Первые шаги вверх по лестнице
Производство энергии в митохондриях позволяет эукариотическим клеткам быть больше бактерий: в среднем примерно в 10 тысяч — 100 тысяч раз. Большой размер приносит с собой энергетическую эффективность. В определенных пределах (которые, вероятно, определяются эффективностью «распределительных сетей») чем больше, тем лучше. Эта непосредственная выгода, проистекающая из непосредственного преимущества, вероятно, уравновешивает непосредственные недостатки большого размера — необходимость иметь больше генов, больше энергии и улучшенную организацию. Эта выгода, возможно, помогла эукариотам сделать первые шаги вверх по лестнице восходящей сложности.
Пара загадок все еще не дает мне покоя, но, думаю, они объяснимы. Во-первых, часто говорят, что энергетическая эффективность не может быть мишенью естественного отбора, потому что большим животным нужно больше еды. Энергетическая экономия проявляется только на уровне отдельных клеток или в пересчете на грамм массы. Критики теории не преминут заметить, что естественный отбор, как правило, действует на особей, а не на граммы массы. Не спорю, но окружающая среда и потребности организма все же связаны с размером тела. Мы видели, что крыса в семь раз прожорливее человека: в пересчете на единицу массы ей нужно в семь раз больше пищи, чем нам. Однако она не сильнее и не быстрее «относительно» своей окружающей среды. Термин «относительно» в данном случае имеет вполне реальное наполнение. Ясно, что крыса не может убить буйвола, а мы можем, как можем мы поймать и убить любое другое животное. Мир животных определяется их размером, и в нашем собственном мире нам нужно в семь раз меньше пищи в день, чем крысам. Соответственно, мы можем в семь раз дольше обходиться без пищи и воды. Масштаб нашего преимущества даже более очевиден, если учесть, сколько мы съедаем относительно собственной массы. Мышь, например, должна ежедневно съедать половину массы тела, иначе она умрет от голода, а ежедневно мы потребляем всего 2 % собственной массы. По-моему, это явное преимущество. Я не говорю, что преимущество размера всегда оказывается решающим. Маленький размер нередко выгоден, что нашло отражение в разнообразных эволюционных тенденциях. Однако энергетическая эффективность, связанная с большим размером, несомненно, оказала глубокое влияние на эволюцию эукариот.
Вторая загадка — это всеобъемлющий характер энергетического преимущества. Ранее в части 4 мы говорили в основном о млекопитающих и пресмыкающихся. Мы разложили энергетические сбережения на составляющие и пришли к выводу, что они предоставляют организму реальные возможности, а не следуют из ограничений фрактальной сети. С другой стороны, мы видели, что бактерии ограничены соотношением площади поверхности к объему, и это действительно ограничение, а не возможность. Есть ли преимущество размера у отдельных эукариотических клеток, например у амеб? Или у деревьев, или у креветок? Может быть, похоронив универсальную константу, мы потеряли и право делать какие-либо обобщения за пределами класса млекопитающих?
Не думаю. Я пока не приводил других примеров, потому что ответы на них менее очевидны, ведь млекопитающим и пресмыкающимся уделяли меньше внимания, чем другим животным. Тем не менее подозреваю, что большинство организмов, включая одноклеточных, получают те же выгоды. У более крупных организмов эти выгоды связаны со знакомым нам явлением — оптом дешевле. Как и в обществе, выгода зависит от затрат на установку и эксплуатацию оборудования, торговых издержек и т. д. Все они налагают ограничения на преимущества размера, но в пределах этих ограничений выгода все равно существенна. Дело в том, что живые организмы очень консервативны в принципах своего функционирования. В частности, они всегда имеют модульную организацию. Как отдельные клетки, так и многоклеточные организмы представляют собой мозаику функциональных частей. У многоклеточных дыхание или очищение от шлаков происходит в органах, а у одноклеточных — в органеллах, например в митохондриях. Модульные функции в пределах клетки — это, например, генетическая транскрипция, синтез белков, синтез мембран, закачивание солей, переваривание пищи, распознавание сигналов и реакция на них, производство энергии, перемещение, транспорт молекул и т. д. Думаю, что экономить за счет масштаба на уровне клеток можно не хуже, чем у многоклеточных организмов.
Настало время вернуться к уже затронутой проблеме числа генов. Мы говорили, что сложным организмам нужно больше генов, и обдумали гипотезу Марка Ридли о том, что накоплению генов и усложнению способствовало появление полового процесса. Однако мы видели, что ключ к усложнению — это, скорее всего, не половой процесс, во всяком случае, не он ограничивает число генов у бактерий и одноклеточных эукариот. Интересно, нельзя ли объяснить увеличение числа генов у эукариот с точки зрения энергетической эффективности более крупных клеток? Большие клетки, как правило, имеют большое ядро. Не исключено, что для сбалансированного роста во время клеточного цикла нужно примерно постоянное соотношение объема ядра к объему клетки (еще одна степенная зависимость!).
Это означает, что в процессе эволюции размер ядра, а с ним и содержание ДНК, подстраивается к изменениям объема клетки для обеспечения оптимального функционирования. Значит, по мере увеличения размеров клеток их ядро тоже увеличивается и в нем становится больше ДНК, хотя эта лишняя ДНК необязательно кодирует большее число генов. Это объясняет C-парадокс, который мы обсуждали в главе 1, а также то, почему Amoeba dubia имеет в 200 раз больше ДНК, чем человек, хотя эта ДНК кодирует меньше генов.
Эту лишнюю («избыточную») ДНК часто считают мусором, и действительно, она может быть чисто структурной, но может и выполнять какую-то полезную функцию — от структурной поддержки хромосом до обеспечения участков связывания, регулирующих активность многих генов. Избыточная ДНК также служит сырьем для новых генов, закладывая основы сложности. Последовательности многих генов свидетельствуют, что они произошли от избыточной ДНК. Может быть, происхождение сложности было связано с такой простой вещью, как пропорциональное изменение размера? Как только у эукариотических клеток появились митохондрии, у большого размера появилось селективное преимущество. Большим клеткам нужно больше ДНК, а с ней пришло сырье для большего числа генов и большей сложности. Обратите внимание, что эта ситуация прямо противоположна ситуации у бактерий: жесткое давление отбора заставляет бактерии терять гены, а эукариотам, наоборот, выгодно их приобретать. Если Ридли прав, и половой процесс — способ отсрочить мутационный коллапс, то необходимость иметь больше ДНК при увеличении размера было тем самым давлением отбора, которое и привело к появлению пола.
Для эукариотических клеток обладание митохондриями расширило жизненные возможности. Благодаря им большой размер из абсолютно невероятного стал вполне возможным, что перевернуло с ног на голову ограничения мира бактерий. С большим размером пришла и большая сложность. Но были и недостатки, связанные с конфликтом между митохондриями и содержащей их клеткой. Последствия этой долгой битвы тоже оказались глобальными, оставив на жизни неизлечимые глубокие шрамы, но даже эти шрамы обладали и созидательной и разрушительной силой. Без митохондрий не было бы клеточного самоубийства, но не было бы и многоклеточных особей; не было бы старения, но не было бы и разных полов. У темной стороны митохондрий было даже больше возможностей переписать сценарий жизни.
Часть 5
Убийство или самоубийство?
Трудное рождение индивидуума
Когда клетки тела изнашиваются или повреждаются, они погибают в результате насильственного самоубийства — апоптоза. Клетка распадается на части, они упаковываются и перевариваются. Нарушение механизмов апоптоза приводит к раку — конфликту интересов отдельных клеток и организма в целом. Видимо, апоптоз необходим для обеспечения целостности многоклеточных организмов, но почему независимые клетки согласились на гибель ради высшего блага? Сегодня апоптоз контролируется митохондриями, которые унаследовали машину смерти от своих предков-бактерий. Так неужели целостность индивидуума действительно родилась в борьбе не на жизнь, а на смерть?
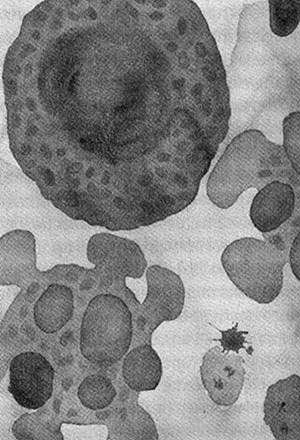
Смерть от апоптоза: жить клетке или погибнуть, решают митохондрии
«Я мыслю, следовательно, я существую», сказал Декарт. Так и хочется спросить: «Но что такое я?» Природа индивидуума, долгое время ускользавшая от философов и ученых, начала проясняться только недавно. Можно сказать, что индивидуум — это организм, состоящий из генетически идентичных клеток, которые специализированы для выполнения разных задач на благо организма в целом. С эволюционной точки зрения вопрос стоит так: почему эти клетки обуздали свои эгоистичные интересы и альтруистично сотрудничают в составе организма? Конфликты на разных уровнях организации — между генами, органеллами и клетками — неизбежны, но, как ни парадоксально, без этих междоусобных войн, возможно, никогда не возникли бы прочные связи клеток в пределах особи. Такие конфликты подхлестнули эволюцию молекулярной «полиции», которая обуздывает эгоистичные интересы примерно так, как правосудие обеспечивает приемлемое поведение членов общества. Центральным моментом полицейского контроля в организме является программируемая клеточная смерть — апоптоз. Апоптоз регулируется митохондриями, что наводит на мысль о том, что именно они сыграли ключевую роль в возникновении индивидуума. В этой части книги мы увидим, что в туманной глубине эволюционных времен митохондрии действительно были тесно связаны с возникновением многоклеточных особей.
