Возбуждения ЭТИХ КЛеТОК, Vm . nx П 2 страница
Триггерные ритмы, связанные с ин токсикацией дигиталисом (волокна Пуркинье) и зависящие от воздейст вия адреналина на предсердные клетки коронарного синуса, имеют как
сходство, так и различия [Johnson N. el a)., 1986]. В лервом случае, по-видимому, сразу же устанавливается постоянная частота эктопического ритма; во-втором — отмечается постепенное укорочение длины циклов, т. е. «разогрев» ритма с последующей его стабилизацией. Дигиталисно-токсиче-ские триггерные ритмы могут прерываться остро. Триггерный ритм, исходящий из коронарного синуса, замедляется постепенно («охлаждение») до момента исчезновения. Правда, эти факты еще нуждаются в дальнейшем клиническом подтверждении.
Имеет смысл рассмотреть некоторые вопросы дифференциальной диагностики триггер-ных, анормальных автоматических и реципрокных (re-entry) ритмов в экспериментальной работе и в процессе клинических электрофизиологических исследований. Пока эта проблема не может считаться решенной, но мы усматриваем определенную пользу в систематизации имеющихся в литературе электрофизиологических данных, что должно приблизить нас к распознаванию механизмов аритмий у больных. Что касается задержанных постдеполяризаций и триггер-ных ритмов, то мы использовали в основном результаты, получаемые при дигиталисной интоксикации [Johnson N. et al., 1986; Rosen M., 1986, 1988]:
а) задержанные постдеполяризации (триггерные ритмы) могут быть вызваны одним-двумя преждевременными экстрастимулами, т. е. тем же приемом, что и re-enlry. Однако воспроизводимость тригтерных ритмов при достижении критического интервала экстрастимула обычно ниже 30% (в волокнах Пуркинье), тогда как воспроизводимость re-entry высока [Moak J., Roscn M., 1984]. Кроме того, длина возвратного цикла вслед за преждевременным экстрастимулом, вызвавшим задержанные постдеполяризации, укорачивается по мере уменьшения интервала сцепления экстрастимула (прямая зависимость). При re-entry зависимость об-
|
|
|
ратная: интервал между экстрастимулом и первым тахикардическим комплексом удлиняется при укорочении интервала сцепления преждевременного экстрастимула. Это — важный признак re-entry [Rosen M., 1986, 1988]. С помощью преждевременных экстрастимулов не удается генерировать анормальную автоматическую активность;
б) задержанные постдеполяризации (триггерные ритмы) значительно легче прерываются под воздействием одиночных преждевременных экстрастимулов, чем анормальные автоматические ритмы. Подавление триггерных ритмов, формирующихся в волокнах Пуркинье периинфаркт-ной зоны, достигается более легко с помощью ранних экстрастимулов, чем поздних [El-Sherif N. et al., 1983; Dangman K., Hoffman В., 1985]. Триг-герная активность в мышечных волокнах митрального клапана в равной степени устраняется как ранними, так и поздними экстрастимулами [Wit A., Cranefield P., 1976];
|
|
|
в) электрическая стимуляция длительностью 15 с и длиной цикла около 300 мс быстро прерывает триггерные ритмы, в основе которых лежат задержанные постдеполяризации, например дигиталисно-токсические, но она слабее влияет на анормальные автоматические ритмы, как, впрочем, и на триггерные ритмы, зависящие от ранних постдеполяризаций [Rosen M., 1986].
Еще одно замечание имеет отношение к терминологии. В литературе нередко можно встретить термин «триггерный автоматизм», что, по сути дела, неверно, так как триггерные (наведенные) ритмы не связаны со спонтанной диастолической деполяризацией — автоматизмом.
НАРУШЕНИЯ ПРОВОДИМОСТИ
Движение импульса в сердечном волокне зависит от нескольких взаимодействующих факторов: силы электрического стимула — ПД в возбуж-
денном участке волокна, электрического ответа соседнего еще не возбудившегося участка волокна, межклеточного электротонического взаимодействия, пассивных свойств клеточных мембран, анатомических особенностей строения волокон (их величины, типа, геометрии, направленности). Изменения каждого из этих факторов могут приводить к нарушениям проводимости [Gettes L. et al., 1985].
