Морфофункціональний стан головного мозку інтактних тварин.
Отримані дані показали, що всі досліджувальні показники інтактної групи тварин знаходяться в межах фізіологічної норми. Коливання маси тіла інтактних кролів було незначним, в межах нормального фізіологічного росту - 4,14±0,37 до 4,87±0,41 кг. Тварини були активними, мали густий та блискучий шерстяний покрив, добрий апетит.
Біохімічним дослідженням ліпідного ліпідного спектру сироватки крові тварин встановлено, що вміст ЗХ знаходяться в межах 0,85±0,06 - 0,88±0,09 мМ/л; ЛПНЩ - 1,35±0,12 - 1,48±0,16 г/л; ЛПВЩ – 6,08±0,90 - 6,33±0,83 мМ/л; ТГ – 0,51±0,20 - 1,00±0,25 мкМ/л; ФЛ – 0,97±0,15 - 1,08±0,21 г/л; ІА – 0,82±0,17 - 0,91±0,18.
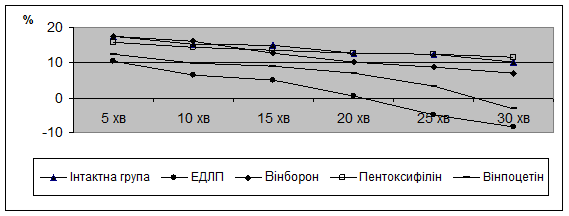 Об’ємна швидкость мозкового кровотоку (ОШМК) у внутрішніх сонних артеріях, основною функцією яких є транспорт крові до мозку, в тварин інтактної групи знижується повільно з 17,56±0,29 до 12,62±0,22 % протягом 30 хв. дослідження (рис. 1).
Об’ємна швидкость мозкового кровотоку (ОШМК) у внутрішніх сонних артеріях, основною функцією яких є транспорт крові до мозку, в тварин інтактної групи знижується повільно з 17,56±0,29 до 12,62±0,22 % протягом 30 хв. дослідження (рис. 1).
Рис.1. Динаміка об’ємної швидкості мозкового кровотоку при ЕДЛП та її фармакокорекції.
Макроморфометричними вимірами параметрів головного мозку інтактних тварин було встановлено, що маса головного мозку кролів становить 13,00±0,71 г, об’єм головного мозку - 12,50±0,78 г/мг. Параметри довжини головного мозку складають - 44,60±3,44мм, довжина півкуль - 30,00±0,71мм. Висота мозку - 17,00±1,22мм, ширина мозку - 33,00±1,58 мм.
При мікроморфометричному дослідженні артерій малого калібру м’якої мозкової оболонки інтактної групи тварин було встановлено, що зовнішній їх діаметр становить 42,89±1,66 мкм, внутрішній – 23,99±0,51 мкм, товщина стінки – 9,44±0,61 мкм. Площа поперечного перерізу артерій малого калібру м’якої мозкової оболонки становить 472,67±36,86 мкм2, площа просвіту – 155,97±6,62 мкм2, площа стінки – 316,69±32,35 мкм2 (табл. 1).
|
|
|
Таблиця 1
Морфометричні параметри артерій малого калібру м’якої мозкової оболонки при ЕДЛП та її фармакокорекції (М±m).
| Показник | Групи тварин | ||||
| Інтактна | ЕДЛП | Вінборон | Пентоксифілін | Вінпоцетін | |
| Площа перерізу (мкм2) | 472,67± 36,85 | 786,48± 21,72 * | 536,03± 15,69*/** | 650,97 ±32,08*/** | 607,03± 40,76*/** |
| Зовнішній діаметр (мкм) | 42,89± 1,66 | 47,05± 0,53 * | 43,86± 0,57*/** | 45,58± 0,38*/** | 43,38± 0,57*/** |
| Внутрішній діаметр (мкм) | 23,99± 0,51 | 16,04± 0,95 * | 22,71± 1,24*/** | 18,07± 2,14* | 19,93± 0,85*/** |
| Площа просвіту (мкм2) | 155,97± 6,62 | 107,76± 0,84 * | 131,15± 1,59*/** | 123,82± 2,28*/** | 127,40± 1,26*/** |
| Площа стінки (мкм2) | 316,69± 32,35 | 677,07± 22,10 * | 404,88± 16,54*/** | 529,67± 30,25*/** | 479,62± 39,54*/** |
| Товщина стінки (мкм) | 9,44±0,61 | 15,50±0,54 * | 10,57± 0,83*/** | 13,24± 1,05*/** | 11,72± 0,53*/** |
| Індекс Вогенворта (%) | 206,57± 20,75 | 628,93± 17,81 * | 317,53± 15,59*/** | 427,67± 21,07*/** | 371,62± 26,65*/** |
Примітка. * - різниця достовірна порівняно з інтактною групою (р≤0,01);
