Взаимодействие неаллельных генов
Тема 4.2 Взаимодействие генов (аллельных и неаллельных)
1. Взаимодействие аллельных генов (полное и неполное доминирование).
2. Взаимодействие неаллельных генов (комплементарность, полимерия, эпистаз).
3. Плейотропное действие генов.
4. Пенентрантность и экспрессивность.
Взаимодействие аллельных генов
Аллель – одна из двух или более альтернативных форм гена, каждая из которых характеризуется уникальной последовательностью нуклеотидов. Аллельные гены находятся в одинаковых локусах (участках) гомологичных хромосом и отвечают за развитие одинаковых признаков.
1.1 Полное доминирование
В лекции 6 были рассмотрены схемы, когда доминантный ген полностью подавлял рецессивный (полное доминирование). Примером полного доминирования у человека является наследование резус-фактора.
В 1940 г. Ландштейнер и Винер обнаружили резус-фактор (Rh), антиген, содержащийся в крови человека и макака-резуса. В том же году Левин и Стентсон исследовали сыворотку крови женщины, которая родила мертвый плод и в анамнезе которой имело место переливание крови мужа, совместимой по АВ0 группе. При этом ими были обнаружены особые антитела – белок (резус-фактор). Резус-фактор – это антигенный белок липопротеид, расположенный на мембранах эритроцитов у 85% людей. Именно по наличию или отсутствию антигена D традиционно выделяют резус-положительных (Rh+) и резус-отрицательных (Rh-) людей. В настоящее время известны три пары неаллельных генов, расположенных близко друг к другу и контролирующих фенотипическое проявление резус-фактора. Однако для определения вероятности наследования часто бывает достаточно расценивать резус-фактор как простой менделирующий признак (наследующийся по законам Менделя, за который отвечает пара аллельных генов. Тогда человек с резус-отрицательной кровью будет гомозиготой по рецессивному аллелю (dd). А резус-положительный человек может быть как гомозиготой по доминантному алелю (DD), так и гетерозиготой (Dd).
|
|
|
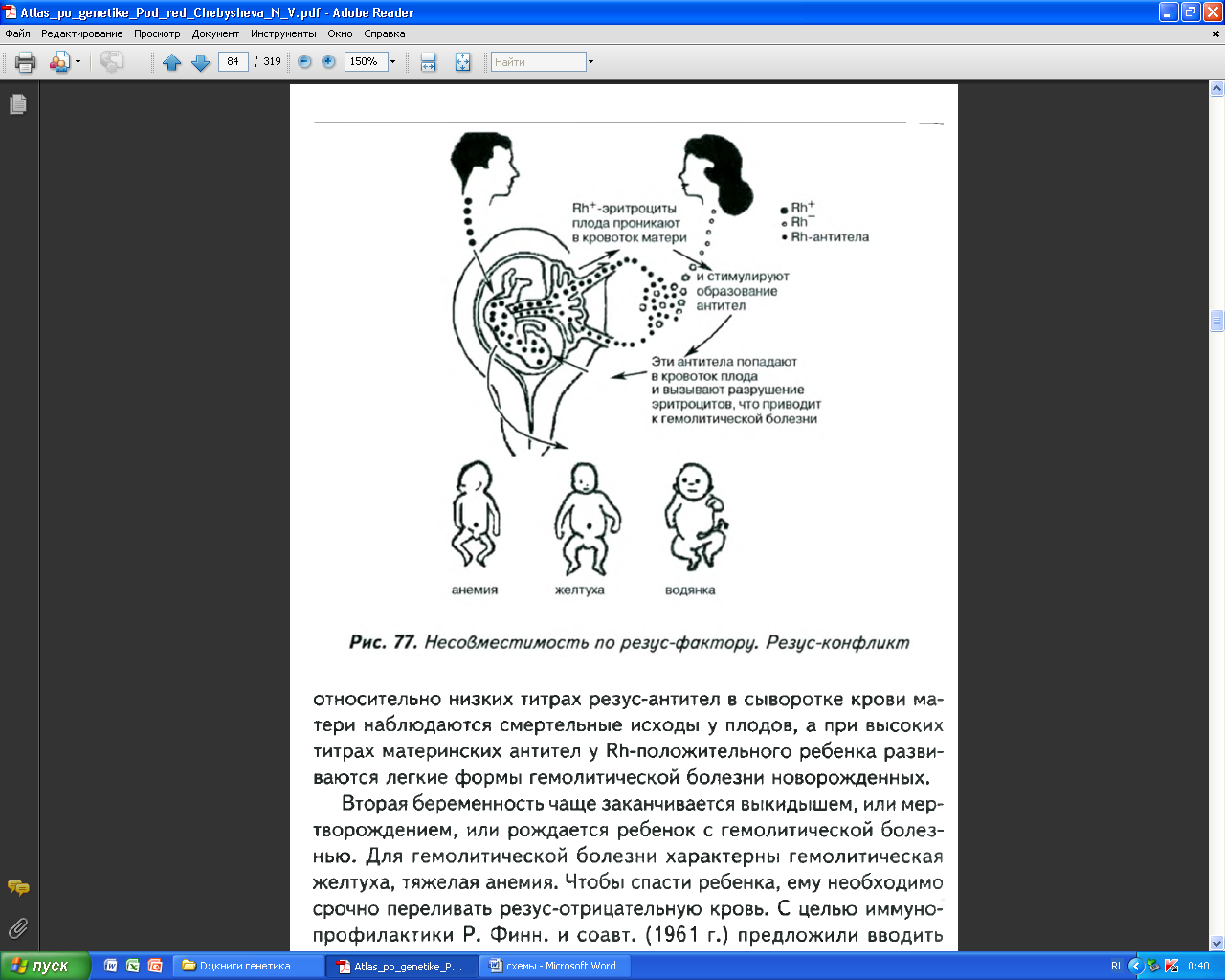