|
|
|
Нередко причиной медленного проведения или блокады бывает снижение потенциала покоя (максимального диастолического потенциала) в клетках, которым в нормальных условиях свойствен быстрый электрический ответ (клетки Пуркинье, сократительные клетки предсердий и желудочков). Скорость проведения импульса в этих клетках непосредственно связана с крутизной и амплитудой фазы О ПД, т. е. с такими характеристиками, которые определяются процентом открытых быстрых Na каналов мембраны в момент возбуждения и натриевым электрохимическим градиентом (соотношение вне- и внутриклеточной концентраций ионов Na+). В свою очередь, существует тесная зависимость между процентом Na каналов, способных к открытию, и величиной максимального диастоличе-ческого потенциала мембраны. Если под влиянием патологических воздействий он понижается, уменьшается и ПД, соответственно замедляется проведение импульса. Потенциал действия со сниженной фазой 0 за счет инактивации быстрых Na каналов мембраны отражает «подавленный быстрый ответ». При уменьшении потенциала покоя до уровня —50 мВ инактивируется около 50% Na каналов, и возбуждение (проведение) становится невозможным. Возникающие блокады могут быть как однонаправленными, так и двунаправленными.
|
|
|
В части случаев, даже при значительном уменьшении потенциала покоя, проведение импульса сохраняется, правда, резко замедленное. Воз-
буждение клеток, т. е. регенеративная деполяризация мембраны, обеспечивается в этих условиях медленными входящими Са++- и Na+-TOKa-ми, поскольку медленные Са, Са — Na и Na каналы мембраны устойчивы к снижению потенциала покоя. «Быстрые» клетки превращаются в «медленные» клетки с выраженным торможением скорости проведения или с возникновением блокады.
Одной из форм нарушенной проводимости является декрементное (затухающее) проведение, т. е. прогрессирующее замедление проведения в сердечном волокне, по длинни-ку которого постепенно снижается эффективность стимула (ПД) и (или) возбудимость ткани. Этот процесс обычно развертывается в функционально подавленных волокнах, на что обратил внимание еще J. Erlanger (1906), впервые выдвинувший концепцию о декрементном проведении. В 1928 г. F. Schmitt и J. Erlanger предложили модель декрементного проведения: на длинную мышечную полоску, вырезанную из желудочка черепахи, они воздействовали раствором, содержавшим высокую концентрацию ионов К+. Вероятно, такой же характер носят нарушения проводимости в волокнах Пуркинье, сохранившихся после экспериментального инфаркта миокарда, а также блокады в местах соединения волокон Пуркинье с мышечными сократительными волокнами [Bigger J., 1980; Gil-mour R. et al., 1985].
Другая форма нарушенной проводимости представлена неравномер ным проведением. Если в параллельно расположенных сердечных волокнах проведение становится декре-ментным, но не в одинаковой степени, то вместо единого фронта возбуждения появляются опережающие и запаздывающие волны. Деполяризация соседних волокон происходит неодновременно, общая эффективность стимула падает, и возможно развитие частичной или полной блокады его проведения.
В 80-х годах рядом авторов подчер-
кивается значение неравномерностеи в распределении межклеточных соединений для возникновения нарушений проводимости [Spach M. et al., 1981, 1982, 1986]. Основанием для такого вывода послужили данные об анизотропности сердечной мышцы и связанным с ней более быстрым проведением импульса вдоль, чем поперек мышечного волокна. Соответственно, общее время открытия Na каналов оказывается более продолжительным, когда импульс проводится вдоль волокна, чем поперек его. Такого же рода неравномерности проявляются в тех участках, где мышечные пучки разветвляются пли соединяются с другими пучками, что и в норме приводит к внезапному замедлению ПД. В патологических условиях при ослаблении деполяризующего тока или межклеточных связей здесь могут возникать блокады.
Гипотеза, развиваемая G. Мое и сотр. [Antzelevitch С., Мое G., 1981; Antzelevitch С. el al., 1985], основывается на многочисленных экспериментальных данных, показывающих, что электротоническое взаимодействие между двумя возбудимыми участками, разделенными небольшой зоной высокого сопротивления, сопровождается резким замедлением проводимости в дистальном участке волокна. Такие факторы, как ограниченная ишемия миокарда, местная высокая концентрация ионов К+, локальное сдавление или охлаждение и другие воздействия могут вызвать невозбудимость небольшого сегмента в сердечном волокне (волокне Пуркинье) и тем самым способствовать электро-тонически опосредованному ступенчатому торможению передачи импульса через невозбудимую зону. По мнению С. Antzelevilch и соавт. (1985), этот механизм играет даже более важную роль в развитии частичных или полных блокад в сердце человека, чем изменения амплитуды ПД или скорости возрастания его фазы 0 (электрический ответ).
Мы кратко рассмотрели особенности механизмов блокирования им-
пульсов. При изложении отдельных форм аритмий будет уделено внимание таким нарушениям, как скрытое проведение, однонаправленные блокады и др. Непосредственное отношение к расстройствам проводимости имеет и повторный вход импульса (re-entry).
