Механизм возникновения адаптаций
ВОПРОСЫ К ЭКЗАМЕНУ
По дисциплине «Теория эволюции»
1. Натурфилософия античных мыслителей.
2. Внутрипопуляционный полиморфизм.
3. Основные биологические обобщения метафизического периода: систематизации (К. Линней), преформизма (Ш. Бонне), эпигенеза (К.Вольф), трансформизма (Ж. Бюффон, Ж. Сент-Иллер), креационизма (К. Линней).
4. Принцип основателя и видообразования.
5. Эволюционная теория Ч. Дарвина.
6. Методы изучения микроэволюции.
7. Первая теория эволюции органического мира Ж. Б. Ламарка.
Создателем первой эволюционной теории стал выдающийся французский естествоиспытатель Жан Батист Ламарк. Учёный считал, что наиболее общие категории явлений, такие как пространство, движение, материя и время, созданы Богом, а все остальные объекты образованы самой природой. Своей задачей Ламарк считал поиск того пути, по которому шла природа, формируя существующее многообразие живых существ. Эволюционную теорию Ламарк изложил в двухтомном труде «Философия зоологии» (1809). Учёный определил два основных направления эволюционного процесса: постоянное усложнение уровня организации живых существ, происходящее во времени (градация, от лат. gradatio – постепенное повышение), и увеличение разнообразия под действием условий среды. Таким образом, эволюционную теорию Ламарка можно разделить на две части: учение о градации организмов и учение об изменчивости.
|
|
|
Учение о градации организмов. Ламарк считал, что первые организмы произошли из неорганической природы путём самозарождения. Их дальнейшее развитие привело к усложнению живых существ, поэтому классификация организмов не может быть произвольной, она должна отображать процесс движения от низших форм к высшим. Всех животных учёный разделил на 14 классов, которые распределил по степени усложнения организации, образовав 6 ступеней – градаций (рис. 4). Самый низший уровень в этой системе занимали инфузории, наиболее высокий – млекопитающие. Для того чтобы объяснить механизм усложнения живых существ, Ламарк предположил существование у всех организмов стремления к совершенствованию, изначально заложенного в них Богом (принцип самосовершенствования). Одновременное наличие в природе и простых, и более сложных организмов Ламарк объяснял постоянно продолжающимся процессом самозарождения жизни.
Учение об изменчивости. Совершенствуясь, организмы вынуждены приспосабливаться к условиям внешней среды. Для того чтобы объяснить, как возникает разнообразие на каждой ступени «лестницы существ», Ламарк сформулировал два закона.
Закон упражнения и неупражнения органов: постоянное употребление органа ведёт к его усиленному развитию, а неупотребление – к ослаблению и исчезновению. Согласно этому закону необходимость доставать листья на деревьях ведёт к тому, что жираф, стараясь до них дотянуться, постоянно вытягивает шею, в результате чего она становится длинной. Муравьеду, чтобы доставать муравьёв из глубины муравейника, приходится вытягивать язык, и он становится тонким и длинным, а перепонки между пальцами ног у водоплавающей птицы возникают из-за постоянного раздвигания пальцев и растягивания кожи во время плавания. Примером исчезновения органов в результате неупражнения является редукция глаз у крота.
|
|
|
Закон наследования благоприобретённых признаков: под действием постоянных упражнений и неупражнений органы изменяются, и возникшие изменения наследуются. По мнению Ламарка, вытянувшаяся в течение жизни шея жирафа будет передана следующему поколению, которое родится уже с более длинной шеей. Открытие в XX в. материальной основы наследственности – ДНК – окончательно опровергло возможность наследования благоприобретённых признаков.
Для того чтобы доказать, что признаки, приобретённые в течение жизни, не передаются по наследству, известный исследователь Август Вейсман отрезал хвосты подопытным мышам на протяжении 22 поколений. Однако никакого укорочения хвостов у потомков не произошло.
|
|
|
Значение теории Ламарка. Учение Ламарка стало первой целостной эволюционной теорией. Учёный определил предпосылки эволюции (изменчивость и наследственность) и указал направление эволюции (усложнение организации). Однако, правильно оценив развитие природы от простого к сложному, Ламарк не смог вскрыть причины эволюции. Созданная теория не могла объяснить многие существующие явления, такие как наследование нефункционирующих признаков (например, рудиментарные органы), появление мимикрии или покровительственной окраски.
Эволюционные идеи Ламарка не нашли поддержки у современников и подверглись критике со стороны многих учёных, одним из которых был Жорж Кювье – основоположник сравнительной анатомии и палеонтологии.
8. Примеры и механизмы возникновения адаптаций. Классификация адаптаций.