Огромная практическая ценность системы резус стала очевидной, когда была установлена связь между этими антителами и несчастным случаями при переливании крови. Кроме того, было показано, что именно резус – несовместимость матери и плода является причиной иммунного конфликта, приводящего к эритробластозу плода и гемолитической болезни новорожденных. Иммунный конфликт возникает при вынашивании резус-отрицательной матерью положительного плода.
Эритробластоз – это болезнь плода и новорожденного, в 90% случаев вызванная иммунным конфликтом по резус-фактору: в случае, если у резус-отрицательной матери развивается резус-положительный плод. В период беременности, когда у плода начинает вырабатываться в крови резус-положительный белок, в организме матери начинают вырабатываться резус-антитела, которые, накапливаясь, губительно действуют на плод, вызывая у него агглютинацию эритроцитов. За первую беременность, как правило, антитела еще не успевают накопиться в критическом для плода количестве, но в последующие беременности иммунный конфликт приводит или к патологиям плода, или к рождению детей, страдающих анемией, гемолитической болезнью, водянкой или другими патологиями. Иногда такие дети отстают в развитии и имеют ослабленное здоровье. При этом резус-антигены связываются, так что в ее организме уровень антител на положительные антигены снижается, и вред для здоровья следующего ребенка уменьшается.
|
|
|
Основным механизмом патогенеза всех форм этого заболевания является гемолиз (интенсивное разрушение форменных элементов крови). Именно поэтому эритробластоз называют гемолитической болезнью новорожденных, которая проявляется прежде всего в виде анемии, желтухи, водянки. Это заболевание нередко заканчивается летально, может сопровождаться поражением ЦНС, что приводит к спастическим параличам и умственной и физической осталости.
|
|
|
У 85% людей содержится в крови этот фактор (резус-положительные). Резус-несовместимость встречается в 10% от общего числа родов, а частота эритробластозов 1/25 иммунных конфликтов. Основной способ лечения эритробластозов – удаление антител из крови больного ребенка и замещение крови. Это достигается обменным переливанием крови сразу после рождения.
