Наследование признаков, сцепленных с полом
Моногибридное скрещивание
Моногибридное скрещивание включает анализ наследования признаков, определяемых лишь одной парой аллельных генов. Мендель определил, что при скрещивании особей, отличающихся одной парой признаков, все потомство фенотипически однообразно. Здесь имеется в виду скрещивание гомозиготных особей, различных фенотипически. Например, при скрещивании гомозиготного желтого гороха (генотип АА) с гомозиготным зеленым (генотип аа) все потомство будет желтым, но гетерозиготным (генотип Аа). Ход скрещивания изображен в первой ступени схемы на рис. 5. Получившиеся гетерозиготные особи называются гибридами, а поскольку они гетерозиготны по одной паре генов, их называют моногибридами.
Скрещиваемые особи могут быть не обязательно гомозиготными. Для случаев, когда обе особи гетерозиготны, Менделем установлено: при скрещивании моногибридов во втором поколении происходит расщепление признаков на исходные родительские в отношении 3:1. 3/4 потомков оказывается с признаками, обусловленными доминантным геном, 1/4 — с признаками рецессивного гена.
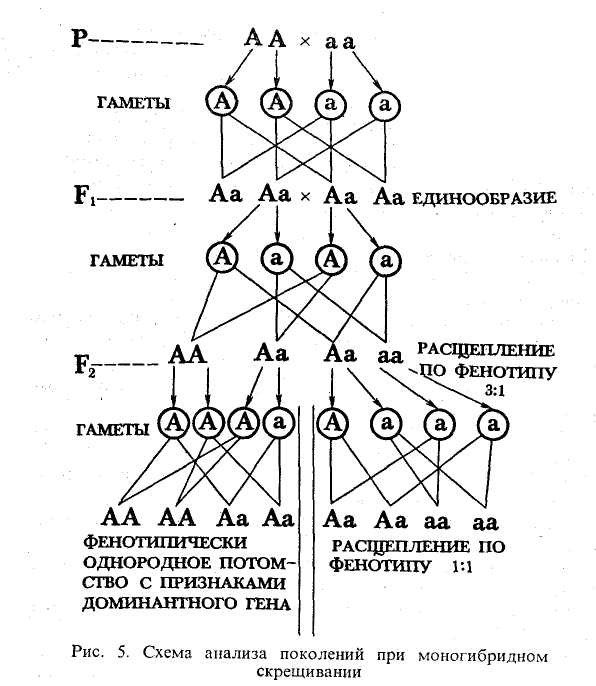 Как и почему происходит фенотипическое расщепление в отношении 3:1, можно понять из второй ступени схемы на рис. 5. Здесь важно обратить внимание на то, что у моногибридов образуется два типа гамет: гаметы с геном А и гаметы с геном а. И тех и других поровну. В процессе оплодотворения разные гаметы отцовского и материнского организмов имеют равновероятную возможность слиться друг с другом. Поэтому возможно формирование генотипов потомства: \/4АА, 2/4Аа и \/4аа. Фенотипически первые три будут с проявлением доминантного гена, один из четырех — с проявлением рецессивного гена. Правда, точное расщепление 3: 1 можно получить лишь при анализе бесконечно большого числа потомков. В случаях же малого числа их можно говорить только о вероятности появления особей с тем или иным признаком.
Как и почему происходит фенотипическое расщепление в отношении 3:1, можно понять из второй ступени схемы на рис. 5. Здесь важно обратить внимание на то, что у моногибридов образуется два типа гамет: гаметы с геном А и гаметы с геном а. И тех и других поровну. В процессе оплодотворения разные гаметы отцовского и материнского организмов имеют равновероятную возможность слиться друг с другом. Поэтому возможно формирование генотипов потомства: \/4АА, 2/4Аа и \/4аа. Фенотипически первые три будут с проявлением доминантного гена, один из четырех — с проявлением рецессивного гена. Правда, точное расщепление 3: 1 можно получить лишь при анализе бесконечно большого числа потомков. В случаях же малого числа их можно говорить только о вероятности появления особей с тем или иным признаком.