По поводу эгоистичных генов, альтруизма и пределов естественного отбора было излито немало яда. В основе споров, заставивших научное сообщество позабыть о приличиях, лежал простой вопрос: на что действует естественный отбор? Действует ли он на гены, особей, группы особей (например, родственников) или на вид в целом? Внимание к этой проблеме привлекла прекрасно написанная работа Веро Винн-Эдвардса «Распространение животных в связи с общественным поведением» (1962 г.). Он объяснял многие аспекты общественного поведения не отбором на уровне особей, как считал Дарвин, а отбором на уровне вида. Поведение — это только вершина айсберга. Казалось, с такой точки зрения легче объяснить и многие другие особенности. Например, старение не приносит особи никакой пользы (какой нам прок от старости и смерти?), но вполне полезно для вида в целом, так как приводит к обновлению популяции, предотвращая скученность и ограничивая потребление ресурсов. Отдельной особи нет никакой пользы и от существования двух полов, так что в качестве взятки к полу прилагается острое эротическое наслаждение (просто удовольствия, надо полагать, было бы недостаточно). Чтобы произвести на свет одного потомка, при половом размножении часто нужны два родителя, то есть затраты вдвое больше, чем при делении пополам, и это не говоря о сложностях, связанных с поиском партнера. Хуже того, половой процесс приводит к случайному перераспределению тех самых генов, которые обеспечили успех родителей, и, следовательно, является потенциальной обузой. Его очевидная ценность заключается в быстром распространении изменчивости и полезных адаптаций в популяции, а это выгода для вида в целом.
Реакцию на эти идеи часто называют ультрадарвинизмом, но это пренебрежительное клеймо несет мало смысла. Как, спрашивается, работает отбор на уровне вида? Можно назвать несколько возможных путей. Например, быстрый оборот популяции может приводить к повышению темпов эволюции, что может быть выгодно для одного вида в противовес другому в быстро меняющихся условиях (например, во время глобального потепления или после удара метеорита). Еще одна возможность, которую Ричард Докинз называет «эволюция способности эволюционировать», связана с генетической «гибкостью» вида. В некоторых видах заложены большие способности к эволюции, выраженные в их морфологии и поведении, чем в других. В большинстве случаев, однако, «слепота» эволюции означает, что отбор на уровне вида просто не может работать. Пол — сложное явление, возникшее не за один день. Если преимущества от существования полов имеет только вид как целое, и если эти преимущества начинают проявляться только тогда, когда пол сформировался окончательно, что происходит, пока этот признак (пол) формируется? Те особи в популяции, которые сделают первый шаг в сторону приобретения пола, будут уничтожены естественным отбором прежде, чем получат какие-либо преимущества, потому что будут нести двойные затраты, связанные с полом, и страдать от случайного перераспределения полезных признаков. Или вот другой пример: нестареющие особи передадут потомству свои препятствующие старению гены, и те будут доминировать в популяции просто потому, что их носители имеют больше времени на передачу их потомкам. Таким образом, с одной стороны, казалось, отбор может работать на уровне вида лишь ограниченным числом способом, а с другой — что некоторые «самоотверженные» признаки могли возникнуть только за счет отбора на уровне вида.
Начиная с 1960-х гг., Уильям Хамильтон, Джордж К. Уильямс, Джон Мейнард Смит и другие пытались объяснить альтруистичные признаки отбором на уровне особей, родственных групп или генов. Новый подход свелся к математическим исследованиям совокупной приспособленности , суть которой хорошо суммировал Джон Б. С. Холдейн в спорах за кружкой пива: «Готов ли я пожертвовать жизнью ради спасения брата? Нет, но я готов ею пожертвовать ради спасения двух братьев, четырех племянников или восьми двоюродных братьев». (Обоснование: 50 % генов у него общие с братьями, 25 % — с племянниками и 12,5 % — с двоюродными братьями, так что распространенность его генов, по крайней мере, не понизится). Немало желчи изливалось на такие нагруженные смыслом термины, как «эгоистичный». В биологии они имеют специальное определение, но в повседневной жизни насыщены эмоциональными обертонами. Книга Ричарда Докинза «Эгоистичный ген» вдохновляла или, напротив, приводила в бешенство целое поколение, и во многом потому, что была так блестяще написана. Каждый читатель ощущал беспощадную логику вывода: живые организмы — одноразовые машины выживания их генов, смертные марионетки в руках практически бессмертных кукольников-генов. Единственный здравый подход к эволюции, говорит Докинз, предполагает, что мы должны прекратить заниматься самолюбованием и посмотреть на динамику популяций с точки зрения генов.
Мысль о том, что ген является «единицей отбора», критиковалась с самых разных сторон. Чаще всего критики утверждали, что гены невидимы для естественного отбора: они представляют собой инертные куски телеграфной ленты, которые всего лишь кодируют белки или РНК. Более того, отношения между геном и белком, который им кодируется, неоднозначны. Один и тот же ген можно разбить на части по-разному, так что он будет кодировать несколько разных белков; кроме того, теперь мы знаем, что многие белки выполняют не одну, а много функций. В ряде случаев один и тот же ген может оказывать прямо противоположное действие. Классический пример: один и тот же ген предохраняет от малярии, если присутствует в половинной дозе (в гетерозиготном состоянии), но вызывает серповидно-клеточную анемию в двойной дозе (в гомозиготном состоянии). Все это так, но ничего из сказанного не умаляет способности ориентированного на гены подхода объяснять эволюционные тенденции. Объектом отбора может быть особь, но только гены передаются следующему поколению. Ключ к понятию «эгоистичный ген» в том, что при половом размножении ни особь, ни ее отдельные клетки, ни даже хромосомы не переходят в следующее поколение. Тела исчезают и возникают вновь, недолговечные и неповторимые, как облака. Только гены, словно древние горы, пребывают в веках. С точки зрения популяции в эволюционном времени изменения частот генов — лучший способ количественной оценки эволюции. В некоторой степени это математическая подпорка сложной проблемы, но это также и правда, какой бы горькой она ни была.
С точки же зрения эгоистичных генов возникновение индивидуума — не проблема. Если конгломерат клеток, который мы называем телом, передает свои гены следующему поколению, то эти гены будут успешны, а гены, обладатель которых не смог их передать, не будут. Тело — продукт генов, сотрудничающих ради собственных эгоистичных интересов, а именно — как можно более широкого копирования. Докинз говорит об этом совершенно ясно: «Некоторые люди прибегают к метафоре, говоря, что тело — это колония клеток. Я предпочитаю считать тело колонией генов, а клетку — удобным рабочим участком химической промышленности генов»[58].
Суть проблемы эгоистичного гена в том, что только ген передается из поколения в поколение. Поэтому он является самой стабильной эволюционной единицей: ген — это «репликатор». Докинз поясняет, что это относится только к организмам, размножающимся половым путем, то есть к большинству эукариот, хотя и не ко всем. Применительно к бактериям эта концепция не имеет такой же силы, потому что они размножаются бесполым путем. В данном случае можно сказать, что отдельная клетка переходит из поколения в поколение, а накопление мутаций означает, что сами гены все же меняются. Собственно говоря, в стрессовых условиях бактерии могут даже ускорять частоту мутаций в генах. Поэтому не совсем понятно, на что действует отбор у бактерий — на гены или на клетку в целом. Во многих отношениях именно клетка является репликатором.
Мутации необязательно меняют фенотип (внешний вид или функцию организма), но они по определению меняют сам ген, и даже могут до неузнаваемости исказить его последовательность, хотя для этого должны пройти эпохи. Мутации накапливаются потому, что многие из них не влияют или почти не влияют на функции, и поэтому естественный отбор «не замечает» их (нейтральные мутации). Большинство генетических различий между людьми, в среднем одно различие на каждые 1000 «букв» ДНК (всего — миллионы букв), скорее всего, являются результатом нейтральных мутаций. Когда мы рассматриваем очень разные виды, две последовательности могут различаться настолько, что практически невозможно усмотреть в них какое-либо родство, если не принять во внимание весь спектр промежуточных форм (более близкородственных видов). Тогда можно увидеть, что якобы неродственные гены на самом деле находятся в состоянии родства. Физическое строение и функции белков, кодируемых абсолютно непохожими генами, часто удивительно хорошо сохраняется в процессе эволюции, несмотря на существенные изменения аминокислотных компонентов. Очевидно, строение и функции белка подвергаются действию отбора, в то время как нуклеотидная последовательность гена может быть относительно пластичной. Представьте, что вы вернулись в компанию, где работали прежде: никто из ваших бывших коллег там уже не работает, но организационно-правовая форма, административные структуры и общий дух остались точно такими, какими вы их помните.
Поскольку гены могут меняться, а клетка и ее составные части, по сути, не меняются, бактериальную клетку можно считать более устойчивой эволюционной единицей, чем ее гены. Например, цианобактерии («изобретатели» фотосинтеза), несомненно, модифицировали свои генетические последовательности с течением эволюции, но, как говорят ископаемые свидетельства, за миллиарды лет почти не изменились внешне. Если, как утверждал Докинз, худший враг эгоистичного гена — эта конкурирующая (полиформная, или измененная) форма того же гена, то нейтральные мутации, искажая гены, путают им все карты. По мере накопления нейтральных мутаций генетические последовательности расходятся. Один и тот же ген может иметь миллионы разных форм (у разных видов), и все эти формы в той или иной степени искажены. Этот факт лежит в основе любых построений, основанных на расхождении генов. Итак, эволюция противопоставляет случайную силу мутаций, которые все время искажают генетические последовательности, превращая эгоистичный ген в его злейшего врага и эгоистичные интересы генов, которые «хотят» точно копировать самих себя.
О том, что ген не является «единицей отбора» у бактерий, свидетельствует еще целый ряд фактов. Говорят, что при клональном размножении все гены передаются «скопом», так что их судьба является общей с судьбой клетки. Это не совсем так. Во-первых, бактерии меняются генами, а во-вторых, они часто оказываются жертвой вирусов-бактериофагов, которые «заряжают» их эгоистичной ДНК. Тем не менее, в то время как эукариоты напичканы «паразитической» ДНК («эгоистичными» последовательностями, бесполезными для организма в целом), бактерии имеют маленькие геномы и почти лишены «паразитической» ДНК. Как мы видели в части 3, бактерии теряют лишнюю ДНК, включая функциональные гены, потому что это ускоряет их размножение. «Эгоистичных» генов то и дело наказывают изгнанием в недружественную окружающую среду. Возможно, горизонтальный перенос генов у бактерий стоит считать диверсией со стороны эгоистичных генов, но, в общем, он продолжается только до тех пор, пока клетке нужны дополнительные гены, а потом они снова теряются наряду с любыми другими ненужными генами. Не сомневаюсь, что при желании все это можно истолковать с точки зрения теории эгоистичного гена, но, по-моему, такое поведение гораздо проще объяснить с точки зрения издержек и выгод самих клеток.