ПОВТОРНЫЙ ВХОД ИМПУЛЬСА (RE-ENTRY) И ЕГО КРУГОВОЕ ДВИЖЕНИЕ (CIRCUS MOVEMENT)
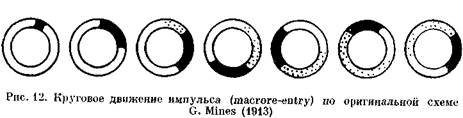
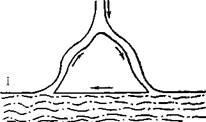
Как возможный механизм сердечных аритмий re-entry было распознано еще в начале XX в. [Мауег А., 1906, 1908; Mines G., 1913, 1914; Car-rey W., 1914]. Этим термином обозначают явление, при котором импульс, совершающий движение по замкнутому пути (петле, кругу, кольцу), возвращается к месту своего возникновения и повторяет движение (рис. 12). Фундаментальное изучение re-entry было предпринято F. Schmitt, J. Erlanger (1928) в уже упоминавшихся нами опытах с мышечной полоской из желудочка черепахи, подвергнутой воздействию давления, холода или раствора с высокой концентрацией ионов К+. Авторы, в частности, предположили, что избыток ионов К 1~ в наружной среде вызывает продольное разделение мышцы на два пути с антероградной блокадой проведения по одному из них. Искусственный стимул распространяется аптероградно по другому пути, а затем ретроградно продвигается по ранее блокированному пути к месту стимуляции. Это было первое четкое упоминание о возможности однонаправленного блокирования. Схемы повторного входа, предложенные исследователями для разветвленного и неразветвленного волокна, воспроизводятся с небольшими изменениями на рис. 13.
F. Schmitt, J. Erlanger (1928) указали также, что аналогичный процесс циркуляции может возникать в серд-
|
|
|
|
|
|

|
|
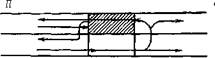
Рис. 13. Оригинальные схемы re-entry, пред- Рис. 14. Схема re-entry вокруг анатомиче-
ставленныо F. Schmitt, J. Erlanger (1928) ского препятствия: macrore-entry no M. Al
l-повторный вход в разветвленных мышечных lessie (объяснение в тексте),
волокнах; II—повторный вход в синцитиальной
структуре мышечной полоски.
це млекопитающих в очень небольших петлях, т. е. в форме microre-en-try. В экспериментальных работах, выполненных в 70-х годах, эта гипотеза получила подтверждение [Crane-field P., Hoffman В., 1971; Crane-field P. et al., 1971; Wit A. el al., 1972; Sasyniuk В., Mendez С., 1973]. Например, вызванное концентрированным раствором ионов К+ торможение скорости проведения в волокнах Пуркинье собаки до 0,01—0,1 м/с и укорочение в них периода рефрак-терности сопровождаются уменьшением кольцевого пути повторного входа до очень небольших размеров (^1 мм). Длина волны возбуждения, равная произведению из скорости проведения на длительность рефрактерности, соответствует величине такой минимальной петли reentry.
Современные представления о reentry усложнились, но они по-прежнему основываются на классических данных. Различают: a) macrore-entry (макрориентри), или упорядоченное
(ordered) re-entry; б) microre-entry ( микрориентри ), или «случайное» (random) re-entry. Разумеется, при таком делении учитывают размеры петли (круга), в которой осуществляется повторный вход. Однако не меньшее значение имеют электрофизиологические особенности каждого из этих двух подвидов re-entry. Мы приводим их описание, основываясь на результатах известных экспериментальных исследований М. Allessie и сотр. (1974—1984).
Для формирования macrore-entry с характерными для него свойствами требуются определенные условия:
а) наличие устойчивой замкнутой петли, длина которой зависит от пе риметра анатомического невозбудимого препятствия, вокруг которого движется импульс (рис. 14);
б) однонаправленная блокада проведения в одном из сегментов петли re-entry [Quan W., Rudy Y., 1990];
в) длина движущейся волны возбуждения должна быть короче длины петли; благодаря этому перед
фронтом («головой») распространяющегося по кругу импульса всегда имеется участок ткани, вышедший из состояния рефрактерности и восстановивший свою возбудимость; этот сегмент, или «окно возбудимости», имеет протяженность до 20% длины всей петли. Именно в этот «зазор» стараются попасть, нанося экстрастимулы, чтобы прервать круговое движение импульса при реципрокных тахикардиях. Укорочение рефракторного периода клеток, образующих петлю, способствует расширению «окна возбудимости», но оно не оказывает влияния на скорость распространения импульса и частоту ритма. Умеренное удлинение периода рефрактерности суживает «окно возбудимости» тоже без воздействия на скорость движения импульса по петле и на частоту ритма. При значительном удлинении рефрактерности «окно возбудимости» может закрыться, циркулирующая волна наталкивается на участок, находящийся в состоянии функциональной рефрактерности; движение импульса резко замедляется либо прекращается [Feld G. et al., 1986]. Описанный механизм mucrore-entry лежит, как полагают, в основе трепетания предсердий, а также некоторых форм реци-прокной тахикардии [Медвинский А. Б., Перцов А. М., 1989].
