта «эгоистическая» ДНК состоит из не кодирующих последовательностей нуклеотидных пар и поэтому не должна влиять на фенотип особи. К некодирующим относятся мобильные генетические элементы (мобильные диспергированные гены). У эукариот они могут составлять до 20% генома. К «эгоистической» относят также сателлитную ДНК, которая может составлять до половины всей ДНК хромосом и которая, как недавно предполагалось, не транскрибируется и не несет никакой генетической информации.
Сейчас стало известно, что сателлитная ДНК играет существенную роль в митозе и мейозе и ее определение как «эгоистической» неоправданно. То же относится и к ДНК теломер, находящихся на концах хромосом, и к спейсерам (отрезкам ДНК, разделяющим гены), и к интронам (некодирующим участкам внутри гена). Все эти структуры ДНК оказывают то или иное влияние на проявление генов и доступны для естественного отбора.
Представление о селективной нейтральности каких-то участков генетического кода не выдержало проверки: все сначала считавшиеся нейтральными изменения генетического материала в ходе детального исследования оказывались связанными с особенностями фенотипа, т. е. доступными для естественного отбора. Таким образом, молекулярный дрейф не является самостоятельным эволюционным фактором, а лишь увеличивает частоту мутаций, поставляя тем самым материал для естественного отбора.
Изучение белкового полиморфизма разных популяций и подвидов некоторых видов дрозофилы показало, что все перестановки в последовательности аминокислот оказываются в популяциях в состоянии сбалансированного полиморфизма и часто носят клинальный характер. Эти два обстоятельства указывают на серьезное адаптивное значение таких изменений (хотя адаптивное значение каждой конкретной перестановки неясно).
В свое время критика ведущего, творческого значения отбора в эволюции часто была вызвана недостаточно глубоким изучением биологии отдельных видов. Например, возникало сомнение в том, как отбор может «улавливать» ничтожные изменения характера распределения пигмента на крыле бабочки или едва заметные изменения общего тона окраски полевок. Детальное изучение таких случаев в природе показало высокую эффективность отбора и его способность «подхватывать» даже такие признаки.
Еще Ч. Дарвин и С.С. Четвериков обращали внимание на возможность возникновения безразличных наследственных уклонений. Впоследствии Дж. Холдейн привел расчеты, свидетельствующие о возможности снижения приспособленности популяций в результате увеличения полиморфизма (концепция генетического груза). По его мнению, отбор, направленный по одному или немногим аллелям, в конце концов должен привести к снижению гетерозиготности. Однако работы, связанные с изучением тонких биохимических изменений методом электрофореза, показали присутствие значительного числа вариантов белков (энзимов) в каждом из локусов при анализе больших популяционных выборок. Наличие такой гетерогенности — очевидный результат действия отбора, идущего всегда одновременно по многим аллелям (см. гл. 10). Отбор по многим аллелям и соотбор, приводящий к возникновению шлейфа автоматически соотбираемых мутаций, показывают, что в конечном счете нет мутаций нейтральных, вредных или полезных безотносительно к условиям. Любая мутация может быть в одном случае явно вредной, в другом — явно полезной, а в третьем, при рассмотрении ее без учета целостности генотипа, кажущейся нейтральной. Поэтому утверждать, что число различий в последовательности аминокислот оказывается нейтральным,— это значит принимать наши современные, неизбежно ограниченные знания за абсолютную истину. Логичнее все же предположить, что такие различия, совпадающие с уровнем морфофизиологической дивергенции сравниваемых форм (табл. 20.1), отражают какие-то неясные еще сегодня, но тем не менее функционально важные адаптивные особенности групп. «Недарвиновская эволюция» на молекулярном уровне за счет нейтральных мутаций и дарвиновский естественный отбор взаимодополняющие и не противоречащие друг другу концепции (Ю.П. Алтухов, Н.Н. Воронцов).
19. Правила эволюции групп.
Правило необратимости эволюции (Л. Долло, 1893) гласит, что эволюция — процесс необратимый и организм не может вернуться к прежнему состоянию, уже осуществленному в ряду его предков. Так, если в эволюции наземных позвоночных на каком-то этапе от примитивных амфибий возникли рептилии, то рептилии, как бы ни шла дальше эволюция, не могут вновь дать начало амфибиям. Вернувшись в просторы Мирового океана, рептилии (ихтиозавры) и млекопитающие (киты) никогда не становятся рыбами. Можно сказать, что если какая-то группа организмов в процессе эволюции вновь «возвращается» в адаптивную зону существования ее предков, то приспособление к этой зоне у «вернувшейся» группы будет неизбежно иным.