Механизм возникновения адаптаций
В широком смысле слова под адаптацией понимается гармония организмов (в том числе и популяций, видов) со средой обитания. В узком смысле под адаптацией понимают специальные свойства, способные обеспечить выживание и размножение организмов в конкретной среде. Из этого ясно, что адаптации являются относительными: адаптация к одним факторам среды не обязательно останется таковой в других условиях.
|
|
|
Для возникновения адаптации необходимо наличие элементарного эволюционного материала — наследственной изменчивости — и элементарных эволюционных факторов, прежде всего отбора. Появление в популяции и биогеоценозе нового удачного фенотипа или особей — носителей удачных мутаций — еще нельзя рассматривать как адаптацию. Появление селективно ценного генотипа является элементарным адаптационным явлением. Так же как появление элементарного эволюционного явления — изменения генотипического состава популяции — еще не есть эволюционный процесс, так и появление элементарного адаптивного явления еще не означает возникновения адаптации. Об адаптации можно говорить лишь после возникновения специализированного признака у популяции (вида) к элементам среды. Достигается это при «подхвате» отбором элементарного адаптационного явления и стойком изменении генотипического состава популяции. В этом случае конкретные полезные уклонения отдельных особей превращаются в норму для популяции в целом.
При формировании адаптаций происходит превращение случайного (элементарного адаптационного явления) в необходимое для популяции и вида формирование признаков и свойств. Случайные наследственные изменения в ходе эволюции направленно (и творчески) перерабатываются отбором для создания адаптаций (см. гл. 10).
Это положение Ч. Дарвин подтвердил на многих примерах: формировании яркой окраски цветков у насекомоопыляемых растений, удлинении шеи жирафы, формировании млечных желез млекопитающих и т. д. Так, говоря о возникновении сходства насекомого с каким-либо предметом, он писал: «Если принять, что на долю одного из насекомых выпало некоторое сходство с сухим сучком или опавшим листом и что насекомые изменяются в разных направлениях, то, конечно, все те изменения, которые делают его более похожим на такой предмет и благоприятствуют его сокрытию, будут сохраняться, тогда как другие изменения... совсем исчезнут». Первоначальное приобретение насекомым случайного сходства с каким-либо неживым предметом вовсе «нельзя считать невероятным, принимая во внимание почти бесконечное количество окружающих предметов и разницу в форме и окраске существующих насекомых». Приспособления не возникают в готовом виде, а складываются в процессе многоступенчатого отбора удачных вариантов из множества изменившихся особей в чреде поколений.
В эволюционном смысле понятие «адаптация» должно относиться не столько к отдельной особи, сколько к популяции и виду. Изменения же в пределах отдельной особи в ответ на те или иные изменения окружающей среды происходят в пределах унаследованной каждой особью нормы реакции. Это и изменения ориентации хлоропластов в клетке под влиянием света, и изменения интенсивности транспирации и дыхания растений в течение суток, и т. п. С эволюционной точки зрения адаптацией во всех этих случаях будет способность организмов данного вида меняться в соответствии с действием внешнего фактора, т. е. норма реакций данного признака или свойства. Конкретные же адаптивные проявления реакций в процессе отдельного онтогенеза иногда называются акклимациями, а групповые изменения в пределах нормы реакций вида, сходные у групп особей,— модификациями
Классификация адаптаций
С эволюционной точки зрения важно не простое описание множества различных адаптаций, а классификация их по происхождению, принадлежности к разным аспектам среды, масштабу.
Пути происхождения адаптаций. По происхождению различают преадаптивные, комбинативные и постадаптивные адаптации. В случае преадаптации потенциальные адаптационные явления возникают, опережая существующие условия. Мутационный процесс и скрещивания приводят к накоплению в популяциях скрытого (мобилизационного) резерва наследственной изменчивости. Часть его в будущем может быть использована для создания новых приспособлений (С.М. Гершензон).
Один из примеров превращения ранее существовавших в скрытом виде в генофонде отдельных мутаций в адаптацию был описан выше применительно к явлению индустриального меланизма (см. гл. 10).
При преадаптивном пути возникновения адаптаций нередко с успехом используются прежние особенности организма, возникшие в иных условиях. При этом некоторые сложные приспособления могут возникать, как бы «опережая» условия, при которых эти особенности окажутся адаптациями. Например, наличие швов в черепе млекопитающих облегчает роды, хотя их возникновение не было связано с живорождением.
При возникновении адаптаций комбинативным путем существенно взаимодействие новых мутаций друг с другом и с генотипом в целом. Эффект мутаций зависит от той генотипической среды, в состав которой в будущем они войдут. Скрещивание особей дает разнообразное сочетание мутантного аллеля с другими аллелями того же и других генов. Это приводит к изменению эффекта проявления мутации путем взаимодействия генов. При этом может быть или усиление (комплиментация), или подавление (эпистаз) его выражения в фенотипе; кроме того, обычно мутантный аллель под действием многих генов проявляется градуированно (полимерия). Во всех случаях создается реальная возможность для быстрой смены одних адаптаций другими. Комбинативный путь формирования адаптаций, видимо, наиболее распространенный в природе.