|
|
|
В генетике различают еще возвратное и анализирующее скрещивание. Возвратное — это скрещивание гибрида с гомозиготной особью (третья ступень схемы на рис. 5). Анализирующее -скрещивание гибрида с гомозиготной особью по рецессивным генам аллеля (правая часть третьей ступени схемы на рис, 5)
Ди- и полигибридное скрещивание
В природе не существует моногибридов и дигибридов. Каждый организм имеет несколько пар хромосом, в которых находится огромное число генов. Поэтому любой организм всегда гетерозиготен по многим аллелям. Однако для исследователя важно, сколько пар ал-лельных генов подвергается анализу. Если одна пара, его называют моногибридом, если две — дигибридом, три — тригибридом и т. д. При скрещивании моногибридов между собой в потомстве возможны только два фенотипа — исходные родительские. При скрещивании же ди- и полигибридов в потомстве появляются различные комбинации признаков, которых не было у родителей. Тем не менее при этом существуют тоже определенные закономерности. Мендель установил, что при скрещивании ди- и полигибридов во втором поколении происходит расщепление каждой пары признаков, независимо друг от друга, в отношении 3:1.
|
|
|
 Почему так происходит? Случаи независимого расщепления возможны тогда, когда гены каждой из анализируемых пар признаков находятся в разных парах хромосом. Вспомним, что во время мейоза гомологичные хромосомы расходятся в разные гаметы. Можно себе представить всю картину следующим образом. Если хромосома с геном А пойдет в одну гамету, то хромосома с геном а должна идти в другую гамету. Хромосома с геном В идет в одну гамету, с геном b— обязательно в другую. Но расхождение негомологичных хромосом не зависит одна от другой. Если в одну гамету ушла хромосома с геном А, то в эту же гамету может уйти хромосома с геном В. Если же с хромосомой А ушла хромосома с b, то хромосома с геном В должна уйти в гамету с хромосомой а. Иначе говоря, у дигибридов при расположении анализируемых аллелей в разных парах хромосом возможно четыре типа комбинаций генов в гаметах (рис. 6), и число этих комбинаций равновероятно — всех по 1/4. Вообще нужно иметь в виду, что у моногибридов возможно лишь 2 (21) типа гамет, у дигибридов —4 (22), у тригибридов —8 (23), т. е. при анализе я аллелей число возможных вариантов гамет равно 2". Решение задач на дигибридное скрещивание облегчается решеткой Пениста, составляемой соответственно числу возможных вариантов гамет. При скрещивании дигибридов она будет включать четыре типа мужских гамет, которые записываются по горизонтали, и четыре типа женских гамет, которые записываются по вертикали. Запись гамет следует производить строго в определенном порядке, как это показано на рис. 7. Тогда открываются интересные закономерности, облегчающие решение разнообразных задач. По одной диагонали собираются генотипы всех гомозиготных особей: в левом верхнем углу все. доминантные гены, ниже вправо — доминантные А и рецессивные b, дальше — рецессивные а и доминантные В, в правом нижнем углу — все рецессивные гены. Эта диагональ показывает все четыре класса фенотипов в случаях полного доминирования. По второй диагонали собираются все дигибриды. В остальных клеточках оказываются моногибриды. Характерное для скрещивания дигибридов расщепление фенотипов при условии полного доминирования в обоих аллелях 9:3:3:1 показано па рис. 7 штриховкой.
Почему так происходит? Случаи независимого расщепления возможны тогда, когда гены каждой из анализируемых пар признаков находятся в разных парах хромосом. Вспомним, что во время мейоза гомологичные хромосомы расходятся в разные гаметы. Можно себе представить всю картину следующим образом. Если хромосома с геном А пойдет в одну гамету, то хромосома с геном а должна идти в другую гамету. Хромосома с геном В идет в одну гамету, с геном b— обязательно в другую. Но расхождение негомологичных хромосом не зависит одна от другой. Если в одну гамету ушла хромосома с геном А, то в эту же гамету может уйти хромосома с геном В. Если же с хромосомой А ушла хромосома с b, то хромосома с геном В должна уйти в гамету с хромосомой а. Иначе говоря, у дигибридов при расположении анализируемых аллелей в разных парах хромосом возможно четыре типа комбинаций генов в гаметах (рис. 6), и число этих комбинаций равновероятно — всех по 1/4. Вообще нужно иметь в виду, что у моногибридов возможно лишь 2 (21) типа гамет, у дигибридов —4 (22), у тригибридов —8 (23), т. е. при анализе я аллелей число возможных вариантов гамет равно 2". Решение задач на дигибридное скрещивание облегчается решеткой Пениста, составляемой соответственно числу возможных вариантов гамет. При скрещивании дигибридов она будет включать четыре типа мужских гамет, которые записываются по горизонтали, и четыре типа женских гамет, которые записываются по вертикали. Запись гамет следует производить строго в определенном порядке, как это показано на рис. 7. Тогда открываются интересные закономерности, облегчающие решение разнообразных задач. По одной диагонали собираются генотипы всех гомозиготных особей: в левом верхнем углу все. доминантные гены, ниже вправо — доминантные А и рецессивные b, дальше — рецессивные а и доминантные В, в правом нижнем углу — все рецессивные гены. Эта диагональ показывает все четыре класса фенотипов в случаях полного доминирования. По второй диагонали собираются все дигибриды. В остальных клеточках оказываются моногибриды. Характерное для скрещивания дигибридов расщепление фенотипов при условии полного доминирования в обоих аллелях 9:3:3:1 показано па рис. 7 штриховкой.