Есть еще один аспект, в котором именно клетку, по крайней мере клетку бактерий, логичнее рассматривать в качестве эгоистичной единицы. Дело в том, что гены кодируют не клетки, а аппарат, который их создает. Они кодируют белки и РНК, а они потом строят все остальное. Это не мелочная придирка, а существенная разница. Все клетки, даже бактериальные, имеют сложнейшую структуру, и чем больше мы их изучаем, тем лучше понимаем, что от нее зависит функционирование клетки. Мы уже убедились в части 2, что клетки — это не мешки с ферментами. Как ни странно, в генах, по-видимому, нет ничего, что кодировало бы структуру клетки. Например, мембранные белки направляются в конкретные мембраны за счет хорошо известных кодирующих последовательностей, но никакие последовательности не указывают, как создать такую мембрану с нуля, и не определяют, где ее нужно создать: липиды и белки добавляются к уже существующим мембранам. Сходным образом новые митохондрии всегда образуются путем деления старых; их нельзя создать на ровном месте. То же самое относится, например, к центриолям — тельцам, принимающим участие в формировании микротрубочек цитоскелета при делении клетки.
Это значит, что на фундаментальном клеточном уровне природа определяет воспитание, а воспитание — природу. Иными словами, гены целиком и полностью завязаны на существование клетки, а она может быть создана только за счет действия генов. Соответственно, гены всегда передаются в клетке, например в яйцеклетке или в бактерии, и никогда не передаются «отдельным пакетом». Вирусы, а они-то как раз и представляют собой «отдельный пакет», «оживают», только получив доступ к механизму существующей клетки. Вот как это сформулировал примерно двадцать лет назад выдающийся микробиолог Фрэнклин Харольд (мы с ним уже встречались в части 2 книги), и с тех пор почти ничего не изменилось:
«Геном — единственное вместилище наследственной информации, и именно он определяет форму, а окружающая среда лишь вносит мелкие корректировки. Но в попытках разобраться, как именно геном это делает, мы открываем одну китайскую коробочку за другой, а последняя коробочка в наборе оказывается пустой. Продукты генов поступают в уже имеющийся организованный матрикс, который состоит из образовавшихся раньше продуктов генов, а функциональная экспрессия генов зависит от места, где они оказываются, и от сигналов, которые они получают. Форма не прописана однозначно ни в одном из сигналов, но подразумевается сочетанием сигналов с определенным структурным контекстом. В конечном счете, только клетки делают другие клетки».
Подведем итог: есть много причин считать эгоистичной единицей эволюции бактериальную клетку, а не ее гены. Может быть, как говорил Докинз, с появлением пола у эукариот все изменилось, но если мы хотим понять глубинную подоплеку эволюции, мы должны присмотреться к бактериям, два миллиарда лет царившим на Земле.
Эти различия в подходах проясняют, почему самыми выдающимися критиками теории эгоистичного гена были микробиологи, например Линн Маргулис. На самом деле, Маргулис откровенно критиковала математический неодарвинизм в целом. Она даже сравнивала его с френологией — викторианской зацикленностью на связи между формой черепа и преступными наклонностями — и утверждала, что неодарвинизм придет к столь же бесславному концу.
Чувствуется, что концепция эгоистичного гена вызывает у Маргулис отвращение, но дело не только в этом. Бактерии действительно ведут себя более прилично: они не поедают друг друга, а образуют гармоничные сообщества. Укоренившееся представление о бактериях как злобных патогенах ложно. Маргулис считала, что эволюция, в общем, бактериальное дело, и ее можно объяснить сквозь призму взаимного сотрудничества бактериальных консорций, в том числе эндосимбиозов, включая тот, что дал начало эукариотической клетке. Эти консорции хорошо подходят бактериям, потому что хищническое поведение у них не окупается. Как мы видели в части 3, процесс дыхания через клеточную мембрану не дает появиться крупным бактериям, способным физически захватывать другие клетки. Для бактерий существенно, кто быстрее растет, а не у кого больше пасть. Учитывая, что в бактериальных экосистемах пищи всегда мало, бактериям выгоднее питаться экскрементами других бактерий, а не драться с ними за ресурсы. Если одна бактерия живет за счет сбраживания глюкозы с образованием молочной кислоты, другая может окислять молочную кислоту до углекислого газа, третья — превращать углекислый газ в метан, четвертая — окислять метан и т. д. Жизнь бактерий — постоянная переработка отходов производства, а это проще всего делать, объединившись.
Стоит припомнить, что даже кооперативные партнерства имеют шанс на долгосрочное существование только в том случае, если они более выгодны партнерам, чем независимая деятельность. Что бы мы ни имели в виду, говоря об успехе — выживание клеток или выживание генов, — виден лишь результат, то есть клетки или гены, которые оказались успешнее других. Клетки, альтруизм которых простирается настолько далеко, что они умирают ради других, исчезают без следа. Так многие юные герои, защищавшие свою страну и павшие в бою, были оплаканы родными, но сами не оставили потомства. Я имею в виду, что сотрудничество не всегда альтруистично. И все же взаимоотношения бактерий далеки от традиционного представления о животных нравах, которое выразил Теннисон в строке об окровавленных клыках и когтях природы. Может быть, среди бактерий и нет взаимопомощи, но нет и агрессии; по крайней мере, их нравы не наводят на мысль об окровавленном оскале.
Это различие между бактериальным и эукариотическим миром отчасти объясняет раскол между Маргулис и неодарвинистами, такими как Докинз. Как мы видели, идеи Докинза об эгоистичных генах звучат довольно сомнительно, если попробовать приложить их к бактериям (сам Докинз и не пытался этого делать). Однако Маргулис считает, что весь узорный покров эволюции сплетен бактериями. Они вошли в нашу плоть и кровь, определяя даже наше сознание за счет тонких сетей микротрубочек в мозге. На самом деле, вся биосфера представлялась Маргулис плодом сотрудничества бактерий, и это нашло отражение в «гипотезе Геи», предложенной ею вместе с Джеймсом Лавлоком. В своей последней книге «Приобретение геномов: теория происхождения видов», написанной в соавторстве с сыном, Дорионом Саганом, Маргулис утверждает, что даже у растений и животных новые виды образуются, как у бактерий, в процессе слияния геномов, а не в результате постепенного расхождения, как считал Дарвин и считают современные биологи. Возможно, теория слияния геномов справедлива в ряде конкретных случаев, но в основном она отступает под давлением тщательного эволюционного анализа последних ста лет. Отвергая неодарвинизм, Маргулис намеренно настраивает против себя большинство традиционных эволюционистов[59]. Мало кто из них выказывает такое терпение, как покойный Эрнст Майр, предпославший книге Маргулис мудрое предисловие.
В нем он хвалит изложенное в книге видение бактериальной эволюции, но осторожно предупреждает читателя, что идеи Маргулис неприменимы к подавляющему большинству многоклеточных организмов, включая все 9000 видов птиц, которыми долгие годы занимался он сам. Половое размножение означает, что гены конкурируют за место на хромосомах, а после возникновения хищничества у эукариот природа, как ни печально, действительно обзавелась окровавленными клыками и когтями.
Учитывая концептуальное расхождение в подходах, забавно, что, когда дело доходит до особи, взгляды Докинза и Маргулис не слишком отличаются. Как мы видели, Докинз считал индивидуум колонией сотрудничающих генов, а Маргулис — колонией сотрудничающих бактерий, которую можно считать колонией сотрудничающих бактериальных генов. Как Докинз, так и Маргулис считают индивидуум сущностью, основанной на сотрудничестве. Вот что пишет Докинз в замечательной книге «Рассказ предка»: «Моя первая книга, „Эгоистичный ген“, могла бы называться „Сотрудничающий ген“ — и ни слова в самой книге менять бы не пришлось. <…> Эгоизм и сотрудничество — две стороны дарвиновской медали. Каждый ген стремится к собственному эгоистичному благополучию, сотрудничая с другими генами в своей среде — пуле генов, перемешиваемом половым процессом, — с целью построения общих тел».
Однако эти идеи, ставящие во главе угла сотрудничество, затеняют конфликт между разными эгоистичными сущностями, из которых состоит особь, в частности между клетками и митохондриями. Хотя такой конфликт прекрасно согласуется с философией Докинза, в «Эгоистичном гене» он не стал развивать эту мысль. Она нашла отражение в следующей книге Докинза «Расширенный фенотип», а в 1980-х и 1990-х гг. — в работах биолога Лео Бусса и других ученых. Сосредоточив внимание на таких конфликтах и их разрешении, эволюционные биологи поняли, что колонии клеток (или, если хотите, генов) не являются истинными индивидуумами, а образуют менее прочную ассоциацию, в которой отдельные клетки все еще способны к независимым действиям. Многоклеточные животные, такие как губки, могут распадаться на части, а каждая часть может дать начало новой колонии. Общность цели преходяща, потому что судьба отдельных клеток не связана намертво с судьбой многоклеточной колонии.