Постадаптивный путь возникновения адаптаций связан с редукцией ранее развитого признака и переводом определяющих его реализацию генов в рецессивное состояние (или использованием ранее существующего органа в других целях — не тех, что определили его появление посредством соответствующего давления отбора). При переводе генов, влияющих на развитие редуцируемых органов, в рецессивное состояние (что весьма вероятно) они включаются в скрытый резерв наследственной изменчивости. Эти гены сохраняются в генофонде популяций и время от времени могут проявляться фенотипически (например, атавизмы, см. гл. 6). В случае установления отбором положительной связи между такими генами и новыми условиями среды они могут дать начало развитию новых признаков и свойств.
При постадаптивном пути новые адаптации возникают посредством использования ранее существовавших структур в случае смены их функций (см. гл. 16). Так, висцеральный скелет у предков позвоночных состоял из жаберных дуг, представленных нерасчлененными кольцами и охватывавших передний конец пищеварительной трубки. Они служили как бы распоркой для пищеварительной трубки, препятствуя ее спадению. Однако в ходе дальнейшей эволюции с усилением функции дыхания жаберные дуги становятся частью системы нагнетания жидкости. В дальнейшей эволюции жаберные дуги принимают на себя функции хватания и превращаются в челюсти.
При классификации адаптаций с нескольких позиций любая адаптация, просмотренная одновременно в свете различных подходов, характеризуется достаточно определенно и четко (табл. 11.1).
| Таблица 11.1. Классификация адаптаций (по Н.В. Тимофееву-Ресовскому и др., 1969) | |
| Принцип классификации | Группа адаптаций |
| По происхождению | Возникающие преадаптивным, комбинативным и постадаптивным путями |
| По принадлежности к разной среде | Генотипические (онтогенетические), популяционно-видовые, биоценотические |
| По эволюционному масштабу | Специализированные и общие |
| По характеру возникающих изменений | Упрощающие строение системы, усложняющие строение, сохраняющие строения систем и уровень сложности |
| По длительности сохранения в онтогенезе | Кратковременные, повторяющиеся и постоянные |
Получение такой четкой и определенной характеристики адаптации может показаться имеющим сугубо теоретическое значение. Но, как мы неоднократно подчеркивали, эволюционная теория в обозримом будущем должна стать основой сознательного существования человечества в биосфере, основой направленной переделки и учета возможных последствий вторжения человека в планетарные процессы. И при этом проблема возникновения, формирования, трансформации адаптации живых организмов приобретает неизмеримо большее значение, чем то, которое она сейчас имеет в «биологических» отраслях хозяйства (сельское хозяйство, микробиология, промысловое хозяйство и т. п.).
Адаптации в разных средах. По принадлежности к аспектам среды адаптации бывают различными. Любой результат естественного отбора связан с тем или иным изменением биотической среды, которая в соответствии с уровнями организации живого
(см. гл. 4) может быть подразделена на генотипическую, онтогенетическую, популяционно-видовую и биоценотическую. Подразделения среды отличаются и специфическими адаптациями.
Для генотипической среды характерны целостность генотипа особи и взаимодействие генов между собой. Целостность генотипа определяет особенности доминирования генов и развитие коадаптаций. На молекулярном уровне мы встречаем тонкую адаптивную организацию строения и взаимодействия молекул, обеспечивающих эффективное воспроизведение и самоконструирование биополимеров. Возникает вопрос: все ли особенности строения биополимеров оказываются адаптивными? С точки зрения генетического кодирования ясно, что не все, поскольку существует явление вырожденности генетического кода (см. далее гл. 20, раздел 1). Однако должны ли мы признать за явлениями на молекулярном уровне организации жизни только функции генетического кодирования? Не слишком ли мы мало знаем для того, чтобы уверенно говорить об отсутствии других функций у кодонов, скажем УЦУ и УЦЦ, кодирующих одну и ту же аминокислоту серии?
На клеточном уровне исследования мы обнаруживаем многочисленные органеллы со сложной структурой и множеством функций, определяющих слаженный метаболизм клетки и ее функционирование как целого.
Адаптации на уровне отдельной особи связаны с онтогенезом — упорядоченными во времени и пространстве процессами реализации наследственной информации, наследственным осуществлением морфогенеза. Здесь, как, впрочем, и на других уровнях, мы встречаем коадаптации — взаимные приспособления. Например, лопатка и тазовая кость подвижно сочленены с головкой плечевой и бедренной костей. Кости, подвижно прикрепленные друг к другу, имеют взаимные приспособления для обеспечения нормальной работы. В основе коадаптации лежат различные корреляции, которые и регулируют онтогенетические дифференцировки.