|
|
|
|
|
|
По схеме рис. 7 можно проследить, например, ход скрещивания гороха, различающегося по двум парам признаков — цветности и форме горошины. Возьмем А — ген желтого цвета, а — зеленого цвета, В — ген гладкой формы горошины, b- ген морщинистой формы. При скрещивании гомозиготного желтого гладкого гороха с гомозиготным зеленым морщинистым все потомство оказывается по фенотипу желтым гладким (закон единообразия), по генотипу — дигетерозиготным, т. е. все растения — дигибриды. При скрещивании дигибридов между собой по фенотипу получается потомство четырех классов: желтые гладкие горошины, желтые морщинистые, зеленые гладкие и зеленые морщинистые. Соотношение их соответственно 9 :3 :3 :1. Два класса фенотипов — желтые морщинистые и зеленые гладкие — появляются в потомстве как новые комбинации. Но если мы будем рассматривать все потомство только по одной паре признаков, не обращая внимания на другую, то увидим независимое расщепление каждой пары признаков в отношении 3 : 1, Так, растений с желтыми горошинами оказалось всего 12 из 16, а зеленых — 4 из 16, т. е. 3: 1, хотя как желтые, так и зеленые горошины были и гладкими и морщинистыми. Точно так же с формой горошины: 12 гладких и четыре морщинистых, что соответствует 3:1.
По решетке Пеннета можно судить о характере расщепления потомства при разнообразных генотипах родителей. Например, при скрещивании дигибрида с гомозиготной рецессивной особью получается четыре класса фенотипов в отношении 1 :1 :1 : 1 (см. правый столбец на рис. 7). При скрещивании дигибрида с особью aabb получается два фенотипических класса в отношении 1 :1 (см. третий столбец на рис. 7). При скрещивании особи ААВЪ с ааВ b получится два фенотипических класса в отношении 3 :1 (см. четыре правые верхние клеточки). И так далее. 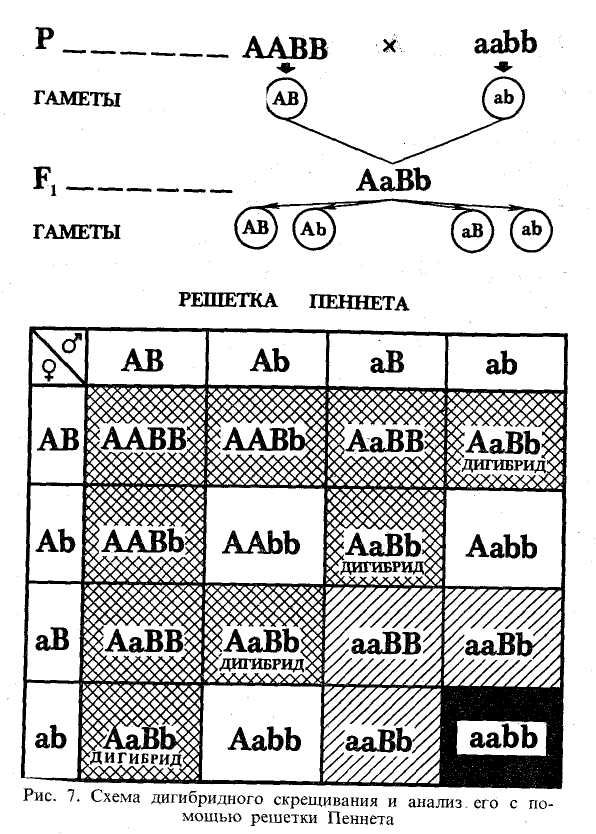
При скрещивании тригибридов происходит расщепление по фенотипу на восемь классов в отношении 27 :9 :9 :9 :3 :3 :3 :1 (рис. 8) В решетке Пеннета по диагонали с верхнего левого угла в правый нижний также идут гомозиготные особи, показывающие все возможные фенотипические классы. По второй диагонали располагаются все ■ тригибриды. Моногибриды образуют оригинальный орнамент: Сс копируют шахматное поле, ВЬ объединяются в восемь полей по четыре квадрата также в шахматном порядке, Аа объединяются в два поля по 16 квадратов в правом верхнем и левом нижнем углах. Решение задач на ди- и полигибридное скрещивание во многих случаях может производиться без вычерчивания решетки Пеннета, хотя решетка всегда дает более наглядную картину. Во всех случаях важно помнить ряд положений. Число возможных комбинаций гамет, т. е. число возможных генотипов в потомстве при прлигибридном скрещивании равно 4", что соответствует числу клеточек в решетке Пеннета. Возможное число вариантов (классов) фенотипов равно 2", а число вариантов (классов) генотипов - 3". При скрещивании полигибридов во втором поколении тоже происходит независимое расщепление каждой пары признаков в отношении 3 1.