У истинных особей такое своеволие жестоко подавляется. Эгоистичные интересы клеток подчинены общей цели. Для этого используются разные средства, в том числе раннее отделение клеток зародышевой линии. Оно приводит к тому, что подавляющее большинство клеток тела (так называемые соматические клетки) не передают потомству свои гены напрямую и могут, так сказать, лишь зачарованно следить за тем, как это делают клетки зародышевой линии. Подобный вуайеризм был бы невозможен, если бы все клетки тела не были бы связаны едиными генами, то есть не происходили бы от одной оплодотворенной яйцеклетки (зиготы) путем бесполого размножения. Клетки зародышевой линии тоже не передают свои гены непосредственно следующему поколению, но передают их точные копии, что тоже неплохо, а по большому счету вообще то же самое. Но политики пряника иногда недостаточно, и иногда приходится прибегать к политике кнута. Эгоистичные конфликты между самими клетками, пусть и генетически идентичными, может разрешить жесткое полицейское государство, сравнимое разве что с Советским Союзом сталинских времен. Нарушителей не преследуют по закону, а физически уничтожают.
Эта драконовская система приводит к тому, что естественный отбор перестает выбирать между независимыми сущностями, составляющими особь, и начинает действовать на новом, более высоком уровне, то есть выбирать между соперничающими особями. Однако тень былых раздоров лежит даже на самых крепких, на первый взгляд, телах. Она напоминает о том, что единство индивидуума было завоевано с трудом, а вот утратить его очень легко. Одной такой тенью прошлого является рак. Рак и уроки, которые можно из него извлечь, мы обсудим в следующей главе.
Конфликт в теле
Рак — жуткий призрак конфликта, раздирающего индивидуума изнутри. Одна клетка вдруг выходит из-под централизованного контроля и начинает размножаться на бактериальный манер. Последовательность событий на молекулярном уровне может служить одной из самых лучших иллюстраций естественного отбора в действии. Посмотрим же, что происходит при раке.
Часто, хотя и не всегда, рак возникает в результате генетических мутаций. Одной мутации, как правило, недостаточно. Клетка должна накопить от восьми до десяти мутаций в довольно специфичных генах. Только после этого она трансформируется в злокачественную клетку, которая ставит свои интересы выше интересов тела. Генетические мутации обычно накапливаются с возрастом случайным образом, но только особое сочетание приводит к возникновению рака: в основном мутации должны затрагивать две определенные группы генов: онкогены и гены-супрессоры опухолей. Эти гены кодируют белки, контролирующие нормальный клеточный цикл — то, как клетки растут или умирают в ответ на сигналы из определенных источников.
Продукты онкогенов в норме сигнализируют клетке, что она должна делиться в ответ на конкретный стимул (например, чтобы заменить мертвые клетки после инфекции), но при раке выключатель этих клеток «западает» в положении «включено». Напротив, продукты генов-супрессоров опухолей в норме сдерживают неконтролируемое деление клетки. Они аннулируют сигналы к пролиферации, заставляя клетку прекратить деление или принуждая ее к самоубийству. При раке они «западают» в положении «выключено». В клетках есть обширная система проверок и контроля, и именно поэтому нормальная клетка превращается в злокачественную клетку, как правило, только после накопления 8–10 мутаций. Некоторые люди с генетической предрасположенностью к раку наследуют часть мутаций от родителей, поэтому пороговое число «новых» мутаций, которые должны накопиться, прежде чем начнется рак, у них ниже.
Трансформированные клетки перестают откликаться на приказы тела. Размножаясь, они превращаются в опухоль. Не любая опухоль — это рак; разница между доброкачественной и злокачественной опухолью огромна. Прежде всего, чтобы диаметр опухоли превысил пару миллиметров, ей нужно питание. Опухолевым клеткам недостаточно медленного поглощения питательных веществ через поверхность, им нужно внутреннее кровоснабжение. Для этого они должны производить определенные химические посредники (факторы роста) в определенном количестве. Они будут стимулировать прорастание в опухоль новых кровеносных сосудов. Дальше опухоль должна переварить окружающие ткани, чтобы освободить себе пространство для роста. Для этого опухолевые клетки выделяют сильно-действующие ферменты, разрушающие тканевую структуру. Вероятно, самая страшная стадия процесса — переход к метастазированию, то есть к распространению опухоли в другие части тела. Для этого нужно определенное сочетание почти взаимоисключающих свойств. Клетки должны быть достаточно «скользкими», чтобы покинуть опухоль, но достаточно «липкими», чтобы закрепиться на стенках кровеносных сосудов в других частях тела. Они должны ускользнуть от внимания иммунной системы во время прохода по кровеносным или лимфатическим сосудам, и часто им это удается, потому что они маскируются, сбиваясь в кучки, несмотря на свою скользкую поверхность. Прибыв на место, клетки должны проникнуть через стенки сосудов в ткань и там остановиться. Наконец, на протяжении всего пути они должны сохранить способность к пролиферации, ведь только тогда они смогут основать аванпост рака в другом органе.
К счастью, лишь немногие клетки обладают диалектическими качествами, необходимыми для метастазирования. Тем не менее мало кто из нас не сталкивался с раком, если не лично, то у родных или знакомых. Как же клеткам удается приобрести необходимые качества? Дело в том, что раковые клетки возникают путем естественного отбора. На протяжении жизни клетки в ее генах происходят сотни мутаций, и некоторые из них затрагивают гены, контролирующие клеточный цикл. Тогда клетка сбрасывает оковы, в норме не дающие ей прорастать. Скоро это уже не одна клетка, а колония клеток, и все они мутируют. Многие мутации нейтральные, другие вредные, но со временем какие-то из них, а потом и следующие, подтолкнут клетку к злокачественному росту. Всякий раз потомки клетки пролиферируют: одиночный мутант дает начало популяции клеток, а потом ее вытесняет следующая клетка, адаптированная к следующему шагу. За несколько лет или даже месяцев рак захватывает тело. У раковых клеток нет будущего; как и мы сами, они обречены на смерть. Они просто делают то, что должны, а именно растут и меняются, покорные неумолимой логике изменчивости и отбора.
Что является единицей отбора при раке — ген или клетка? Как мы видели на примере бактерий, более логично считать эгоистичной единицей сами раковые клетки. Они размножаются не половым путем, а бесполым, как бактерии. Их гены могут меняться быстрее, чем фенотип: по внешнему виду раковой клетки под микроскопом, как правило, можно сказать, откуда она взялась. Исследовав опухоль в легком, как правило, можно сказать, является ли она первичной (то есть возникла из клеток легкого) или вторичной, то есть возникла из клеток другой ткани, например молочной железы, и переместилась в легкое за счет метастазирования. Во втором случае она сохраняет ряд атавистических признаков клеток молочной железы, например способность к производству гормонов. В то же время раковые клетки известны своей генетической неустойчивостью: их хромосомы теряются, «ломаются», причудливо сплетаются. Таким образом, раковые клетки некоторое время сохраняют прежнее обличье, а их гены меняются до неузнаваемости за счет мутаций и перетасовок. Если здесь и есть «эгоистичная» эволюционная единица, то это клетка, которая преодолевает все препятствия и в конце концов убивает хозяина — путь, предсказанный судьбой, почти как у Макбета.
В контексте рака слово «эгоистичный» звучит неубедительно. Злокачественная опухоль никоим образом не стремится к свободе. Она просто впадает в детство, возвращаясь в те времена, когда не было особи и все клетки делали что хотели. В этом смысле рак хорошо иллюстрирует бессмысленность эволюции. Клетки размножаются, а те, что размножаются успешнее всех, оставляют больше потомков. Точка. Трудно приписать раку какой-то высший смысл, это абсолютно бессмысленный процесс. Какой контраст с бактериальной инфекцией — еще одним хорошим примером эволюции в микрокосме! При размножении бактерий тоже складывается впечатление отлаженного механизма, но там четко просматривается цель. Мы можем ужасаться инфекциями, но бессмысленными их не назовешь: у бактерий есть жизненный цикл, задача, будущее. Они не обречены, ведь погубив одно существо, они заразят другие. (Конечно, это мнимая разница — на самом деле ни у бактерий, ни у раковых клеток «цели» нет, — но пример хорош. Раковые клетки не могут пережить тело, и поэтому преходящая природа их успеха особенно очевидна.)
Рак, при всей его бессмысленности, по крайней мере, хорошо иллюстрирует препятствия на пути к рождению индивидуума. Если сегодня раковые клетки могут чинить беззаконие, какая надежда была у первых многоклеточных индивидуумов? Клетки-дезертиры тогда имели шансы выжить в одиночестве. Как же первые индивидуумы подавили мятежные наклонности собственных клеток? Возможно, они, как и мы сегодня, убивали нарушителей за счет механизма программируемой клеточной смерти, или апоптоза, вынуждая клетки-диссиденты к самоубийству. Апоптоз есть даже у организмов, которые проводят часть времени как независимые свободноживущие клетки, а часть — в составе колонии. Встает вопрос: как и почему возник апоптоз у одноклеточных организмов? С какой стати потенциально независимая клетка «согласилась» на самоубийство?
Многое из того, что мы знаем об апоптозе, стало известно из исследований его роли при раке. Чем больше мы узнаем, тем лучше осознаем, что главную роль в апоптозе играют митохондрии. А если мы проследим путь этого процесса из глубины времен, то окажется, что апоптоз эволюционировал у первых эукариот из попыток митохондрий и содержащих их клеток-хозяев манипулировать друг другом, причем в те далекие времена, когда даже колониальные организмы были еще редкостью.
Хроника предсказанной смерти
У клеточной смерти есть два обличия: некроз — насильственная, неожиданная, моментальная гибель от удара ножа (агония, кровавые пятна на ковре), и апоптоз — заранее обдуманное самоубийство (порошок цианистого калия, тихий стук падающего тела). Апоптоз не оставляет улик, и такая манера разделываться с диссидентами вполне соответствует тоталитарному режиму организма. Напротив, смерть от некроза вызывает воспалительную реакцию — расследование поджога, в ходе которого полиция находит другие тела и которое еще долго вызывает общественные волнения.