На онтогенетическом уровне разнообразны комплексные адаптации физиолого-биохимического характера. В условиях повышенной температуры и недостатка воды нормализация жизнедеятельности растений достигается накоплением в клетках осмотически активных веществ, закрытием устьиц. Повреждающее влияние солей на сильно засоленных почвах в некоторой степени может нейтрализоваться накоплением специфических белков, усилением синтеза органических кислот и т. д.
Популяционно-видовая среда проявляется во взаимодействии особей в пределах популяций и вида в целом. Популяционной среде соответствуют надорганизменные, популяционно-видовые адаптации. К популяционно-видовым адаптациям относятся, например, половой процесс, гетерозиготность, мобилизационный резерв наследственной изменчивости, определенная плотность популяций и т. д. Для обозначения ряда специальных внутривидовых адаптаций существует термин «конгруэнции» (С.А. Северцов). Конгруэнции — взаимоприспособления особей, возникающие в результате внутривидовых отношений. Они выражаются в соответствии строения и функции органов матери и детеныша, аппаратов размножения самцов и самок, в наличии приспособлений для отыскания особей противоположного пола, системы сигнализации и разделения труда между особями в стадах, колониях, семьях и т. д.
Чрезвычайно разнообразны способы взаимодействия видов в биогеоценозах. Растения воздействуют друг на друга через изменения не только условий освещенности и влажности, но и выделяя специальные активные вещества, способствующие вытеснению одних и размножению других видов (аллелопатия).
Строго разграничить генотипические, онтогенетические, популяционные и биоценотические адаптации практически трудно. Адаптации, относящиеся к одной из сред «срабатывают» и на других средах; все адаптации подчиняются принципу мультифункциональности (см. гл. 16). Это и понятно, так как разные эволюционные среды (генотипическая, популяционная и биогеоценотическая) тесно и неразрывно связаны между собой: особи существуют только в популяциях, популяции населяют конкретные ценозы. Видовой состав биоценоза, определяя характер межвидовых отношений, оказывает влияние и на генотипическую, и на популяционную среды. Действие естественного отбора на популяции приводит к изменениям и в биоценотической среде, меняя характер межвидовых отношений.
Масштаб адаптаций. По масштабу адаптации делятся на специализированные, пригодные в узколокальных условиях жизни вида (например, строение языка у муравьедов в связи с питанием муравьями, приспособления хамелеона к древесному образу жизни и т. п.), и общие, пригодные в широком спектре условий среды и характерные для больших таксонов. К последней группе относятся, например, крупные изменения в кровеносной, дыхательной и нервной системах у позвоночных, механизмы фотосинтеза и аэробного дыхания, семенное размножение и редукция гаметофита у высших растений, обеспечивающие проникновение их в новые адаптивные зоны. Первоначально общие адаптации возникают как специализированные, они смогут выводить определенные виды на путь широкой адаптивной радиации, на путь арогенеза (см. гл. 15). Перспективные общие адаптации обычно затрагивают не одну, а многие системы органов.
Существуют и другие подходы к классификации адаптаций. Так, по характеру изменений адаптации бывают связаны с усложнением или упрощением морфофизиологической организации. Например, паразитизм обычно ведет к упрощению и редукции ряда органов. В то же время переход к активному образу жизни связан с развитием и дифференциацией ряда важных органов нападения и защиты. Адаптации, связанные с выработкой социального, общественного образа жизни у высших беспозвоночных и позвоночных,— более сложные приобретения, чем адаптации микроорганизмов и растений.
Подобно различиям по эволюционному масштабу, адаптации могут отличаться и по онтогенетическому масштабу (длительность сохранения в онтогенезе). Одни адаптации в онтогенезе имеют кратковременное значение, другие сохраняются и на более длительный срок. Одни ограничиваются зародышевыми стадиями развития (см. гл. 14), другие носят повторяющийся характер (сезонные изменения в окраске у животных и растений, различного рода модификаций и т. п.), третьи имеют постоянное значение в жизни особи (строение жизненно важных систем и органов). Изучение адаптаций, отличающихся по приуроченности к разным этапам онтогенеза, важно для понимания эволюции онтогенеза.
9. Основные свойства живого.
Основные свойства живого
Когда-то считалось, что живое можно отличить от неживого по таким свойствам, как обмен веществ, подвижность, раздражимость, рост, размножение, приспособляемость. Но анализ показал, что порознь все эти свойства встречаются и среди неживой природы, а поэтому не могут рассматриваться как специфические свойства живого. Как это ни странно, но до сих пор нет удовлетворяющего всех определения понятия «жизнь, живое».