Резус-фактор наследуется как аутосомно-доминантный менделирующий признак.
1.2 Неполное доминирование
Встречаются признаки, которые в случае гетерозиготного сочетания аллелей будут приводить к промежуточному проявлению признаков по фенотипу. Такое наследование называют неполным доминированием. При этом доминантный ген не полностью подавляет рецессивный.
Например: В брак вступает курчавая женщина (доминантный признак) и мужчина с прямыми волосами (рецессивный признак). У них родились дети с волнистыми волосами. Какой прогноз в отношении внуков, если дети вступят в брак с людьми, имеющими волнистые волосы.
Схема решения:
А – курчавые волосы.
а- прямые волосы.
Аа – волнистые волосы.
Р АА х аа
G А а
F1 Аа (курч.)
F2
| м ж | А | а |
| А | АА (курч.) | Аа (волн.) |
| а | Аа (волн.) | Аа (прям.) |
|
|
|
Таким образом, получились следующие вероятности возможных фенотипов по этому признаку у внуков: 1АА – курчавые, 2Аа – волнистые, 1аа – прямые.
Другой пример – наследственное заболевание человека – синдром Тея-Сакса. Гомозиготы по рецессивному (мутантному) аллелю страдают тяжелым нарушением липидного обмена и умирают в течение первых трех лет жизни. Эти нарушения обусловлены дефектность фермента гексоаминидазы, участвующего в обмене липидов. У гетерозигот имеется лишь одна копия мутантного гена, и фенотипически они нормальны, однако активность фермента у них снижена наполовину, он достаточен для нормального метаболизма.
Кроме того, по принципу неполного доминирования у человека наследуются размер носа, рта, глаз, расстояние между глазами.
1.3 Кодоминирование
Кодоминирование – это тип взаимодействия аллельных генов, при котором в гетерозиготном состоянии оказываются и работают вместе два доминантных гена одновременно, то есть каждый аллель определяет свой признак.