Как ни странно, биологи далеко не сразу признали важность апоптоза. Биология, в конце концов, это изучение жизни, и в некотором смысле смерть как отсутствие жизни не входит в сферу ее интересов. Многие первые сообщения о программируемой клеточной смерти считались любопытными курьезами, не более. Одним из первых (в 1842 г.) апоптоз описал немецкий революционер, ученый и философ-материалист Карл Фохт, бежавший в Женеву от политического преследования. Несколько позже он стал мишенью блестящего политического памфлета Карла Маркса «Герр Фохт» (1860 г.) из-за своих отношений с Наполеоном III. Но не будем углубляться в политику, а вспомним лучше его скрупулезное исследование метаморфоза жабы-повитухи из головастика во взрослую особь. Фохт использовал микроскоп, чтобы проследить судьбу нотохорда — гибкого, примитивного позвоночника головастика. Его интересовал вопрос, превращаются ли клетки нотохорда в позвоночник взрослой жабы, или они исчезают, а позвоночник формируется за счет новых клеток? Выяснилось, что верен второй ответ: клетки нотохорда погибают (как мы теперь знаем, за счет апоптоза), а их место занимают новые клетки.
Другие ученые XIX в. тоже обращали внимание на апоптоз в связи с метаморфозом. Август Вайсманн, великий немецкий основоположник эволюционной биологии, заметил в 1860-х гг., что в процессе превращения гусеницы в бабочку многие клетки тихо погибают. Как ни странно, он не стал увязывать этот факт со старением и смертью, хотя именно исследования на эти темы впоследствии принесли ему известность. Большинство последующих описаний клеточной смерти тоже были сделаны эмбриологами и затрагивали изменения, происходящие во время развития. Был открыт удивительный факт: у эмбрионов рыб и цыплят погибают целые популяции нейронов. У нас, кстати, тоже. В некоторых областях мозга более 80 % нейронов, образующихся на ранних стадиях развития, исчезают еще до рождения! Клеточная смерть позволяет исключительно точно смонтировать электропроводку мозга: между конкретными нейронами образуются функциональные связи, а на их основе образуются нейронные сети. Многие эмбриологические процессы хочется уподобить ваянию из камня. Как скульптор отсекает ненужные кусочки от глыбы мрамора, чтобы создать прекрасную статую, так и тело созидается за счет изъятия, а не добавления. Наши пальцы, например, образуются за счет запланированной смерти клеток в промежутках между пальцами, а не за счет удлинения зачатков.
У лапчатых птиц, например уток, некоторые клетки в промежутках не погибают, и на лапах остаются перепонки.
Важность апоптоза у взрослых организмов оценили гораздо позже. Сам термин предложили Джон Керр, Эндрю Уайли и Аластер Керри (Университет Абердина) в 1972 г. Придумать его помог им Джеймс Кормак, профессор древнегреческого языка из того же университета («апоптоз» означает «опадание листьев»). Впервые этот термин был использован в названии их статьи в «Британском онкологическом журнале»: «Апоптоз как фундаментальный биологический феномен с множественными функциями в регуляции кинетики тканей». Кстати, Гиппократ обозначал словом «апоптоз» отмирание поврежденных фрагментов костей, а Гален называл так отпадание струпьев.
Джон Керр заметил, что размер печени у крыс не постоянен. Он динамически меняется с изменениями тока крови. Если поток крови в определенных долях печени нарушается, это компенсируется их уменьшением, а уменьшаются они по мере того, как клетки погибают за счет апоптоза (это занимает недели). Напротив, при восстановлении кровотока клетки начинают размножаться, и соответствующие доли постепенно увеличиваются в весе (время восстановления, опять же, исчисляется неделями). Такая эквилибристика происходит везде. В человеческом организме ежедневно умирают, заменяясь новыми, примерно 10 миллиардов клеток. Это не внезапная насильственная смерть, а тихая и незаметная смерть от апоптоза, когда соседние клетки подъедают все свидетельства гибели. То есть апоптоз уравновешивает клеточное деление. Из этого следует, что он не менее важен для нормальных физиологических процессов, чем деление клеток.
Керр, Уайли и Керри показали, что апоптоз протекает практически одинаково в совершенно разных обстоятельствах: при нормальном эмбриональном развитии и при тератогенезе (нарушениях формирования эмбриона), в здоровых тканях взрослого организма, при раке и при регрессии опухоли, а также при усадке ткани, связанной с неиспользованием или старением. Апоптоз также играет важнейшую роль в иммунном ответе организма. Клетки иммунной системы, действующие против тканей собственного тела, во время развития совершают апоптоз, что позволяет иммунной системе различить «свое» и «чужое». После этого клетки иммунной системы используют свои особенности для того, чтобы заставить поврежденные или зараженные клетки тоже совершать апоптоз. Такой скрининг позволяет уничтожить раковые клетки до того, как они начнут пролиферировать.
Последовательность событий при апоптозе отрежиссирована до мельчайших подробностей. Клетка сжимается, на ее поверхности появляются выросты, похожие на пузырьки. ДНК и ядерные белки (хроматин) конденсируются рядом с ядерной мембраной. Наконец, клетка распадается на апоптотические тельца (мелкие, окруженные мембраной структуры), а клетки иммунной системы подбирают их. По сути дела, клетка сама разделывает себя на кусочки, а другие клетки тут же поедают их; такой вот не афишируемый каннибализм. Учитывая все это, становится понятно, что для апоптоза нужна энергия. Клетка не может совершить апоптоз, если не располагает достаточным количеством АТФ. Этим апоптоз очень отличается от характерного для некроза набухания и разрыва клеток. Кроме того, в отличие от некроза, апоптоз не влечет за собой никаких последствий, например воспаления; то, что клетки больше нет, можно понять только по ее отсутствию. Предсказанная смерть тут же забывается.
Палачи
Более десяти лет «евангелисты апоптоза» — Эндрю Уайли и несколько других исследователей — пытались убедить научное сообщество в важности этого явления. Поначалу они сталкивались с непониманием и равнодушием. Уайли начал обращать других в свою веру после того, как показал, что при апоптозе хромосомы распадаются на сегменты с характерным «лестничным» паттерном, который можно выявить при биохимическом анализе. Теперь апоптоз можно было диагностировать в лабораторных условиях, и подозрения циничных биохимиков, что под видом открытия им пытаются преподнести артефакт электронной микроскопии, оказались беспочвенны. Но переломный момент наступил в середине 1980-х гг., когда Роберт Хорвитц (Массачусетский технологический институт) начал идентифицировать гены, отвечающие за апоптоз, у нематоды Caenorhabditis elegans (за это исследование он получил Нобелевскую премию 2002 г., разделив ее с Сиднеем Бреннером и Джоном Салстоном). Эта микроскопическая нематода как модельный объект имеет целый ряд крупных преимуществ. Во-первых, она прозрачная, так что судьбу отдельных клеток можно проследить под микроскопом. Во-вторых, во время эмбрионального развития путем апоптоза погибает небольшая, предсказуемая группа клеток (131 из 1090 соматических клеток, образующих организм нематоды). В-третьих, средняя продолжительность жизни С. elegans составляет всего 20 дней, и ее развитие нетрудно прослеживать в лабораторных условиях.
Хорвитц и его коллеги открыли несколько генов смерти, продукты которых провоцируют апоптоз у этой нематоды. Это было само по себе крайне интересно, но еще более неожиданным и важным оказалось другое открытие — точные эквиваленты генов смерти есть у мух, млекопитающих и даже растений. К тому времени исследователи рака уже выявили ряд этих генов, но какое отношение они имели к раку, было непонятно. Исследования, выполненные на нематодах, не только помогли прояснить их функцию, но и в очередной раз продемонстрировали фундаментальное единство жизни. Гены смерти человека оказались родственными соответствующим генам нематод, но это далеко не все. Оказалось, что, используя методы генетической инженерии, эти гены человека можно перенести в геном нематоды (заменив ее собственные гены), и они будут там прекрасно работать! Мутации, «выключающие» гены смерти, приводят к тому, что нематоды не теряют 131 клетку за счет апоптоза. Следствия данного открытия для изучения рака были очевидны: если у людей мутации этих генов приводят к тому же результату, то зарождающиеся раковые клетки вместо того, чтобы совершить самоубийство, будут размножаться с образованием опухоли.
К началу 1990-х гг. исследователи поняли, что некоторые онкогены и гены-супрессоры опухолей действительно контролируют судьбу клетки, влияя на апоптоз. Иными словами, рак возникает из-за клеток, потерявших способность к апоптозу, а они теряют эту способность после мутаций, затрагивающих гены смерти. К генам смерти относятся любые гены, который подталкивают клетку к апоптозу, и к ним могут относиться как онкогены, так и гены-супрессоры опухолей, потому что и те и другие могут отменить приказ совершить самоубийство. Как сказал в свое время Уайли, «билет до апоптоза прилагается к билету до рака, и чтобы добраться до рака, нужно сначала аннулировать билет до апоптоза».
За исполнение программы клеточной смерти отвечают особые палачи — белки каспазы. (Раньше эти белки назывались «цистеин-зависимые аспартат-специфические протеазы», но, по-моему, их современное название несколько более выразительно.) У животных описано больше десятка различных каспаз, и 11 из них есть у людей. Все они работают примерно одинаково: режут белки на кусочки. Некоторые из обрезков, в свою очередь, активируются и приступают к разрушению других компонентов клетки, например ДНК. Интересно, что каспазы не производятся на заказ по мере необходимости, а образуются постоянно, а потом ждут мобилизации в неактивном состоянии. Каспазы — это дамоклов меч, подвешенный над клеткой на тонкой нити. Бросает в дрожь от мысли, что эта тихая машина смерти затаилась практически во всех эукариотических клетках.