Пять аксиом теоретической биологии. В одной из последних и наиболее удачных попыток живое характеризуется следующими особенностями, сформулированными Б.M. Медниковым (1982) в виде аксиом теоретической биологии:
1. Все живые организмы оказываются единством фенотипа и программы для его построения (генотипа), передающейся по наследству из поколения в поколение (аксиома А. Вейсмана).
2. Генетическая программа образуется матричным путем. В качестве матрицы, на которой строится ген будущего поколения, используется ген предшествующего поколения (аксиома И.К. Кольцова).
3. В процессе передачи из поколения в поколение генетические программы в результате различных причин изменяются случайно и ненаправленно, и лишь случайно такие изменения могут оказаться удачными в данной среде (1-я аксиома Ч. Дарвина).
4. Случайные изменения генетических программ при становлении фенотипа многократно усиливаются (аксиома Н.В. Тимофеева-Ресовского).
5. Многократно усиленные изменения генетических программ подвергаются отбору условиями внешней среды (2-я аксиома Ч. Дарвина).
Из перечисленных аксиом можно вывести, по-видимому, все основные свойства живой природы. Рассмотрим лишь некоторые свойства, имеющие прямое отношение к процессу эволюционного развития.
Дискретность и целостность – два фундаментальных свойства организации жизни на Земле. Живые объекты в природе относительно обособлены друг от друга (особи, популяции, виды, экосистемы и т.п.). Любая особь многоклеточного животного состоит из клеток, а любая клетка и одноклеточные существа – из определенных органелл. Органеллы состоят из дискретных, обычно высокомолекулярных, органических веществ, которые, в свою очередь, состоят из дискретных атомов, элементарных (тоже дискретных!) частиц. В то же время сложная организация немыслима без взаимодействия ее частей и структур – без целостности. Целостность биологических систем качественно отличается от целостности неживого прежде всего тем, что целостность живого поддерживается в процессе развития. Живые системы – открытые системы, они постоянно обмениваются веществами и энергией со средой. Для них характерна отрицательная энтропия (возрастание упорядоченности), увеличивающаяся, видимо, в процессе органической эволюции. Вероятно, что в живом проявляется способность к самоорганизации материи.
Среди живых систем нет двух одинаковых особей, популяций и видов. Эта уникальность проявления дискретности и целостности живого основана на замечательном явлении конвариантной редупликации.
Конвариантная редупликация (самовоспроизведение с изменениями), осуществляемая на основе матричного принципа (сумма трех первых аксиом),– это, видимо, единственное специфическое для жизни (в известной нам форме ее существования на Земле) свойство. В основе его лежит уникальная способность к самовоспроизведению основных управляющих систем (ДНК, хромосом и генов). Редупликация определяется матричным принципом (аксиома Кольцова) синтеза макромолекул (рис. 4.1). «Размножение» и рост кристаллов принципиально отличаются от такой редупликации по матричному принципу, не обеспечивая появления практически бесконечного числа новообразований (1-я аксиома Дарвина), передающихся по наследству. Благодаря способности к самовоспроизведению по матричному принципу молекулы ДНК смогли выполнить роль носителя наследственности исходных управляющих систем (аксиома Вейсмана). Это свойство нуклеиновых кислот – важная предпосылка эволюции жизни.
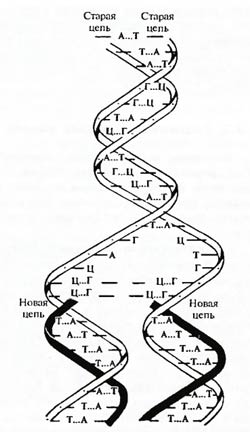
Рис. 4.1. Схемы редупликации ДНК. Процесс связан с разделением пар оснований аденин – тимин (А–Т) и гуанин – цитозин (Г–Ц) и раскручиванием двух цепей исходной спирали. Каждая цепь используется как матрица для синтеза новой цепи (по Дж. Севейджу, 1969).
Исходные управляющие системы, молекулы ДНК примитивных дискретных живых частиц (вирусы, фаги, бактерии и сходные формы), свободно размножающихся клеток-эукариот, половых клеток многоклеточных организмов обладают относительно высокой степенью стабильности, что обеспечивает возможность идентичного самовоспроизведения (явление наследственности). Эволюционный процесс немыслим без наследственной передачи свойств в ряду поколений.