** - різниця достовірна порівняно з групою ЕДЛП (р≤0,01).
|
|
|
При гістохімічному дослідженні в стінці артерій м’якої мозкової оболонки та капілярах кори головного мозку ліпіди не виявляються. Наявність ліпідів спостерігається у вигляді дрібних крапель та напилення в нервових клітинах кори головного мозку.
Результати морфологічного світлооптичного дослідження кори головного мозку інтактних тварин показали, що м’яка мозкова оболонка, яка вкриває кору головного мозку, містить дрібні артерії, від яких в кору розгалужуються багаточисельні капіляри. В корі великого мозку кролів під м’якою мозковою оболонкою спостерігаються такі структурні компоненти: тіла клітин з відростками, їх ядра, ядерця; капіляри. Розрізняються тіла нейронів і тіла нейроглії. Нейрони утворюють в корі головного мозку шість шарів: І – молекулярний, ІІ – зовнішній зернистий, ІІІ – зовнішній пірамідний, ІV – внутрішній зернистий, V – внутрішній пірамідний (гангліонарний), VІ – шар поліморфних клітин.
За характерними особливостями цитоплазми і ядер, а саме вмістом і розподілом в ній тигроїду (гранул хроматофільної субстанції) пірамідні нейрони кори головного мозку поділяються на різні типи: нормохромні, гірохромні, різко гіпохромні, гіперхромні, різко гіперхромні та клітини „тіні”. Результатами визначення вмісту різних типів нейронів, що відображають морфофункціональний стан цих клітин встановлено, що у інтактної групи тварин в ІІІ і V шарах переважають нормохромні (86,61±1,37 %) і невеликий відсоток гіпо- та гіперхромних нейронів. Нормохромні нейрони мають середню інтенсивність забарвлення цитоплазми і рівномірно розподілені в ній гранули хроматофільної субстанції. Ядра таких нейронів, в порівнянні з цитоплазмою, більш світлі, ядерця центрально розміщені або дещо зміщені до ядерної оболонки. Гіпохромні нейрони – частіше з периферичним хроматолізом. Гіперхромні нервові клітини мають темну базофільну цитоплазму та каріоплазму, тому ядра і ядерця погано виявляються. В тварин інтактної групи пірамідні нейрони мають добре виражене клітинне тіло, об’єм якого складає в середньому 3270,11±165,36 мкм3. Цитоплазма заповнена дрібнозернистим тигроїдом. Об’єм цитоплазми - 1786,99 ± 0,13 мкм3. Округлі або овальні ядра мають чітку оболонку та містять одне, рідше два темних ядерця. Об’єм ядер становить - 1483,12 ± 3,51 мкм3, ядерець - 26,42±0,03 мкм3.
|
|
|
Клітини нейроглії в корі великих півкуль представлені двома різновидами: протоплазматичними астроцитами та клітинами мікроглії. Нейрогліальні клітини характеризуються малими розмірами з овальними чи продовгуватими ядрами.
|
|
|
При електронномікроскопічному дослідженні кори великого мозку інтактних тварин виявлено, що серед пірамідальних нейроцитів ІІІ і V шарів кори головного мозку переважають клітини з помірною електронною щільністю, що на світлооптичному рівні відповідають нормохромним нейроцитам. Такі клітини мають ядро з добре вираженим ядерцем, багато рибосомальних гранул, світлу каріоплазму, високу насиченість органел, високий вміст гранулярної ендоплазматичної сітки, велику кількість полірибосом, мітохондрії з добре вираженим матриксом В корі великого мозку інтактних тварин виявляються гемокапіляри соматичного типу з вузьким просвітом, неперервною цитоплазмою ендотеліоцитів. Поверхня ендотеліоцитів гладенька, її плазмолема утворює мікроворсинки та має неглибокі інвагінації. Базальна мембрана чітко контурована, має рівномірну товщину.