В диплоидном организме каждый признак контролируется одним геном, который представлен одной из двух аллельных форм. Известны примеры, когда один признак проявляется в нескольких различных формах, контролируемых тремя и более аллелями. В таких случаях говорят о множественных аллелях – разновидностях одного гена, отвечающих за фенотипическое проявление одного признака.
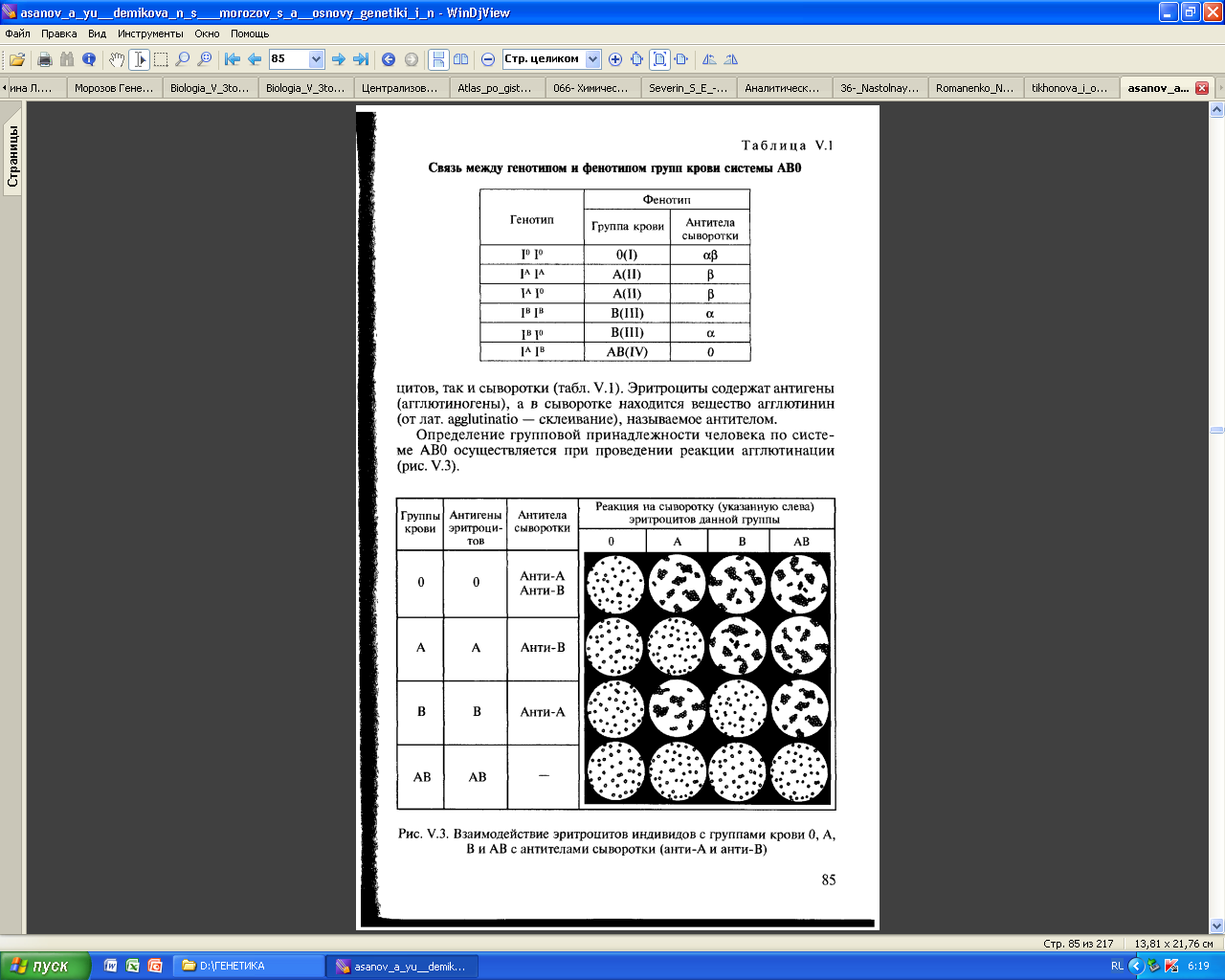
Наследование системы групп крови АВО человека контролируется множественными аллелями. Группа крови контролируется аутосомным геном. Локус этого гена обозначают буквой I от слова «изогемагглютиноген», а три его аллеля буквами А, В и О. Аллели IА и IВ доминантны в одинаковой степени, а аллель IО рецессивен по отношению к ним обоим, и наследуются они как аутосомные менделирующие признаки.
Группы крови определяются антигенами, расположенными на эритроцитах. Антигены – высокомолекулярные вещества, характеризующие индивидуальную принадлежность тканей и располагающиеся на поверхности клеток. Попадание в организм чужеродных антигенов провоцирует начало иммунологического процесса, выработку антител.
Система групп крови АВ0.
Первая АВ0 - система групп крови была открыта Ланштейнером в 1900 г. В 1911 г. было показано, что группы крови наследуются. В 1924 г. Бернштейн установил, что система групп крови АВ0 контролируется серией множественных аллелей (А, В, 0) одного локуса гена у человека. Любой индивид может иметь максимум 2 аллеля из серии.
Дифференцировка крови по системе АВ0 на 4 группы основана на комбинации двух изоантигенов (А и В) в эритроцитах и двух антител (α и β) – в плазме крови. Это необходимо учитывать при переливании крови.
1. Группа 0 (I) – в эритроцитах нет агглютиногенов А и В на эритроцитах, имеются агглютинины α и β.