Подчеркнем еще раз, что при самовоспроизведении управляющих систем в живых организмах происходит не механическое повторение, а воспроизведение с внесением изменений (1-я аксиома Ч. Дарвина). Неизбежность таких изменений вытекает из физико-химических свойств молекул ДНК. Любая достаточно сложная молекулярная и сверхмолекулярная структура обладает ограниченной степенью стабильности. Время от времени она претерпевает структурные изменения в результате движения атомов и молекул. Эти изменения, если не ведут к летальному исходу, будут многократно усиливаться (аксиома Тимофеева-Ресовского) и затем передаваться по наследству в результате самовоспроизведения по матричному принципу. Конвариантная редупликация означает возможность передачи по наследству дискретных отклонений от исходного состояния, т.е. мутаций.
10. Темпы эволюции групп. Вымирание групп и его причины.
Темпы эволюции групп
Эволюция как грандиозный процесс изменения органического мира Земли характеризуется определенными временными параметрами. Скорость (темп) — одна из самых важных особенностей процесса эволюции в целом. Выяснение конкретной скорости эволюционного изменения групп часто важно и для решения чисто практических задач, например, связанных с возникновением и распространением новых болезнетворных микроорганизмов, появлением резистентности насекомых к инсектицидам и др.
В эволюции большее значение имеет не абсолютное, астрономическое, время, а число поколений. Причины этого будут понятны, если вспомнить, что механизм возникновения адаптаций сводится, по существу, к изменению концентрации аллелей в популяции в чреде поколений (см. гл. 11). Однако для многих ископаемых форм мы не можем с достаточной точностью оценивать скорость их эволюции числом поколений. Поэтому, несмотря на некорректность астрономического времени для характеристики темпов эволюции, приходится пользоваться и этим параметром.
Существует и другой подход для определения темпа эволюции группы — учет скорости смены видов (точнее — фратрий) в одном филогенетическом ряду (Дж. Симпсон).
Проблема выбора критериев скорости эволюции. Определить скорость эволюции не всегда просто. При рассмотрении темпов эволюции часто имеют в виду изменение не только целых групп, но и отдельных признаков. Хотя изменения групп основаны на онтогенетических дифференцировках (на появлении и развитии отдельных признаков в онтогенезе особей, см. гл. 14), все же эти проблемы различны; проблема скорости изменения отдельных признаков должна рассматриваться вместе с другими особенностями эволюции органов и функций (см. гл. 16).
Теоретически скорость эволюции вида можно определить по темпу его изменения в ходе филетической эволюции. Но как измерить этот темп? Вероятно, таким мерилом могло бы быть число элементарных эволюционных явлений (см. гл. 8) в расчете на поколение, а затем число возникших за определенное время (число поколений) адаптаций. Однако пока практически определить названные параметры вряд ли возможно для любой группы. Гораздо удобнее определить темп эволюции группы по числу возникающих видов за определенное время.
Темпы формообразования. Материал, накопленный в настоящее время в ботанике и зоологии, позволяет установить два разных по времени способа видообразования: «внезапный» и «постепенный». Рассмотрим несколько примеров.
Внезапное видообразование связано с быстрой перестройкой всего генома, например при полиплоидии. Известно, что возникновение полиплоидного организма происходит в течение считанных минут. Возникшая полиплоидная особь генетически изолирована от всех остальных особей вида. Часто полиплоидные особи оказываются более устойчивыми по отношению к действию экстремальных факторов внешней среды и получают широкое распространение в условиях, где родительская форма не выживает. Таким образом, новые особи, репродуктивно изолированные от остальных особей в популяции, могут возникнуть за одно поколение. Такой процесс мог лежать в основе возникновения полиплоидных видов в разных группах. Полиплоидные ряды видов известны для многих растений (см. рис. 6.28 и 13.5). Картофель, белый клевер, люцерна, тимофеевка и другие полиплоидные растения возникли таким путем.
Другой тип полиплоидии связан с гибридизацией (аллополиплоидия), объединением геномов сравнительно близких видов (см. гл. 6 и 13). Г.Д. Карпеченко путем гибридизации капусты и редьки была синтезирована новая форма — редько-капуста (Raphanobrassica) (рис. 15.10). Б.Л. Астаурову удалось получить аллодиплоид двух видов шелкопряда: Bombux тоri и В. mandarina.

Рис. 15.10. Схема синтеза формы видового ранга — редько-капусты; А — стручок редьки Raphanus sativus и ее хромосомный набор (18R); Б — стручок капусты Brassica oleracea и ее хромосомный набор (18B); В — первичный, полностью стерильный из-за нарушения мейоза гибрид с хромосомным набором 9R + 9B; Г— аллотетраплоид с хромосомным набором 18R + 18B вполне плодовитый, так как в мейозе 18 хромосом R конъюгируют друг с другом и образуют 9 бивалентов, как и 18 хромосом В (по Г.Д. Карпеченко, 1935)
Наконец, причиной быстрого изменения генома, определяющего генетическую изоляцию вновь возникшей особи от исходной формы, могут быть перестройки хромосом (фрагментация и слияние). У растений возможность участия межхромосомных перестроек в эволюции была экспериментально подтверждена созданием скерды (Crepis nova), отличающейся по морфологии хромосом от родительского вида. У животных роль хромосомных перестроек в видообразовании была показана в обширной серии экспериментальных и популяционно-генетических работ в группе видов дрозофилы Drosophila persimilis — virilis (см. гл. 6, рис. 6.27).