Морфофункціональні зміни головного мозку при експериментальній дисліпопротеїдемії
Результати біохімічного дослідження сироватки крові показали, що внаслідок тривалого холестеринового навантаження у кролів порушується ліпідний обмін. Це проявляється зростанням рівня ЗХ майже в 5 разів, ЛПНЩ – в 5,3 раза, ІА – в 1,76 раза, ФЛ – в 2,66 раза, та зниженням концентрації ЛПВЩ – в 13,6 раза порівняно з показниками інтактної групи.
В сонних артеріях, в тварин з ЕДЛП спостерігається прогресивне зниження об’ємної швидкості мозкового кровотоку (ОШМК) з 10,34±0,50 % через 5 хв до мінус 8,28±0,39 % через 30 хв., проти більш стабільного зменшення ОШМК з 17,56±0,29 % до 10,10 ± 0,22 % у кролів інтактної групи за цей самий проміжок часу (див. рис.1).
Макроморфометричним дослідженням параметрів головного мозкувстановлено, що тривале навантаження тварин холестерином призводить до явищ дистрофії та атрофії головного мозку. Так, у тварин з ЕДЛП маса головного мозку зменшується на 13,08 %, об’єм - на 12,88 % та довжина головного мозку зменшується - на 14,35 %, довжина півкуль зменшується на - 17,33 %, висота - на 11,76 %, ширина - на 3,63 % в порівняні з відповідними даними інтактної групи тварин.
При мікроморфометричому дослідження встановлено, що у кролів з ЕДЛП в корі головного мозку виявляються структурно-функціональні зміни. В першу чергу порушення виникають у артеріях малого калібру м’якої мозкової оболонки, що має суттєве значення, оскільки вони відіграють одну з основних ролей у кровопостачанні органів (С.В.Шорманов, 1982). В результаті дослідження встановлено збільшення площі поперечного перерізу на 66,39 %, зовнішнього діаметру – на 9,70 %, товщини стінки судин – на 64,19 %, площі стінки – в 2,14 рази, та індексу Вогенворта – в 3,04 рази, а також зменшення площі просвіту на 30,90 % та внутрішнього діаметру артерій – на 33,14 % в порівнянні з групою інтактних тварин ( див. табл.1.).
Звуження просвіту та суттєве зростання індексу Вогенворта, свідчить про істотне зменшення пропускної здатності артерій малого калібру м’якої мозкової оболонки в умовах змодельованої патології (С.В. Шорманов, 1982). Судини мікроциркуляторного русла, які відносяться до третього структурно-функціонального - метаболічного - рівня артеріальної системи мозку, також перетерплюють значні зміни. В мікроциркуляторному руслі, в умовах даної патології, зменшується кількість функціонуючих капілярів. Діаметр їх різний по довжині, капіляри звужуються та супроводжуються багаточисельними гліальними елементами.
При гістохімічному дослідженні кори головного мозку кролів з ЕДЛП спостерігається поява ліпідів в стінці артерій м’якої мозкової оболонки та капілярів кори головного мозку у вигляді дрібних крапель та напилення.
Порушення постійного притоку крові до певних ділянок мозку викликає дистрофічні зміни і загибель клітин мозку з наступним порушенням функцій. Мікроморфометричним дослідженням встановлено, що найбільш чутливими до ішемії є пірамідні нейрони кори головного мозку, що співпадає з даними інших авторів (В.В. Серева, М.А. Пальцев, 1998). Серед пірамідних нейронів ІІІ і V шарів кори головного мозку переважають клітини з вираженими проявами ішемії. Ядро і цитоплазма таких клітин вже не має чіткої структурованості. Встановлено зменшення розмірів і зміна будови пірамідних нейронів у тварин з ЕДЛП. В порівнянні з інтактною групою об’єм тіл цих клітин у цих тварин зменшується на 45,27 %, об’єм цитоплазми - на 39,92 %, об’єм ядра - на 51,69 %, об’єм ядерця - на 55,46 %.
Відмічено також зменшення кількості нормохромних пірамідних нейроцитів на 34,18 %, за рахунок збільшення кількості гіперхромних, гіпохромних, різко гіпо - і гіперхромних, а також „клітин-тіней” (табл.2).