При другой разновидности повторного входа — microre-entry — движение импульса происходит по малому замкнутому кольцу, не связанному с каким-либо анатомическим препятствием. Теоретическое обоснование этому процессу было дано еще Th. Lewis (1925). М. Allessie и соавт. производили картографирование левого предсердия кролика в тот момент, когда с помощью электрического экстрастимула была вызвана пред-сердная тахикардия с частотой от 400 до 800 в 1 мин. Регистрацию возбуждения в различных участках предсердия производили как по его периметру, так и по радиусу. Можно было видеть, что импульс совершал не только круговое, но и цептростреми-
|
|

Рис. 15. Схема re-entry no малому кругу, не связанному с анатомическим препятствием — leading circle no M. Allessie (объяснения в тексте).
тельное движение в разных направлениях. По мере приближения к центру амплитуда и скорость подъема фазы О ПД понижались, и возбуждение затухало. Клетки в центре циркулировавшей волны давали только локальный электрический ответ, поскольку они поддерживались в рефракторном состоянии под воздействием поступавших с разных сторон импульсов. Место схождения этих импульсов служило функциональной основой для циркуляции волны возбуждения. Оно как бы заменяло анатомическое препятствие и защищало возбуждение от шунтирования (рис. 15).
Такое явление можно сравнить с водоворотом и воронкой в его центре. М. Allessie и соавт. (1977, 1980) назвали эту движущуюся, вращающуюся систему термином leading circle (leading circuit), т.е. «ведущий кружок» (цикл), или ведущая петля microre-entry, которая и определяет частоту возбуждения миокарда предсердий.
В предсердиях может быть несколько таких кругов, и самый меньший из них оказывается ведущим, так как в круге с наименьшим диаметром будет и самое короткое время обращения волны возбуждения. При столь малых размерах круга стимулирующий эффект движущейся волны оказывается достаточным, чтобы возбудить лежащий впереди участок миокарда, еще не вышедший из состояния функциональной рефрактерности. Другими словами, в кольце microre-entry нет «окна», т. е. зоны полностью восстановленной возбудимости; «голова» волны непосредственно следует за ее «хвостом». Длина ведущего круга оказывается равной длине волны возбуждения.
| 36 |
|
|
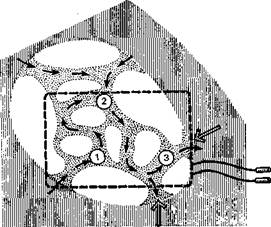
Рис. 16. Схема сложного многокольцевого re-entry в зоне инфаркта миокарда (по N. El-Sherii' и соавт.).
Ниже суммированы основные свойства leading circle:
а) размеры ведущего круга не являются фиксированными, они определяются длиной волны возбуждения, которая, в свою очередь, зависит от длительности функционального рефрактерного периода (ФРП) мышечной ткани и от скорости проведения в ней импульса; укорочение ФРП или замедление скорости проведения ведет к сужению (уменьшению) ведущего круга; когда же ФРП удлиняется, а скорость проведения возрастает, ведущий круг увеличивается в размерах; как видно, активность малого круга детерминируется не его длиной, а электрофизиологическими свойствами мышечных волокон, образующих этот круг;
б) в ведущем круге отсутствует участок, полностью восстановивший свою возбудимость; воздействовать на такой круг можно только с помощью стимула, сила которого значительно превышает диастолический порог возбуждения миокарда;
в) частота ритма, вырабатываемого в ведущем круге, обратно пропорциональна длительности ФРП: при его укорочении число импульсов в единицу времени возрастает.
Необходимо упомянуть еще об одном механизме — re-entry в неразветвленном волокне. Речь идет об {(отраженном повторном входе-,) (reflected re-entry). Развивающие эту
концепцию J. Jalife, G. Мое (1981), С. Antzelevitch и соавт. (1985) разработали экспериментальную модель, основу которой составляет электрото-нически опосредованное замедление проводимости. В неразветвленном волокне Пуркинье создается узкая зона (2 мм) функциональной невозбудимости, через которую осуществляется медленное элсктротоническое движение импульса от проксималь-ного к дистальному участку волокна. Если время этого антероградного движения велико, то создаются условия для электротоничсского тока в ретроградном направлении с повторным возбуждением проксимального участка волокна, вышедшего из состояния рефрактерности. Таким образом, импульс движется вперед и назад через один и тот же функционально блокированный сегмент благодаря электротонической передаче, а не вследствие продольного разделения волокна на 2 канала, как предусматривается в более старых моделях.
Дата добавления: 2019-02-22; просмотров: 163; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!