Правило необратимости эволюции в наше время получило существенное уточнение. Успехи генетики позволяют говорить о возможности повторного возникновения признаков на основе обратных мутаций.
Но признание обратимости отдельных признаков (П.П. Сушкин и др.) в филогенезе — отнюдь не признание обратимости эволюционного процесса в целом. Обратная мутация по признаку может привести к повторному возникновению данного аллеля, но не генотипа в целом; к вторичному появлению данного признака, но не фенотипа в целом. Статистически вероятно повторное возникновение мутаций, но статистически невероятно повторное возникновение генных комплексов и целых фенотипов (Н.Н. Воронцов).
Правило прогрессирующей специализации (Ш. Депере, 1876) гласит, что группа, вступившая на путь специализации, как правило, в дальнейшем развитии будет идти по пути все более глубокой специализации.
Если в процессе эволюции одна из групп позвоночных, скажем ветвь рептилий, приобрела адаптации к полету, то на последующем этапе эволюции это направление адаптации сохраняется и усиливается (например, птеродактили в свое время все более приспосабливались к жизни в воздухе). Поскольку организм определенного строения не может жить в любой среде, в выборе адаптивной зоны или ее части группа ограничена особенностями строения. Если эти особенности несут черты специализации, то организм обычно «выбирает» (точнее, в результате борьбы за существование попадает во все более частную среду), где его специализированные приспособления могут обеспечить успешное выживание и оставление потомства. Но обычно это ведет лишь к дальнейшей специализации.
Частный случай этого общего правила прогрессивной специализации — увеличение размеров тела особей в процессе эволюции позвоночных животных. Увеличение размеров тела, с одной стороны, связано с более экономным обменом веществ (уменьшение величины относительной поверхности тела) и должно рассматриваться как частный случай специализации. С другой стороны, дает хищнику преимущества в нападении, а жертве — преимущества в защите. Связь организмов в цепях питания неизбежно вызывает увеличение размеров тела во многих группах. У представителей других групп происходит уменьшение размеров тела. Например, при переходе к подземному образу жизни и обитанию в закрытых норах многие грызуны вторично стали более мелкими. Интересно, что сопряженной эволюции подверглась ласка (Mustela nivalis) — один из наиболее облигатных потребителей мышевидных грызунов в средней полосе. Ласка приобрела размеры тела, позволяющие преследовать мелких грызунов в норах. На этом примере видно, что выделяемые эмпирические эволюционные правила имеют относительное значение. Характер эволюции зависит в конечном счете от конкретных связей группы с элементами биотической и абиотической среды (всегда при постоянном контроле отбора, идущем на уровне микроэволюционных взаимодействий внутри популяций и биогеоценозов).
Правило происхождения от неспециализированных предков (Э. Коп, 1896) гласит, что обычно новые крупные группы берут начало не от специализированных представителей предковых групп, а от сравнительно неспециализированных. Млекопитающие возникли не от высокоспециализированных форм рептилий, а от неспециализированных. Голосеменные растения возникли от неспециализированных палеозойских папоротникообразных. Ныне процветающая группа цветковых растений возникла не от специализированных голосеменных, а от неспециализированных предков, занимающих промежуточное положение между семенными папоротниками и беннетитовыми (см. гл. 6).
Причина происхождения новых групп от неспециализированных предков в том, что отсутствие специализации определяет возможность возникновения новых приспособлений принципиально иного характера. Трудно ожидать появления среди гельминтов каких-то принципиально новых форм. У таких форм скорее могут возникнуть эволюционные изменения, направленные к лучшему прикреплению внутри организма хозяина, лучшему использованию питательных веществ, более эффективному способу размножения и т. п. Напротив, у такой группы, как практически всеядные средних размеров хищные, живущие в разнообразных условиях, большие потенциальные возможности развития в самых разных направлениях.
Высокая потентность неспециализированных групп в эволюции определяется борьбой за существование, которую они вынуждены вести. Но в историческом плане именно эти жесткие требования среды и приводят потомков неспециализированных групп на путь арогенеза, вызывая к жизни приспособления, которые потом оказываются перспективными для начала нового широкого аллогенеза (см. гл. 15).