Таблиця 2.
Кількість різних типів пірамідних нейронів в ІІІ і V шарах (%) сенсомоторної кори головного мозку при ЕДЛП та її фармакокорекції (М±m).
| Група тварин | Типи клітин (%) | |||||
| Нормо-хромні | Гіпохром-ні | Гіпер-хромні | Різко гіпохро-ні | Різко гіпер- хромні | Клітини-тіні | |
| Інтакні | 86,61±1,37 | 6,56±0,98 | 4,41±0,46 | 0,64±0,41 | 0,32±0,19 | 0,97±0,36 |
| ЕДЛП | 52,43±2,42* | 5,02±1,08 | 8,97±1,27* | 6,57±1,02 * | 21,97±1,28 * | 5,28±0,70 * |
| Вінбо-рон | 81,03±1,92*/** | 7,82±0,70 ** | 6,03±1,96 | 1,36±0,35 ** | 0,85±0,49 ** | 1,05±0,35 ** |
| Пенток-сифілін | 75,96±2,31*/** | 8,80±0,61*/** | 7,88±1,75* | 3,05±0,24 */** | 2,43±0,25 */** | 1,76±0,24 */** |
| Вінпо-цетін | 78,45±1,05*/** | 8,69±0,53*/** | 6,32±0,51* | 3,27±0,62 */** | 1,60±0,45 */** | 1,34±0,22 ** |
Примітка. * - різниця достовірна порівняно з інтактною групою (р≤0,01);
** - різниця достовірна порівняно з групою ЕДЛП (р≤0,01).
Різко гіпохромні нейрони бідні хроматофільною субстанцією, слабо зафарбовані, в основному з набряклими ядрами, ядерця майже не проглядаються, різко гіперхромні (темні) нейрони інтенсивно зафарбовані, зменшене ядро заповнено інтенсивно зафарбованим ядерним вмістом, в якому погано розрізняються темні ядерця. Верхівковий дендрит у цих клітин прослідковується на великій відстані.
Зростання числа гіперхромних і різко гіперхромних нейроцитів свідчить про розвиток дистрофічних явищ, які є морфологічним відображенням порушень метаболізму в клітинах мозку (Н.В Верещагин, 1997). Чим чіткіше виражена стадія ішемії клітини, тим більше звужується і зменшується її тіло і ядро та збільшується дифузність тигроїда. Ішемічні пірамідні клітини розміщуються групами. У частини таких клітин спостерігається каріолізис, деякі з них перетворюються на „клітини-тіні”. На місці загиблих нейроцитів зустрічаються вогнища макрофагальної інфільтрації. Регресивно змінюються також клітини макро - і мікроглії (пікноз і інтенсивна базофілія ядер). Збільшення числа клітин-тіней і заміщення загиблих нейронів клітинами мікроглії у тварин з ЕДЛП вказує на виснаження резервних можливостей нейронів сенсомоторної кори. Наявність у тварин з ЕДЛП достовірно більшої кількості, в порівнянні з інтактними тваринами, ішемічно змінених нейронів є однією із ознак вираженої гіпоксії мозку (Н.Н. Боголепов та ін, 2003).
Електронномікроскопічні дослідження 3 і 5 шарів кори великого мозку в умовах ЕДЛП показали, що виникає деструкція та дистрофія органел нейроцитів. Спостерігаються „світлі” та інтенсивно „світлі” нервові клітини (аналог гіпохромним і різко гіпохромних), а також „темні” та інтенсивно „темні” (аналог гіперхромним і різко гіперхромним) нейроцити в яких наявні значні патологічні зміни ядра і цитоплазматичних структур. В інтенсивно “світлиих” нейроцитах спостерігаються поодинокі пошкоджені органели, порушення каріолеми, світла нейроплазма. В інтенсивно “темних” нейроцитах: фрагментація ядра, фрагментація канальців ГЕС, руйнування мітохондрій. Кровоносні капіляри характеризуються розширеним просвітом, в просвітленій, неширокій цитоплазмі ендотеліоцитів мало органел, піноцитозних пухирців. Внутрішня поверхня ендотеліоцитів відносно рівна, плазмолема не має мікроворсинок, базальна мембрана тонка.
Дата добавления: 2019-07-15; просмотров: 81; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!