2. Группа А (II) – на эритроцитах содержится агглютиноген А, в сыворотке – агглютинин бета.
3. Группа В (III) – на эритроцитах содержится агглютиноген В, в сыворотке – агглютинин альфа.
4. Группа АВ (IV) – на эритроцитах содержатся агглютиногены А и В, в сыворотке агглютининов нет.
Система АВО определяется тремя аллельными генами, которые расположены на длинном плече 9 хромосомы и обозначаются IА , IВ , IО. Первая группа крови – это гомозигота по аллелю IО (IО IО). Вторая группа крови обнаруживается как при гомозиготности гена IА (IА IА), так и при гетерозиготности (IА IО). Третья группа также возникает в двух вариантах: IВ IВ и IВ IО. Четвертая группа крови формируется при одновременном наличии в организме двух доминантных аллелей: IА IВ.
1.4 Сверхдоминирование
Сверхдоминирование – лучшая приспособленность к среде гетерозигот по сравнению с обоими типами гомозигот. Пример: взаимоотношения между нормальным (S) и мутантным (s) аллелями гена, контролирующего структуру гемоглобина человека. Люди, гомозиготные по мутантной аллели (ss), страдают тяжелым заболеванием крови – серповидноклеточной анемией, от которой они гибнут обычно в детском возрасте. Однако в тропической Африке и других районах, где распространена малярия, в популяции человека постоянно присутствуют все три генотипа – SS, Ss, ss. Гетерозиготы составляют до 40%. Оказалось, что сохранение в популяции летальной аллели s обусловлено тем, что гетерозиготы более устойчивы к малярии, чем гомозиготы по нормальному гену.
1.5 Аллельное исключение
К разновидностям внутриаллельного взаимодействия генов относится и аллельное исключение, когда у гетерозиготного организма в одних клетках активна одна аллель, а в других — другая. Например, у человека и млекопитающих каждая плазматическая клетка синтезирует только одну (свою) цепь иммуноглобулинов (антител). Другим примером аллельного исключения является инактивация одной из двух Х-хромосом у женского организма. Случайный характер инактивации приводит к выключению из функции в одних клетках материнской Х-хромосомы, в других — отцовской.
Взаимодействие неаллельных генов
Зачастую признаки формируются при участии нескольких генов, взаимодействие между которыми отражается на проявлении фенотипа.
2.1 Комплементарность
Комплементарное взаимодействие генов – явление, при котором два неаллельных гена, находясь одновременно в генотипе, приводят к формированию нового фенотипического признака.
Комплементарность бывает доминантной (когда комплементирующие гены доминантны) и рецессивной (когда они рецессивны). Но во всех случаях, когда гены расположены в разных парах хромосом, в основе расщеплений лежат цифровые законы, установленные Менделем.
Например, чтобы человек имел нормальный слух, необходима согласованная деятельность нескольких пар генов, каждый из которых может быть представлен доминантными и рецессивными аллелями. У человека слух нормален, если каждый из них имеет хотя бы по одному доминантному аллелю в диплоидном наборе хромосом. В случае, если хотя бы один из них представлен рецессивной гомозиготой, человек будет глухим. Рассмотрим пример: предположим, что нормальный слух формирует пара генов. Ген А кодирует развитие улитки, ген В отвечает за развитие слухового нерва.
В брак вступают пара глухих, у них рождаются дети с нормальным слухом. Это может быть только в случае, если один родитель страдает глухотой по гену «в» (ААвв), а другой – по гену «а» (ааВВ).
Р: ААвв (глух.) х ааВВ (глухой)
Гаметы: Ав аВ
F1: АаВв (норм. слух).
Все дети получат с гаметами аВ и Ав доминантные аллели, их генотипы будут одинаковы – АаВв, т.е. каждая гамета будет иметь доминантный аллель по каждому гену, которые будут взаимодействовать, дополняя друг друга и формировать новый, по отношению к родителям, признак – нормальный слух. Это и есть комплементарное взаимодействие генов.