Следовательно, в природе возможно быстрое (практически внезапное) образование особи, репродуктивно изолированной от остальных особей того же поколения. В некоторых редких случаях такие особи оказываются способными к размножению и на протяжении последующих поколений могут дать начало целой группе подобных особей. Естественно, что одна-единственная особь (или даже несколько особей) видом названа быть не может уже хотя бы потому, что всякий вид — это всегда генетическая система с множеством свойств, присущих именно системе особей, в том числе с собственным ареалом, многомерной экологической нишей и т. д. Получается, что в природе могут существовать особи, уже не относящиеся к старому виду и еще не образовавшие нового вида.
Решение этой проблемы нашел К.М. Завадский, предложивший называть такие новые формы не видами, а формами видового ранга. Ясно, что из многих таких «кандидатов» в новые виды лишь небольшое число действительно превратится в самостоятельный вид. Для этого они должны оказаться достаточно жизнеспособными и иметь преимущества перед родительскими формами, позволяющими им распространиться и занять определенный ареал. Такое превращение формы видового ранга (представленной сначала немногими особями, а потом отдельными мелкими группами) в реальный вид должно продолжаться значительное число поколений и осуществляется всегда под контролем естественного отбора.
Более обычно в эволюции постепенное формообразование. Сейчас накопилось много данных, позволяющих сравнительно точно определять скорость образования подвидов у самых разных групп. Многие из них касаются послеледниковых формообразований, когда мы, зная время освобождения от ледника тех или иных местностей, можем надежно оценивать период времени, в пределах которого должно было произойти образование подвидов, обитающих сегодня в этих местах (табл. 15.1).
| Таблица 15.1. Примеры скоростей образования новых подвидов | ||
| Вид | Район образования нового подвида | Период, лет |
| Благородный олень (Cervus elaphus) | Великобритания | Около 8 тыс. |
| Ладожская нерпа (Pusa hispida lagopensis) | Оз. Ладога | » 9 тыс. |
| Сайменская нерпа (Pusa hispida saimensi) | Оз. Сайма, Финляндия | » 9 тыс. |
| Хохлатый жаворонок (Galerida cristata) | Низовья р. Нил, Египет | » 8—10 тыс. |
| Сизый голубь (Columba livia) | Оазис Дакла. Северная Африка | » 8—10 |
| Варакушка (Cyanosylvia svecica) | Норвегия | » 8—10 |
| Обыкновенный тюлень (Phoca vitulina) | Северная Канада | » 4 тыс. |
| Европейский кролик (Oryctolagus cuniculus) | О-ва у Великобритании | » 500 |
| Домовая мышь (Mus musculus) | Фарерские о-ва | Не более 3 тыс. |
| 10 подвидов млекопитающих | О. Ньюфаундленд | Не более 12 тыс. |
| 4 подвида рыб | О. Айл-Роял на оз. Верхнее | 12—25 тыс. |
Можно видеть, что подвид часто образуется за несколько тысяч лет, а иногда за сотни или сотни тысяч лет. Полнота палеонтологического материала по отдельным группам в настоящее время настолько велика, что позволяет с уверенностью определять время, необходимое для возникновения новых видов. Особенно удобен для этого анализ последовательных ископаемых форм (см. рис. 6.4; 6.5). Расчеты показывают, что для образования отдельных видов (фратрий) верхнеплиоценовых моллюсков на территории Германии потребовалось несколько миллионов лет, тогда как в Черном море в четвертичном периоде новый вид рода Paphia возник всего примерно за 6000 лет. Один вид зубра (Bison) на Кавказе в среднем плейстоцене существовал примерно 50—60 тыс. поколений, а переход к новому виду занял 10—16 тыс. лет (2—4 тыс. поколений). Оказалось также, что время существования отдельных видов может значительно колебаться — от 0,5 до 5 млн лет, а время существования родов значительно различается, например, у моллюсков и хищных млекопитающих. У моллюсков средняя длительность существования родов — десятки миллионов лет, у хищных — около 8 млн лет. Это означает, что темп формообразования у хищных в десяток раз быстрее, чем у двустворчатых моллюсков. При этом надо учесть, что определяемая таким образом скорость эволюции несколько занижена, так как время видообразования может быть существенно короче, чем время дальнейшего существования вида. Поскольку на палеонтологическом материале надежнее выделяются не виды, а роды и более крупные таксономические единицы, возникает возможность определения скоростей эволюции по числу образованных в единицу времени родов и семейств. На рис. 15.11 приведены такие данные для шестилучевых кораллов на протяжении 550 млн лет — с триаса до неогена. Видно, что в разные геологические периоды скорость эволюции менялась в 3—4 раза: в триасе и юре I семейство кораллов возникало за 5—5,5 млн лет, и за это же время возникало 5—12 родов, то в палеогене — неогена для возникновения семейства требовалось 17—20 млн лет, и на каждое семейство приходилось 40—67 родов. Это позволяет предположить об уменьшении темпов эрогенной эволюции, при увеличении темпов аллогенной эволюции (адаптивной радиации). Конечно, в других группах скорости эволюции могут быть иными.