Все мы сидим под этим дамокловым мечом и должны радоваться, что нить, на которой он подвешен, довольно прочная. После активации каспаз назад дороги нет, но требуются многочисленные проверки и перепроверки, прежде чем древний механизм смерти придет в движение. В последние двадцать лет эта система контроля интенсивно изучается, однако разобраться в хаосе названий и сокращений может только крайне дотошный читатель. Ситуацию усложняет то, что по историческим причинам один и тот же ген называется по-разному у разных организмов. Это напоминает мне кельтскую музыку, где одна и та же мелодия часто известна под разными названиями, а одно и то же название относится к разным мелодиям. Бесконечные вариации на тему — это очень мило, но пониманию не способствует. Приведу пример: ген ced-3 нематоды называется nedd-2 у мыши, dcp-i у плодовой мушки-дрозофилы и ICE (интерлейкин-1-бета-конвертирующий фермент) у людей (так как сначала открыли, что он вовлечен в производство иммунного посредника интерлейкина-1-бета). Когда показали, что ген ced-3 важен для апоптоза у нематод, ученые присмотрелись к гену ICE и поняли, что он тоже отвечает за производство каспазы. Теперь его называют каспаза-1, хотя он, видимо, играет не столь существенную роль в апоптозе у человека, как ced-3 в апоптозе нематод. Каспазы и другие похожие ферменты (паракаспазы, метакаспазы) были найдены у грибов, растений, водорослей, простейших и даже губок. Они практически универсальны у всех эукариотических организмов, и надо полагать, что их предшественники уже были у самых ранних эукариот, то есть, возможно, 1,5–2 миллиарда лет назад.
Я не собираюсь заводить вас в болото подробностей. Скажу только, что регуляция апоптоза — это сложный, многоступенчатый процесс, когда одна каспаза активирует другую, та — следующую и т. д. Под конец каспазного каскада в действие приводится маленькая армия палачей, которая и разрубает клетку на части[60]. Этот процесс может быть обращен вспять практически на каждой ступени за счет других белков, отвечающих за то, чтобы ложная тревога не привела к оргии смерти и разрушения.
Митохондрии — ангелы смерти
Только что описанные представления сложились к середине 1990-х гг. Никакие из них не были опровергнуты. Тем не менее за последнее время интерпретация накопленных фактов настолько изменилась, что сложившаяся в 1990-е гг. парадигма подверглась революционным изменениям. С точки зрения этой парадигмы ядро клетки представляет собой ее операционный центр, и оно же контролирует ее судьбу. Во многих отношениях, конечно, так и есть, но в случае апоптоза это не так. Живительно, но клетки, лишившись ядра, сохраняют способность к апоптозу. Принципиально новым было открытие того факта, что судьбу клетки контролируют митохондрии. Именно они решают, жить ей или умереть.
Машину смерти можно привести в действие двумя способами. Раньше они казались очень разными, но последние исследования находят у них некоторые общие черты. Первый механизм называется внешним путем апоптоза, потому что сигнал к запуску машины смерти подается извне, через «рецепторы смерти» на внешней стороне клеточной мембраны. Например, активированные иммунные клетки производят химические сигналы (такие, как факторы некроза опухоли), связывающиеся с «рецепторами смерти» раковых клеток, находящихся на начальных стадиях трансформации. «Рецепторы смерти» передают в клетку сигнал, который активирует каспазы и вызывает апоптоз. Было понятно, что многие подробности нуждаются в уточнении, но казалось, что картина в общем и целом ясна. Ничего подобного!
Второй механизм запуска машины смерти называется внутренним путем апоптоза. Как следует из названия, толчок к самоубийству приходит изнутри и обычно связан с повреждением клетки. Например, повреждение ДНК в результате ультрафиолетового облучения активирует внутренний путь, приводя к апоптозу клетки без какого-либо внешнего сигнала. Были найдены сотни триггеров внутреннего пути апоптоза, которые не действуют через «рецепторы смерти», а непосредственно повреждают клетку. Они потрясающе разнообразны. Апоптоз вызывают многие токсины и вещества, загрязняющие окружающую среду, а также некоторые лекарства, использующиеся в химиотерапии рака. Вирусы и бактерии тоже могут вызывать апоптоз. Это особенно хорошо видно в случае СПИДа, когда погибают сами иммунные клетки. Вызывают апоптоз и многие стрессовые факторы: перегрев, переохлаждение, воспаление, оксидативный (или окислительный) стресс. Клетки могут совершать апоптоз после сердечного приступа, инсульта или трансплантации органа. Все эти разнообразные пусковые механизмы независимо приводят к одному и тому же результату — активации каскада каспаз, и поэтому характер апоптоза во всех этих случаях очень похож. Надо полагать, сигналы каким-то образом сходятся к одному и тому же «выключателю», который переводит фермент каспазу из неактивной формы в активную. Эта биохимическая задача специфична как замок и подходящий к нему ключ. Но что, спрашивается, может распознать разные сигналы, оценить их силу и направить их по единому пути одним поворотом ключа — активацией каскада каспаз?
Половину ответа дали в 1995 г. Науфал Замзами и его коллеги, работавшие в составе исследовательской команды Гвидо Кремера в Национальном центре научных исследований (Вильжюиф, Франция). Две публикации этой группы в «Журнале экспериментальной медицины» стали одними из самых цитируемых работ в области медицинских исследований. Некоторые факты тогда уже указывали на то, что митохондрии вовлечены в апоптоз, но группа Кремера доказала, что митохондрии играют в этом процессе ключевую роль. В частности, они показали, что одним из главных триггеров апоптоза является деполяризация внутренней митохондриальной мембраны (см. часть 2 книги). Если мембранный потенциал на некоторое время утрачивается, клетки всегда совершают апоптоз. Во второй статье группа Кремера показала, что этот процесс состоит из двух этапов. За деполяризацией мембраны следует резкое увеличение числа свободных радикалов, которое, по-видимому, нужно для перехода к следующей стадии апоптоза.
Такой двухтактный ход — деполяризация митохондриальной мембраны и выброс свободных радикалов — является реакцией практически на все внутренние триггеры. Иными словами, митохондрии являются и сенсорами, и передатчиками самых разнообразных сигналов при повреждении клетки. Достаточно перенести апоптотические митохондрии в нормальную клетку, и ее ядро фрагментируется, а клетка погибнет. Напротив, заблокировав «двухтактный ход», можно задержать или даже предотвратить апоптоз. Однако оставался вопрос: как апоптотические митохондрии взаимодействуют с клеткой? В частности, как они активируют каспазы?
На этот вопрос ответила группа Сяолун Ванга из Университета Эмори (Атланта, Джорджия) в 1996 г. Как выразился один специалист, ответ вызвал «всеобщее остолбенение». Он гласил: «цитохром с». Если помните, мы встречались с ним в части 2. Этот белок, открытый Дэвидом Кейлином в 1930 г., является компонентом дыхательной цепи и отвечает за перенос электронов от комплекса III к комплексу IV. В норме он связан с наружной стороной внутренней мембраны митохондрий, то есть обращен в межмембранное пространство (см. рис. 5). Группа Ванга обнаружила, что при апоптозе цитохром с высвобождается из митохондрий. Оказавшись в клетке, он связывается с несколькими другими молекулами, образуя апоптосому, которая, в свою очередь, активирует одного из последних палачей — каспазу 3. Выброс цитохрома с из митохондрий означает неизбежную смерть клетки; если ввести его в здоровую клетку, она погибнет. Иными словами, неотъемлемая часть дыхательной цепи, отвечающей за производство энергии, необходимой для жизни клетки, является также и неотъемлемой частью апоптоза, отвечающего за ее смерть. Дилемма «жизнь или смерть» зависит от локализации одной конкретной молекулы. Ничто в биологии не сравнится с этим двуликим Янусом: с одной стороны — жизнь, с другой — смерть, а между ними несколько миллионных долей миллиметра.
Цитохром с — не единственный белок, высвобождающийся из митохондрий. Высвобождаются и некоторые другие белки, в том числе и такие, которые вовлечены в апоптоз, и иногда даже более сильно, чем цитохром с. Некоторые из этих белков активируют каспазы, другие (например, апоптоз-индуцирующий фактор, AIF) атакуют другие молекулы (например, ДНК) без помощи каспаз. Как часто бывает в биохимии, подробности кажутся неимоверно сложными, но основополагающие принципы просты: деполяризация внутренней мембраны митохондрий и образование свободных радикалов приводят к выходу в цитозоль цитохрома с и других белков; они активируют определенные ферменты; эти ферменты уничтожают клетку.
Битва жизни и смерти
Когда выяснилось, что жизнь или смерть клетки зависит от локализации цитохрома с и других судьбоносных белков, медицинские исследования, конечно же, обратились к поиску механизма, который приводит к высвобождению этих молекул из митохондрий. Этот механизм тоже непростой, но помогает понять связь между внутренним и внешним путем апоптоза. За исключением ряда случаев (которые, скорее всего, непринципиальны), главную роль в обеих формах клеточной смерти играют митохондрии. Почти всегда именно они держат руку на выключателе машины смерти. Когда достаточное число митохондрий изливает в клетку свои смертоносные белки, клетка совершает самоубийство.
Согласно последним исследованиям Стена Оррениуса и его коллег из Каролинского института (Стокгольм, Швеция), высвобождение цитохрома с происходит в два этапа. На первом этапе белок мобилизуется из митохондриальной мембраны. Цитохром с непрочно связан с липидами мембраны (особенно с кардиолипином) и высвобождается из внутренней митохондриальной только при их окислении. Это объясняет, зачем при апоптозе нужны свободные радикалы: они окисляют липиды внутренней мембраны, что приводит к высвобождению цитохрома с. Но это еще не все. Далее цитохром с переходит в межмембранное пространство, однако не может покинуть митохондрии до тех пор, пока внешняя мембрана не станет более проницаемой. Дело в том, что цитохром с — это белок, а белковые молекулы слишком велики, чтобы просто проникнуть через мембрану. Чтобы он мог покинуть митохондрию, должна открыться какая-то мембранная пора.
Более десяти лет ученые никак не могли понять, что это за пора. Казалось, в разных ситуациях работают разные механизмы и по меньшей мере существуют два типа пор. Один механизм явно предполагает метаболический стресс самих митохондрий, приводящий к избыточному производству свободных радикалов. С повышением уровня стресса во внешней мембране открывается пора переходной проницаемости. Это приводит к набуханию и разрыву мембраны, а также высвобождению белков.