Допустим, что дети вступят в брак с людьми с такими же генотипами.
Р: АаВв (норм. слух) х АаВв (норм. слух)
F2: 9 А-В- (норм. слух): 3 А-вв (глух.): 3 ааВ- (глух.): 1аавв (глух.)
2.2. Эпистаз
Эпистаз – подавление действия одного гена действием другого гена, неаллельного первому.
Различают доминантный (доминантный аллель эпистатирует, или подавляет действие доминантного аллеля другого гена) и рецессивный (рецессивный аллель эпистатирует проявление доминантного аллеля другого гена). Анализ при эпистазе ведется так же на основе дигибридного скрещивания. Цифровые расщепления при эпистазе 12:3:1 (если каждый из двух генов имел самостоятельное проявление, при этом в первом поколении признак особей повторяет фенотип одного из родителей) и 13:3 (ген-ингибитор не имеет самостоятельного проявления, при этом фенотип родителей и первого поколения одинаков).
Подавляющий ген называется геном-супрессором, или ингибитором, а подавляемый – гипостатическим. Этот тип взаимодействия наиболее характерен для генов, участвующих в регуляции работы генов в онтогенезе и работе иммунных систем человека.
Примером рецессивного эпистаза у человека может служить «бомбейский феномен». В Индии описана семья, в которой родители имели вторую (А0) и первую (00) группу крови, а дети – четвертую (АВ) и первую (00). Чтобы ребенок в такой семье имел группу крови АВ, мать должна иметь группу крови В, но не 0. Позже было выяснено, что в системе групп крови АВ0 имеются рецессивные гены-модификаторы, которые в гомозиготном состоянии подавляют проявление антигенов на поверхности эритроцитов. Например, человек с третьей группой крови должен иметь на поверхности эритроцитов антиген группы В, но эпистатирующий ген-супрессор в