Рис. 15.11. Определение скорости эволюции по числу возникающих новых родов (I) и семейств (II) шестилучевых кораллов Scleractinia (из Д. Раупа и С. Стенли, 1974)
Сравнение большого материала по темпам возникновения и длительности развития новых родов позволило выделить три группы родов (рис. 15.12). К первой группе принадлежат брадителические роды, филогенетические реликты (см. ниже); время их развития превышает 250 млн лет. Ко второй группе принадлежат горотелические роды, время развития которых составляет от нескольких до нескольких десятков миллионов лет, и, наконец, к третьей группе — тахителических родов — принадлежат роды, время развития которых невелико, много меньше I млн лет (Дж. Г. Симпсон).
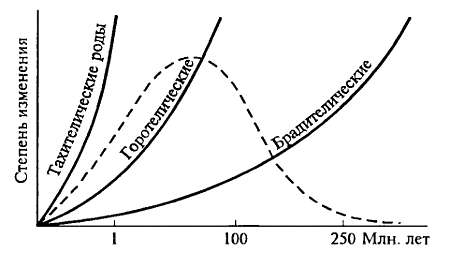
Рис. 15.12. Распределение скоростей эволюции. Три группы родов выделены по темпам возникновения и длительности существования. Большая часть всех изученных родов оказывается горотелическими, а суммарно все роды вместе дают картину, близкую к нормальному распределению (ориг.)
Можно сказать, что ни мутационный процесс, ни волны жизни, видимо, не оказывают решающего влияния на скорость протекания процесса эволюции в любой группе. Но очень велико влияние изоляции и особенно естественного отбора. При определении скорости эволюционного процесса изоляция действует не только как элементарный эволюционный фактор, но и как фактор, активно способствующий перестройке всех биогеоценозов и созданию условий для дальнейшей эволюции. Однако главнейшим фактором, определяющим как продолжительность существования того или иного филума, так и скорость образования новых форм, является естественный отбор.
Успехи молекулярной биологии дали возможность оценить скорость эволюции по замещениям аминокислот в белках и нуклеотидов в нуклеиновых кислотах. Для выражения скорости таких замен принята единица «полинг», равная 10—9 замене на аминокислотный сайт в год (см. гл. 6).
Сравнение аминокислотной последовательности в α- и β-цепях гемоглобина, в молекулах глобулина, цитохрома и других белков показало увеличение числа замен по мере удаления филогенетического расстояния у позвоночных. У всех сравниваемых позвоночных скорость замены для одного и того же белка выражалась сходной величиной в полингах. Разные же белки в пределах позвоночных обычно отличаются по своей скорости эволюции (табл. 15.2). Теперь молекулярно-генетические расчеты позволяют строить филогенетические древа организмов, близкие к морфологическим древам. Скорость молекулярной эволюции (в полингах) не зависит от продолжительности жизни поколения и величины популяций. Постоянство темпов молекулярной эволюции пытаются объяснить фиксацией нейтральных мутаций. Так, на примере α-цепи гемоглобина при сравнении различных позвоночных установлено, что за 7 млн лет происходит замена одной аминокислоты. Данные, полученные по разным белкам, положены в основу идеи молекулярных часов эволюции (см. гл. 6 и 20).
| Таблица 15.2. Скорость эволюции белков у млекопитающих (М. Кимура, 1985) | |
| Белки | Kaa, 10—9 в год |
| Фибринопептиды | 8,3 |
| Панкреатическая рибонуклеаза | 2,1 |
| Лизоцим | 2,0 |
| α-Цепь гемоглобина | 1,2 |
| Миоглобин | 0,89 |
| Инсулин | 0,44 |
| Цитохром C | 0,3 |
| Гистон H4 | 0,01 |
Kaa — среднее число аминокислотных замен, приходящееся на пару гомологичных сайтов двух сравниваемых полипептидов.
Дата добавления: 2021-06-02; просмотров: 461; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!