Исходя из положения о независимом расщеплении каждой пары признаков, любое полигибридное скрещивание мы можем представить как л моногибридных скрещиваний. Тогда количественные соотношения классов фенотипов можно вычислять по формуле (3 + 1)". Вспомним, что при скрещивании моногибридов в потомстве вероятно три особи с проявлением доминантного гена А и одна особь с проявлением рецессивного гена а, иначе говоря, 3А + 1 a . To же будет, если подвергнуть анализу расщепление у дигибридов второй пары - ЗВ + \Ь. Следовательно, при скрещивании дигибридов потомство будет' (ЗА + la )- (3B + 1b) = 9 A В + 3АЬ + ЗаВ + lab , т. е. девять особей с проявлением одновременно признаков, обусловленных генами А и В, три особи с проявлением признаков А и b, три — с признаками а и В, одна особь с обоими признаками рецессивных генов а и b. Тригибрид-ное скрещивание можно представить как три моногибридных: (ЗА + la ).(3 B + 1 b )(ЗС + 1с). И так далее.
Можно подойти к решению задач на ди- и полигибридное скрещивание, зная лишь вероятности появления тех или иных генотипов или фенотипов при моногибридном скрещивании. Вспомним, что при моногибридном скрещивании вероятность появления потомства с генотипом А А равна 1/4, Аа -1/2, аа -1/4, а вероятность появления особей с признаками доминантного гена -3/4, с признаками рецессивного гена — 1/4. При скрещивании дигибридов вероятность появления особей с генотипом аа равна 1/4, с генотипом bb— также 1/4. Для того чтобы высчитать вероятность совпадения двух независимых друг от друга явлений, нужно перемножить вероятности каждого из них между собой. Следовательно, чтобы высчитать вероятность появления в потомстве дигибридов особей со всеми рецессивными признаками, надо 1/4 х 1/4 = 1/16. Чтобы высчитать то же при скрещивании тригибридов, надо 1/4 х 1/4 х 1/4 = 1/64. Чтобы высчитать, сколько получится ди-гетерозиготных особей от скрещивания дигибридов, следует перемножить вероятности получения моногибридов от моногибридного скрещивания; 1/2 х 1/2 = 1/4.
Таким образом можно рассчитать все возможные варианты генотипов и фенотипов от скрещивания полигибридов любой степени. Однако мы рекомендуем этими методами пользоваться лишь тогда, когда будут хорошо освоены все основные положения генетики или только в качестве показа их возможностей. Опасно то, что математические методы отвлекают учащихся и студентов младших курсов от главного: умения вывести правильно гаметы из известного генотипа и правильно скомбинировать генотипы потомства. Кроме того, подобного рода приемы сложны при анализе скрещиваний со сложными смешанными генотипами.
Неполное доминирование
При неполном доминировании доминантный ген неполностью подавляет действие аллельного гена. У гетерозигот функционирующими оказываются оба гена, поэтому в фенотипе признак выражается в виде промежуточной формы. Выраженность признака того или иного гена в ряде случаев может варьировать в определенных пределах. Так, по данным Дж. Ниля и У. Шэлла (1958), у гетерозигот по серповидноклеточной анемии содержание аномального гемоглобина колеблется в пределах от 25 до 45%. Закон единообразия первого поколения при неполном доминировании не теряет своего значения. Но во втором поколении потомство расщепляется фенотипически на три класса в отношении 1 :2 :1. При возвратном скрещивании потомство расщепляется в отношении 1 : 1. Разница состоит в том, что при скрещивании гибрида с гомозиготной доминантной формой получается половина особей с доминирующим признаком, половина — с промежуточным, а при скрещивании с гомозиготной рецессивной формой (анализирующее скрещивание) — половина особей с рецессивным признаком, половина — с промежуточным.