рецессивном гомозиготном состоянии (hh) подавляет действие гена В, так что соответствующие антигены не образуются, и фенотипически проявляется группа крови 0. Описанный локус гена-супрессора не сцеплен с локусов АВ0. Гены-супрессоры наследуются независимо от генов, определяющих группы крови АВ0. Бомбейский феномен имеет частоту 1:13000 среди индусов, живущих в окрестностях Бомбея.
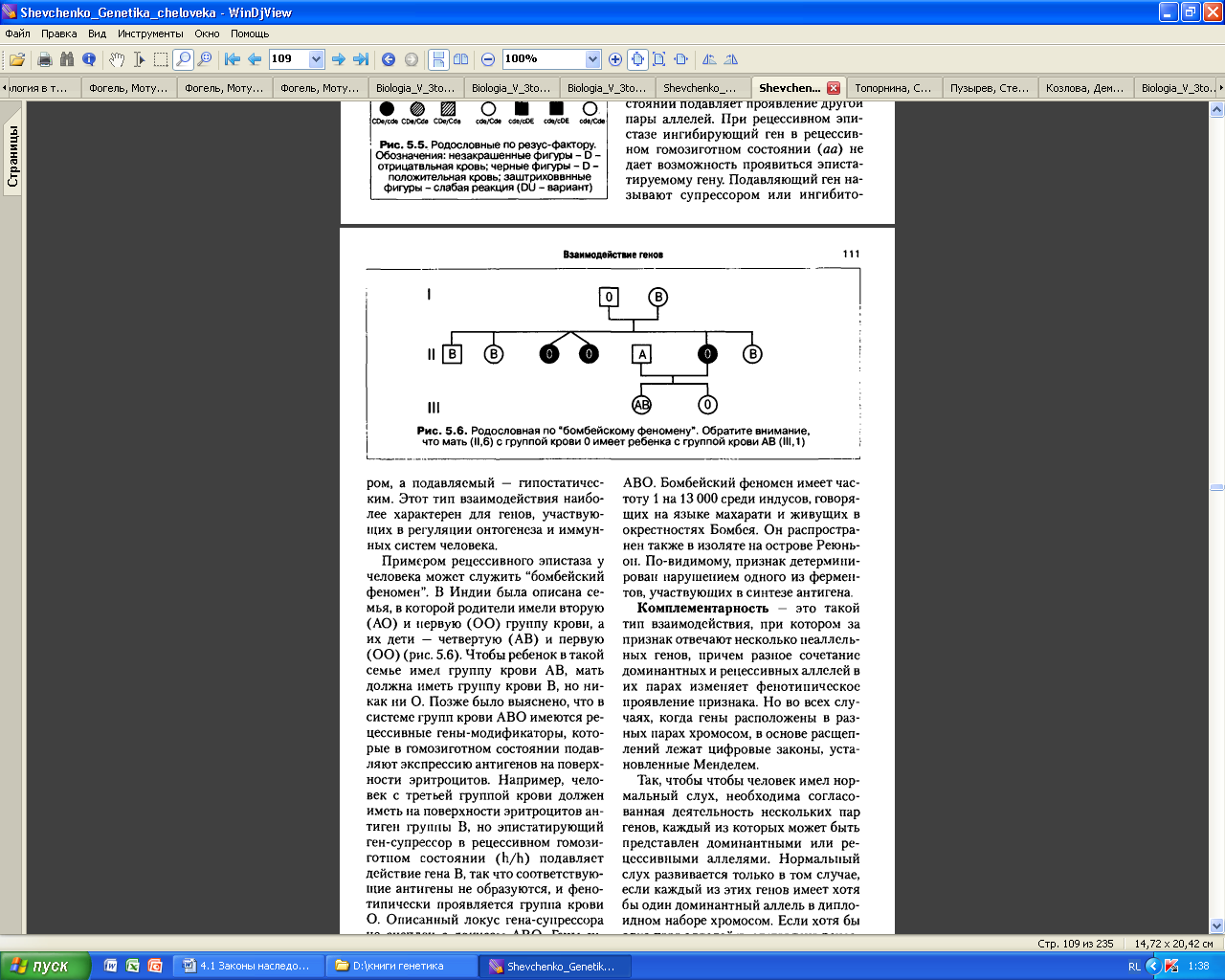
P hhIBIB (III) x HhIAIA
F1 HhIAIB , hhIAIB.
2.3 Полимерия
Полимерия – обусловленность определенного (обычно количественного) признака несколькими эквивалентными (полимерными) генами.
При таком взаимодействии индивидуальное проявление каждого усиливается в результате взаимодействия. Полимерия бывает двух видов:
А. некумулятивная – когда не важно количество доминантных генов в генотипе, а важно его присутствие.
Б. кумулятивная – когда число доминантных аллелей влияет на степень выраженности данного признака.
У человека по типу кумулятивной полимерии наследуется пигментация кожи: чем больше доминантных аллелей, тем больше меланина образуется и тем интенсивнее окраска кожи.
Например, необходимо определить вероятность появления детей с разными фенотипами в браке мулатов (особи дигетерозиготны.
Р АаА1а1 (мулатка) х АаА1а1 (мулат)
| ♀ ♂ | АА1 | Аа1 | аА1 | аа1 |
| АА1 | ААА1А1 (чернокожий) | ААА1а1 | АаА1А1 | АаА1а1 мулат |
| Аа1 | ААА1а1 | ААа1а1 мулат | АаА1а1 мулат | Ааа1а1 Светлокожий мулат |
| аА1 | АаА1А1 | АаА1а1 Мулат | ааА1А1 мулат | ааА1а1 светлокожий мулат |
| аа1 | АаА1а1 мулат | Ааа1а1 Светлокожий мулат | ааА1а1 светлокожий мулат | ааа1а1 белый |
Ответ: чернокожие (ААА1А1) -1; (ААА1а1) – 4; мулаты (ААа1а1) – 6; светлокожие мулаты (Ааа1а1) – 4; белые (ааа1а1) – 1.
