Общая характеристика пирамидных нейронов.
Как уже указывалось выше, пирамидные нейроны делятся на три группы: самые малые — вставочные, средней величины — а ссо циационны е и крупные — проекционные.
Аксоны малых и некоторых средних пирамид не выходят за пределы коры. Они связывают звездчатые и другие нейроны с более или менее близко лежащими пирамидными нейронами разных слоев и потому их называют вставочными. Их можно найти во всех слоях, кроме первого, особенно много их во II и IV слоях коры.
Аксоны значительной части средних пирамид выходят в белое подкорковое вещество и идут к другим участкам того же полушария или к другому полушарию через мозолистое тело. Эти нейроны принято называть ассоциационными, они лежат также во всех слоях коры, кроме первого, но особенно много их в средних слоях коры (рис. 3, нейроны 4, 7, 10, 17).
Большие пирамиды V и VI слоев, в основном, принадлежат проекционной эфферентной системе. Их аксоны уходят через белое подкорковое вещество к нижележащим отделам мозга, образуя пирамидные и экстрапирамидные пути (рис. 3, нейрон 8).
Аксоны ассоциационных и проекционных пирамидных клеток имеют множество коллатералей, которые в основном идут в горизонтальном направлении и оканчиваются обычно на дендритах соседних пирамидных нейронов или даже на дендритах собственного нейрона (Поляков, 9). Многие коллатерали поднимаются в верхние слои — это так называемые возвратные коллатерали (рис. 3, нейроны 7, 10). Многие из них доходят до / слоя, оканчиваясь главным образом в / и II слоях на верхушечных дендритах пирамидных нейронов. Конечные же разветвления ассоциационных волокон оканчиваются главным образом на телах пирамидных и звездчатых клеток в разных участках коры, обычно во // и /// слоях (рис. 27, а, Ь, с). С помощью этих окончаний осуществляется объединение нейронов коры в функциональ-
36
ные нервные круги при восприятии внешнего мира. Конечные же разветвления проекционных пирамидных волокон оканчиваются во всех других отделах мозга и служат в основном для интегрирования деятельности этих отделов.
Структурная характеристика дендритов пирамидных нейронов.
Как указывалось выше, апикальные и базальные дендриты густо покрыты шипика ми. Впервые Суханов (22), а затем многие исследователи подробно изучали эти образования. В последнее время Поляков и Саркисов (1), затем Чанг (10) нашли, что шаровидная головка шипика около 1 мк соединяется с дендритом тоненькой ножкой длиной в 0,5 мк и потому легко отрывается от него. Шипики очень разнообразны по форме и размерам. Количество шипиков на дендритах одного пирамидного нейрона может доходить до 6000. Их особенно много на апикальных дендритах пирамидных нейронов в верхних слоях коры, где на 100 мк их длины приходится 80 шипиков (Пурпура, 23). Поляков, Чанг и другие отмечают, что тело клетки и начальные отделы дендритов лишены шипиков (рис. 28).
По данным Саркисова и Полякова (1,12), у человека в онтогенезе шипики начинают появляться относительно поздно, лишь к моменту рождения. Среди различных нервных клеток раньше всего шипики обнаруживаются у наиболее рано созревающих, крупных пирамидных клеток V слоя и только потом они появляются у других клеток по мере их созревания. Шипики очень чувствительны к различным вредоносным и токсическим агентам. Так, при

Рис 28. Пирамидный нейро н по Полякову
На всех дендритах имеются боковые придатки — шипики, но они начинаются на некотором расстоянии от клетки Само тело клетки, а также более или менее толстые части дендритов лишены шипиков Ясно в и (но, что на апикальных дендритах шипики расположены гуще, чем на базальных. На дендритах данного пирамидного нейрона 3497 шипиков, а — аксон, с — коллатераль
37
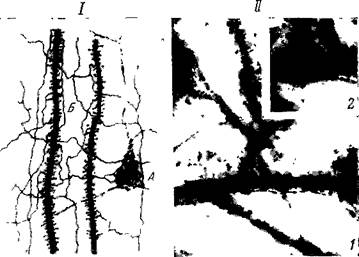
Рис. 29. С ин апт и ческие связи
I — на дендритах и теле клетки (по Чангу): Л — пирамидная клетка, вся покрытая синапсами, образующими вокруг клетки и толстой части дендрита сптетение вроде корзинки, Б—два дендрита с шипиками, часть которых соприкасается с боковыми и конечными синапсами аксонных разветвлений. II — на дендритах Микрофотография (по Полякову): 1 — дендриты с боковыми придатками, ясно видно, что одно нервное волоконце прикасается к ряду боковых придатков; 2 — то же при большом увеличении
перевязке сонных артерий уже через полчаса наступают значительные изменения нормальной структуры шипиков, а при более длительных нарушениях питания нервной ткани они претерпевают полный распад.
По мнению Саркисова, Полякова и Чанга, шипики являются рецепторным аппаратом. И в самом деле, физиологические влияния на дендриты осуществляются главным образом через шипики. В / слое, где преобладают разветвления верхушечных дендритов, имеется большое количество горизонтальных нервных волокон от вставочных нейронов //, /// и IV слоев с восходящими аксонами, конечных разветвлений ассоциацыонных нейронов и их возвратных коллатералей, а также волокон от горизонтальных нейронов первого слоя. Все эти волокна устанавливают синаптические связи с означенными дендритами главным образом через шипиковый аппарат (рис. 29).
С помощью электронномикроскопического исследования обнаружили, что синапсы на шипиках ствола дендрита— боковые синапсы —существенно отличаются от синапсов на конечных разветвлениях апикальных дендритов в первом слое (рис. 30). Эти терминальные синапсы в 3—4 раза крупнее боковых и содержат по 3 митохондрии, в то время как боковые совсем не содержат митохондрий. Кроме того, сам шипиковый аппарат в конечных раз ветвлениях апикальных дендритов значительно крупнее, чем на стволе дендрита и содержит по 4 митохондрии. Все остальные шипики содержат всего по одной митохондрии. Синаптические аппараты на теле клетки и на начальных участках дендрита, где нет шипиков, значительно меньше терминальных синапсов и содержат только по одной митохондрии (Микеладзе, 24).
Дата добавления: 2019-07-17; просмотров: 387; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!