Еще одна пора, которая, скорее всего, более универсальна, связана с большим семейством белков bcl-2. Это название, по большому счету, устарело (оно связано с открытым в 1980-х гг. онкогеном и означает B-клеточная лимфома/лейкемия-2). Сейчас нам известен по крайней мере 21 ген, кодирующий белки семейства bcl-2. Их можно разделить на две большие группы, которые ведут друг с другом сложную и пока малопонятную войну. Одна группа белков защищает от апоптоза. Они находятся на внешней митохондриальной мембране и, видимо, предотвращают образование пор, не давая цитохрому с и другим белкам выходить в цитозоль. Другая группа действует противоположным образом. Они образуют поры, через которые, видимо, цитохром с и другие белки могут покинуть митохондрии. Таким образом, эта группа белков способствует апоптозу. В норме они есть во всей клетке и перемещаются в митохондрии, только получив определенный сигнал. Конечный итог, то есть то, совершает клетка апоптоз или нет, зависит от численного соотношения враждующих членов семейства в митохондриальной мембране, а также от числа митохондрий, принимающих участие в битве. Если сторонники апоптоза численно превосходят его противников, поры открываются, из митохондрий изливаются смертоносные белки и клетка совершает самоубийство.
Существование враждующего семейства белков bcl-2 помогает понять связь между внутренним и внешним путем апоптоза. На исход междоусобной войны влияет множество разных сигналов. Например, и «сигналы смерти» снаружи (внешний путь), и «сигналы повреждения» изнутри (внутренний путь) склоняют чашу весов в пользу апоптоза[61]. Таким образом, белки bcl-2 интегрируют разнообразные сигналы как снаружи, так и изнутри и оценивают их силу. Если смерть перевешивает, во внешней мембране митохондрий образуются поры, цитохром с и другие белки выходят в цитозоль и активируется каспазный каскад. Поэтому последние этапы апоптоза, как правило, одинаковы.
Центральная роль митохондрий в обоих путях апоптоза наводит на мысль, что так было всегда. Мы уже обсуждали, что бактерии и раковые клетки действуют независимо, в своих собственных интересах, и поэтому их можно считать «единицами отбора». Отбор может одновременно действовать на уровне клетки и на уровне особи. Митохондрии некогда были свободноживущими бактериями и привыкли действовать независимо. Став частью другого организма, они, надо полагать, сохранили способность к независимым действиям, по крайней мере на некоторое время, и могли взбунтоваться так, как это делают раковые клетки.
Если сегодня митохондрии убивают клетку, в которой находятся, то, может быть, они делали это и в самом начале симбиоза в своих собственных интересах? Может быть, апоптоз возник не для блага особи, а из-за эгоизма постояльца? Если так, то это скорее убийство, чем самоубийство. Зато понятно, как клетки «согласились» принять смерть — эта была диверсия изнутри. Так есть ли какие-то доказательства того, что митохондрии протащили в эукариотическую клетку машину смерти? Да, такие доказательства есть.
Войны паразитов?
Ген, кодирующий цитохром с, был принесен в эукариотическую клетку предками митохондрий, а впоследствии переместился в ядро (см. часть 3 книги). Мы знаем это потому, что практически идентичная генная последовательность есть у α-протеобактерий и она является частью дыхательной цепи, самым важным вкладом в партнерство. Менее ясно, насколько важен был цитохром с на ранних этапах эволюции апоптоза. Хотя он и играет определяющую роль в апоптозе у млекопитающих и, возможно, у растений, он не нужен для апоптоза у плодовых мушек или нематод. Ясно, что он не является универсальным игроком. Играл он главную роль в инициации апоптоза на ранних этапах эволюции и был потом отстранен от этой роли у нескольких видов, или приобрел ключевое значение относительно недавно, независимо у растений и млекопитающих? Это мы узнаем только тогда, когда будем больше знать об апоптозе у самых примитивных эукариот. Как мы видели, цитохром с — только один из многих белков, которые высвобождаются из митохондрий во время апоптоза. Эти белки имеют очень странные названия: Smac/DIABLO, Omi/HtrA2, endonuclease G, AIF (у плодовой мушки названия таких белков гораздо лучше отражают их функции — Reaper, Grim, Sickle). Некоторые из них иногда играют даже более важную роль, чем цитохром с. Большинство этих белков было открыто в нынешнем тысячелетии, но благодаря многочисленным проектам секвенирования геномов мы уже знаем кое-что об их происхождении. Оно поразительно. За единственным исключением (AIF, апоптоз — индуцирующий фактор), все известные апоптотические белки, высвобождающиеся из митохондрий, имеют бактериальное происхождение. У архей их нет. (Вспомним, о чем мы говорили в части 1: клетка-хозяин, скорее всего, была археем, а митохондрии — бактериями.) Это означает, что у клетки-хозяина практически не было машины смерти. Митохондрии принесли в эукариотический союз не все апоптотические белки, некоторые попали в эукариотические клетки позже, в результате горизонтального переноса генов от других бактерий. Однако единственный вклад архей в машину смерти эукариотической клетки — это AIF, и надо заметить, что у современных архей он не имеет никакого отношения к смерти клеток.
Бактериальное происхождение имеют не только белки митохондрий, но и каспазы, если верить данным, полученным при секвенировании их генов. Впрочем, бактериальные каспазы довольно смирные: они режут белки на кусочки, но не вызывают смерть клеток. Более загадочно происхождение семейства белков bcl-2. Последовательности их генов имеют мало общего как с бактериями, так и с археями. Тем не менее трехмерная структура этих белков напоминает бактериальные белки, в частности, группу токсинов некоторых патогенных бактерий, таких как дифтерийная палочка. Как и вызывающие апоптоз белки семейства bcl-2, бактериальные токсины образуют поры в мембране клетки-хозяина, а иногда даже вызывают апоптоз.
Все это наводит на мысль о том, что большая часть машины смерти была привнесена в эукариотический симбиоз предками митохондрий. Тогда это похоже не на самоубийство, а на коварное убийство, акт вопиющей неблагодарности со стороны постояльца. Эта идея легла в основу убедительной гипотезы, предложенной Хосе Фраде и Теологосом Михелидисом из Института физиологии им. Макса Планка (Мартинсрид, Германия) еще в 1997 г. Данные, накопившиеся с тех пор, по большей части подтверждают ее.
Фраде и Михелидис провели параллель между поведением современной бактерии Neisseria gonorrhoeae (возбудителя гонореи, заболевания, передающегося половым путем) и тем, как могли бы вести себя протомитохондрии. N. gonorrhoeae заражает клетки уретры и шейки матки, а также лейкоциты. Оказавшись внутри, эти бактерии проявляют дьявольскую хитрость. Они продуцируют образующий поры белок PorB (он похож на митохондриальные белки bd-2) и «вставляют» его в клеточную мембрану хозяина, а также в мембрану вакуоли, в которую «завернуты» бактерии внутри клетки. Эти поры плотно закрыты, пока взаимодействуют с АТФ клетки (опять-таки сходным образом ведут себя некоторые белки bcl), но когда запасы АТФ хозяина истощаются, поры открываются. Открытие пор запускает машину апоптоза, и клетка погибает. Сами бактерии тем не менее выживают. Они спасаются бегством, прихватив с собой в качестве провианта аккуратно упакованные погибшие клетки. Таким образом, бактерии живут в клетке, пока та здорова, контролируя ее способность поддерживать запасы АТФ, но как только запасы начинают истощаться, бактерии убивают ее и отправляются на поиски новых клеток. Вот мерзавцы!
Фраде и Михелидис отмечают, что N. gonorrhoea — не единственная бактерия, способная на такое коварство. К сходной тактике прибегает страшный бактериальный хищник Bdellovibrio, с которым мы уже встречались в части 1. Проникнув внутрь другой бактерии, он тоже некоторое время контролирует ее метаболическое здоровье, а потом поедает изнутри. Кстати, Линн Маргулис называла Bdellovibrio одним из возможных предков митохондрий. Еще один претендент — бактерия Rickettsia prowazekii, о которой мы говорили в частях 1 и 3, — тоже внутриклеточный паразит. Такая «биохимическая археология» — свидетельство того, что сначала отношения между митохондриями и содержащими их клетками были паразитическими. Попав внутрь архея, протомитохондрии некоторое время следили за его здоровьем, потом вызывали его смерть, подъедали его останки и шли искать следующего хозяина.
Происхождение апоптоза в вооруженной борьбе будущих партнеров по симбиозу означает, что эукариотический симбиоз начинался с того, что паразит убивал хозяина и шел искать нового. Это, как вы уже догадались, буквально то, о чем говорила Линн Маргулис и другие биологи. В наследство от этих отношений эукариотической клетке досталась машина смерти, которая потом нашла другое применение и стала использоваться для программируемого самоубийства клеток у многоклеточных организмов. Но вовсе не о войнах паразитов говорили мы в части 1, когда обсуждали происхождение эукариотической клетки; нет, там речь шла о сотрудничестве двух миролюбивых прокариот, живших бок о бок в метаболическом браке. В тот момент мы рассмотрели и отбросили допущение о том, что отношения между двумя клетками были паразитическими. Теперь, посмотрев на вещи под другим углом, мы вернулись к этому допущению. В этой области науки ни в чем нельзя быть уверенным: приходится постоянно взвешивать все доводы, имеющие хоть какое-то отношение к делу. Только что обсуждавшийся довод, несомненно, весом. Означает ли это крушение и без того утлой лодчонки наших построений? Неужели мне — о, ужас! — придется переписывать первую часть?
Основы индивидуума
Многоклеточная особь состоит из клеток, сотрудничающих ради высшего блага. Тем не менее они сотрудничают не по любви, а под угрозой смертной казни для любой клетки, которая попробует дезертировать к предковому образу жизни. Время от времени эгоистичным клеткам удается избежать казни, тогда наступает рак. Раковые клетки бесконтрольно размножаются, забыв об общих интересах и подрывая единство тела. Отсрочив свою смерть, они в конце концов убивают бывшего хозяина и сами гибнут вместе с ним.