Правило происхождения от неспециализированных предков оказывается, однако, далеко не всеобщим. Редко бывает так, чтобы специализация затронула в равной степени все без исключения органы и функции организма (вида). Эти остающиеся достаточно лабильными органы и системы могут открыть группе дорогу в другую адаптивную зону. Переход к птицам был осуществлен в группе рептилий, уже способных к лазанию и прыганию по ветвям деревьев, т. е. в специализированной с точки зрения развития приспособлений к передвижению группе. Переход к наземному образу жизни среди позвоночных был осуществлен формами, способными, по-видимому, передвигаться по твердому субстрату, заглатывать и использовать для дыхания кислород воздуха, т. е. опять-таки формами, крайне специализированными в этом отношении по сравнению с большинством обитавших в водоемах видов рыб. Оказывается, признаки, специализированные в одной среде, могут быть эрогенными в иной (М.С. Гиляров). Ясно, что как неспециализированные, так и специализированные формы могут при определенных условиях стать родоначальниками новых, прогрессивных ветвей древа жизни.
Правило адаптивной радиации (Г.Ф. Осборн, 1902) гласит, что филогенез любой группы сопровождается разделением группы на ряд отдельных филогенетических стволов, которые расходятся в разных адаптивных направлениях от некоего исходного среднего состояния. По существу, это правило не что иное, как принцип дивергенции, подробно описанный Ч. Дарвином (1859) при обосновании гипотезы естественного отбора. Дарвин говорил о внутривидовой приспособительной дивергенции к различной пище, несколько различным условиям существования и т. п. и рассматривал ее как обязательный этап образования новых видов. В дальнейшем принцип дивергенции был положен в основу представлений эволюционной морфологии об идиоадаптациях (А.Н. Северцов) и соответственно о развитии группы по пути аллогенеза, а в эволюционной палеонтологии — в основу представлений об «иррадиации» (В.О. Ковалевский, 1875) в процессе становления каждого крупного филума (рис. 15.15, см. рис. 6.4) и классов в систематике (Дж. Хаксли).
Правило чередования главных направлений эволюции. Арогенная эволюция чередуется с периодами аллогенной эволюции во всех группах. И.И. Шмальгаузен (1939) сформулировал это правило как чередование фаз адаптациоморфоза.
Эволюция представляет непрерывный процесс возникновения и развития новых и новых адаптаций — адаптациогенез. Одни из вновь возникающих адаптаций оказываются очень частными, и их значение не выходит за пределы узких условий. Другие дают возможность выхода группы в новую адаптивную зону и непременно ведут к быстрому эволюционному развитию групп в новом направлении. Обычно одна из форм получает какую-то новую адаптацию (или комплекс адаптаций), оказывающуюся основой для следующего быстрого периода филогенетических новообразований
Правило усиления интеграции биологических систем (И.И. Шмальгаузен, 1961) можно сформулировать так: биологические системы в процессе эволюции становятся все более интегрированными, со все более развитыми регуляторными механизмами, обеспечивающими такую интеграцию.
Сейчас в общих чертах известны основные направления такой интеграции, идущей на уровне популяции и биогеоценозов. На уровне популяции это означает поддержание определенного уровня гетерозиготности, которая является основой интеграции всего популяционного генофонда в сложную, лабильную и одновременно устойчивую генетическую систему, способную к саморегуляции (численности, структуры), генетико-экологическому гомеостазису.
Межпопуляционные взаимоотношения интегрируют отдельные популяции и их группы в вид как систему (см. гл. 12, 13). Интеграция в системе биогеоценоза определяет образование разными видами многих взаимодополняющих друг друга экологических ниш и слаженной работой биогеоценоза как элементарной биохорологической структуры в биосфере нашей планеты (с упорядоченными потоками вещества, энергии, информации). Высший уровень интеграции выражается в эволюции самих механизмов эволюции — проблема, которая начинает вырисовываться в современном эволюционном учении.
Несомненно, возможно выделение и других кроме шести выделенных нами выше «правил». Тут открыто широкое поле деятельности для исследователя-эволюциониста.
20. Значение эволюционного учения.
Знание закономерностей эволюционного развития живой природы важно по двум причинам. Во-первых, только эволюционный подход дает возможность достаточно глубоко понять, обобщить и сопоставить данные разных специальных биологических дисциплин. Во-вторых, эволюционный подход необходим для сознательного планирования, предвидения результатов вмешательства человека в развитие биосферы Земли.
Дата добавления: 2021-06-02; просмотров: 427; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!