Полимерные гены, как правило, обозначаются одинаковыми буквами, чтобы подчеркнуть однонаправленность их действия, т.к. при формировании признака не важно, какому гену принадлежат доминантные аллели, а важно их число.
Меланин – пигмент, определяющий все многообразие окрасок покровов. Все цвета волос, за исключением рыжих, составляют непрерывный ряд от темного до светлого, что соответствует уменьшению концентрации пигмента меланина, и наследуются полигенно.
По принципу полимерии наследуются также рост и масса тела.
Плейотропное действие генов
Явления взаимодействия генов указывает на целостность генотипа при индивидуальном развитии организма, а признание принципа взаимодействия допускает, что все гены так или иначе взаимосвязаны в своем действии. Исходя из этого следует, что и один ген может влиять на несколько признаков. Такое множественное действие гена называют плейотропией.
Плейотропное действие генов имеет различные механизмы. Первичная плейотропия обусловлена биохимическими механизмами действия мутантного белка или фермента. Мутантные аллели различных генов, контролирующих синтез коллагена и фибриллина, приводят к нарушению свойств соединительной ткани. Соединительная ткань является основой всех органов, поэтому понятно множественное действие этих мутаций на фенотип при таких наследственных заболеваниях соединительной ткани (синдром Марфана, синдром Элерса-Данло). Пример: У человека синдром Марфана обуславливается аутосомно-доминантным аллелем. Синдром Марфана связан с высоким ростом, значительным удлинением костей пальцев ног и пальцев рук (паучьи пальцы – арахнодактилия), гиперподвижностью суставов, подвывихом хрусталика и аневризмом аорты. Разные симптомы могут быть последствиями дефектов соединительной ткани.
При фенилкетонурии нарушается обмен фенилаланина, в результате чего не синтезируется тирозин. Вследствие этого уменьшается или прекращается образование меланина, что ведёт к гипопигментации кожи, волос и радужки. Патологические метаболиты (фенилпировиноградная кислота и др.) вызывают нарушение процессов развития и функционирования нервной системы (повышенная возбудимость, тремор, судорожные припадки, умственная отсталость). Казалось бы, это очень разнородные симптомы, но в основе всех этих множественных проявлений лежит первичный эффект недостаточности (или отсутствия) активности фенилаланингидроксилазы.
При вторичной плейотропии наблюдается множественность поражения организма, обусловленная осложнениями первичных патологических процессов, между которыми имеется взаимосвязь. Пример: При аутосомно-рецессивном заболевании муковисцидоз наблюдается ошибка в синтезе трансмембранного белка, обеспечивающего ионный транспорт в клетках эндокринных желез. Нарушение ионного транспорта натрия и хлора ведет к формированию густой слизи в поджелудочной железе, других эндокринных железах, бронхах, в результате чего возникают вторичные воспалительные процессы.
Типичный пример плейотропного заболевания – астма. Это заболевание в разной форме присуще почти всем людям. Практически каждый человек страдает аллергией на какой-либо раздражитель, если не с рождения, то в определенный период жизни. Астма, экзема, аллергия и анафилаксия – проявления одного синдрома, связанного с работой определенных клеток организма, активируемыми одними и теми же молекулами иммуноглобулина-Е. Один человек из десяти имеет клинические проявления аллергии – от легких приступов сенной лихорадки до анафилактического шока, который может развиться в считанные секунды после укуса пчелы или воздействия другого фактора и привести к смерти.
Таким образом, плейотропное действие генов обусловливает одну из характерных особенностей клинического проявления наследственных болезней — вовлечённость в патологический процесс многих систем и органов.
Дата добавления: 2021-01-20; просмотров: 310; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!