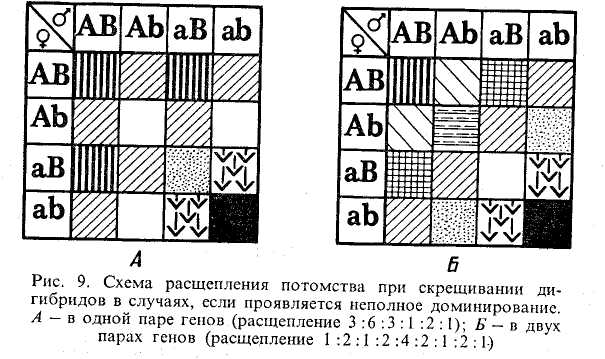
При анализе двух пар аллельных генов, в случаях, когда в одной паре проявляется неполное доминирование, расщепление гибридов во втором поколении произойдет не в отношении 9:3:3:1, а 3.6:3:1:2:1. Если обе пары генов будут проявлять неполное доминирование, то при скрещивании дигибридов произойдет расщепление второго поколения в отношении 1:2:1:2:4:2:1:2:1 (рис. 9).

Наследование признаков, сцепленных с полом
Сцепленными с полом называются признаки, гены которых расположены не в аутосомах, а в гетеросомах. Хромосомы, определяющие пол, отличаются одна от другой. Та, которая покрупнее, называется X-хромосомой, которая поменьше — У-хромосомой. В Х- и У-хромосомах есть какие-то участки, гомологичные друг другу и содержащие аллельные гены. Признаки таких генов должны менделировать, т е. подчиняться закону расщепления в отношении 3.1. Но как в Х-, так и в Y-хромосоме имеются негомологичные участки. В таких участках X-хромосомы содержатся гены, аллельных которым в У-хромосоме нет (например, ген классической гемофилии), и наоборот, в негомологичном участке Y-хромосомы содержатся гены, у которых нет аллелей в X-хромосоме (например, ген, определяющий развитие волосков на краю ушной раковины).
Вспомним, как определяется пол. Как правило, гетерохромосом в организме две. У млекопитающих, в том числе и у человека, клетки
женского организма содержат две одинаковые гетеросомы — X и X , а клетки мужского организма две разные — Х- и У-хромосомы. В процессе мейоза гетеросомы расходятся в разные гаметы. Поэтому все женские гаметы будут нести по одной Х-хромосоме. Мужские же гаметы будут двух типов: половина с X-хромосомой, половина с Y-хромосомой. При слиянии гамет во время оплодотворения определяется пол будущего организма: если яйцеклетка (в ней всегда, есть Х-хромосома) сольется со сперматозоидом, несущим Х-хромосому, будет развиваться женский организм, если же со сперматозоидом, несущим Y-хромосому, разовьется мужской организм. Поэтому говорят, что у млекопитающих женский пол является гомогаметным, мужской — гетерогаметным. Схема решения задач на наследование признаков, сцепленных с полом, иная, чем на аутосомное моногибридное скрещивание. В случае, если ген сцеплен с Y-хромосомой, он может передаваться из поколения в поколение только мужчинам. Если ген сцеплен с X-хромосо-мой, он может передаваться от отца только дочерям, а от матери в равной степени распределяется между дочерями и сыновьями. Если ген сцеплен с Х-хромосомой и является рецессивным, то у женщин он может проявляться только в гомозиготном состоянии. У мужчин второй Х-хромосомы нет, поэтому такой ген проявляется всегда. При анализе признаков, сцепленных с полом, необходимо изменить схему, изображенную на рис. 5, и ввести в запись не только условные обозначения генов, но и гетеросомы. Схема наследования дальтонизма (цветовая слепота, обусловленная рецессивным, сцепленным с Х-хромосомой, геном) изображена на рис. 10.
Следует иметь в виду, что, например, у птиц, некоторых насекомых гетерогаметный пол — женский, гомогаметный — мужской.
Множественные аллели
Иногда в популяции оказывается не два (одна пара) аллельных гена, а три, четыре и больше: A , a 1 a 2, и т. д. Возникают они в результате разных мутаций одного локуса. У каждой особи аллельных генов может быть не более двух, но в популяции их число практически неограниченно. Чем больше аллельных генов, тем больше возможностей комбинаций их попарно. Так, у морских свинок окраска шерсти определяется пятью аллелями одного локуса, которые в различных сочетаниях дают одиннадцать вариантов окраски. Гены множественных аллелей взаимодействуют между собой различным образом. Нередко они образуют последовательные ряды доминирования: А доминирует над a1,a2 и a3; a1доминируют над а2 и a3, а2 доминируют только над a3. Но бывают и более сложные комбинации. У человека по типу множественных аллелей наследуются группы крови по системе АВО.
Дата добавления: 2019-09-08; просмотров: 162; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!