Рак существует потому, что редко встречается у молодых особей. Если бы тело гибло прежде, чем его клетки успевали бы организовать свое размножение (за счет половых клеток), то особь не оставила бы потомства и эгоистичные гены канули бы в Лету. На заре существования многоклеточных организмов, однако, составляющие его эгоистичные клетки имели шансы на независимое существование. В отличие от раковых клеток, они могли жить сами по себе, сохраняя потенциальную возможность основать новую колонию клеток. Эта независимость до сих пор встречается у губок и некоторых других животных, но такая либеральная политика в отношении клеток не дает им подняться к вершинам многоклеточной сложности. Истинная многоклеточность требует абсолютной жертвенности. Но если клетки могли жить независимо, как они согласились подписать собственный смертный приговор?
Сегодня смертная казнь клеток — апоптоз — отправляется митохондриями. Они объединяют сигналы из разных источников, и если их баланс указывает, что клетка повреждена, и потому может начать действовать в собственных интересах, митохондрии активируют клеточную машину смерти. Примерно 10 миллиардов клеток человеческого тела ежедневно умирают тихой и незаметной смертью путем апоптоза. На смену им приходят новые, неповрежденные клетки. Машина смерти состоит из нескольких белков, которые выходят из митохондрий в цитоплазму и активируют дремлющие «ферменты смерти» — каспазы. Эти ферменты расчленяют клетку изнутри и пакуют остатки, чтобы их пустили в дело другие клетки. Ничто не пропадает впустую.
Практически все «белки смерти», высвобождающиеся из митохондрий, а также сами каспазы, были некогда привнесены в эукариотическую клетку бактериальными предками митохондрий. У них по сей день есть ближайшие аналоги среди белков свободноживущих бактерий и особенно среди паразитических. У современных бактерий многие «белки смерти» вовсе не вызывают смерть самих бактерий или каких-либо других организмов, а используются в иных целях. С другой стороны, бактериальные белки семейства поринов — это боевое оружие, активно действующее на другие клетки. Возможно, так было всегда: бактериальные предки митохондрии были паразитами и использовали похожие на порины белки, чтобы расчленить клетку хозяина изнутри и съесть ее, после чего отправлялись искать новую жертву.
Так это было или нет, зависит от истинной природы бактериальных поринов. У современных паразитов они встроены в мембраны клетки-хозяина и безжалостно убивают его, как только он оказывается не в состоянии обеспечивать метаболические запросы паразита. Физически (но не генетически). бактериальные порины подозрительно похожи на митохондриальные порины — белки bcl-2, которые активируют клеточную машину смерти, образуя поры в митохондриальной мембране. Из этого следует и более широкий вывод: эукариотическая клетка родилась в горниле войны между внутриклеточным паразитом, который впоследствии был усмирен и стал митохондрией, и клеткой-хозяином, которая научилась справляться с инфекцией.
Звучит довольно просто, но есть одна загвоздка. В части 1 мы рассмотрели несколько теорий происхождения эукариотической клетки, в частности, «паразитическую модель», в рамках которой митохондрии произошли от похожей на рикеттсию бактерии, и водородную гипотезу, которая утверждает, что изначально союз был основан на взаимной метаболической выгоде: один партнер использовал продукт метаболизма другого, и наоборот. Там я утверждал, что современные данные поддерживают водородную гипотезу, а не паразитическую модель. Однако только что сказанное о паразитах не согласуется с водородной гипотезой, предполагающей мирный метаболический союз. Паразиту может быть выгодно убить хозяина, чтобы найти следующего, а вот метаболическому наркоману незачем убивать поставщика, особенно если мало надежды найти другого. Значит, либо паразитическая модель подрывает доверие к водородной гипотезе, либо она сама неверна, что бы там она ни объясняла. Не могут обе эти теории быть правильными одновременно. Так какая же из них ближе к истине?
Чтобы ответить на этот вопрос, нам сначала нужно отделить проверенные или, по крайней мере, пока не оспоренные факты от хитроумных домыслов. Это нетрудно. Понятно, что митохондрии в большой мере отвечают за машину смерти: они играют ключевую роль в апоптозе современных организмов и наверное внесли важный вклад в его эволюцию. А вот связь между белками bcl-2 и бактериальными поринами, например у Neisseria gonorrhoeae, относится к хитроумным домыслам. Загадочное структурное сходство действительно есть, но оно не доказывает эволюционную связь.
На основании современных данных можно предположить три возможных варианта взаимоотношений между белками bcl-2 и бактериальными поринами. Во-первых, сходство между ними может быть связано с конвергентной эволюцией: и митохондрии и N. gonorrhoeae могли независимо приобрести похожие белки для сходных целей. Никакие генетические данные не исключают эту возможность, а тем, кто сомневается в силе конвергентной эволюции на молекулярном уровне, я рекомендую книгу Саймона Конвей-Морриса «Разгадка жизни». В этом случае между белками bcl-2 и бактериальными поринами не должно быть генетического родства, но можно ожидать структурного сходства, связанного с общностью функций. Есть не так уж много способов проделать большую пору в липидной мембране, и они должны накладывать определенные функциональные ограничения. Если двум разным клеткам нужны большие поры, они, скорее всего, придут к сходным решениям.
Вторая возможность заключается в том, что митохондрии действительно унаследовали белки bcl-2 от своих бактериальных предков, как предложили Фраде и Мехелидис (см. предыдущую главу). Доказать это можно, только найдя общие черты в генетических последовательностях, а они не найдены. Более того, такие общие черты должны быть у представителей α-протеобактерий (предков митохондрий), или придется допустить возможность горизонтального переноса генов на более поздней стадии. Ясно, что если горизонтальный перенос генов происходил позже, он ничего не скажет нам об изначальных отношениях между митохондриями и клеткой-хозяином. Итак, более систематическое изучение генов α-протеобактерий может подкрепить эту гипотезу, но пока что структурное сходство — в лучшем случае повод задуматься.
Наконец, возможно, что N. gonorrhoea и другие паразитические бактерии получили свои порины от митохондрий, а не наоборот. Такой перенос генов от хозяина к паразиту — обычное дело. Если это было так, то следует ожидать сходства последовательностей у генов митохондрий и паразитов. Отсутствие таких сходных последовательностей может быть связано с тем, что их просто не искали (и тогда они всплывут, когда мы отсеквенируем больше генов), а может быть, они просто потерялись, стерев все свидетельства общности происхождения. Такое тоже возможно, так как непрекращающаяся эволюционная война между паразитом и хозяином приводит к исключительному непостоянству генов паразита. Более того, бактериальные порины сами по себе не вызывают весь апоптоз, они просто подключаются к уже существующей машине смерти. По сути дела, они несут с собой портативный тумблер, позволяющий «включить» машину смерти клетки хозяина. Поэтому поведение современных паразитов, вызывающих клеточную смерть, несравнимо с предполагаемым поведением протомитохондрий. Последним пришлось бы притащить в клетку всю машину смерти, наладить ее работу, но при этом не погибнуть самим. (Сегодня, конечно, митохондрии погибают вместе с клеткой.)
Имеющиеся данные не позволяют сделать выбор между этими тремя возможностями. Тем не менее батальное полотно кисти Фраде и Михелидиса, по крайней мере, выглядит непротиворечиво. Или все-таки нет? В этой истории есть целый ряд других заковыристых проблем. Во-первых, и это главное, митохондрии больше не являются независимо размножающимися клетками, и скорее всего они потеряли независимость сразу после начала переноса их генов в ядро клетки-хозяина. Как только несколько важных генов оказались заложниками в ядре, убийство хозяина не сулило митохондриям ничего хорошего. Их будущее было неразрывно связано с будущим хозяина. Можно было попробовать манипулировать хозяином, но убивать его явно не стоило. Напротив, никакие паразиты, даже мельчайшие Rickettsia, не потеряли свою независимость. Они полностью контролируют свой жизненный цикл и свои ресурсы. Им, в отличие от митохондрий, убийство хозяина может сойти с рук.
Когда именно митохондрии потеряли власть над собственным будущим, неизвестно, но скорее всего это произошло на самых ранних этапах эволюции эукариотической клетки. Посмотрим, например, на эволюцию переносчика АТФ — мембранной помпы, экспортирующей АТФ из митохондрий. Этот переносчик впервые позволил эукариотическим клеткам получать энергию в форме АТФ из митохондрий (до этого момента само слово «митохондрии» было к ним неприменимо). Это был символический шаг, так как симбионты потеряли контроль над своими энергетическими ресурсами, а с ним и независимость. Для митохондрий это означало переход от статуса гостя к статусу пленника. Мы можем довольно точно датировать этот переход, сравнивая последовательности гена, кодирующего переносчика АТФ, у разных групп эукариот. Тот факт, что этот переносчик встречается во всех группах эукариот, включая растения, животных, грибы, водоросли и простейших, наводит на мысль, что он возник до расхождения этих групп, то есть очень давно и, само собой, до появления многоклеточных животных (судя по ископаемой летописи, возможно, за несколько сотен миллионов лет).
Итак, налицо временной провал. Кажется весьма вероятным, что митохондрии потеряли автономию задолго до возникновения настоящих многоклеточных организмов. В тот период убийство хозяина не сулило митохондриям никаких преимуществ, потому что они уже не могли существовать независимо. Клеткам-хозяевам, в свою очередь, была абсолютно невыгодна смерть, потому что тогда они еще не являлись частью многоклеточного организма. Таким образом, нынешние преимущества апоптоза — жесткой полицейской системы многоклеточного организма — не имели силы.
Это парадокс. Судя по всему, специализированная машина смерти была никому не нужна. Казалось бы, естественный отбор должен был избавиться от нее, но этого не произошло. Мы также знаем, что происхождение существенной части ее механизма связано с митохондриями. В довершение всего, водородная гипотеза, которую я так превозносил, утверждает, что эукариотическая клетка родилась от метаболического союза двух мирно сосуществующих клеток, которым не было выгодно убивать друг друга. Кажется, я завел вас в тупик. Одна из клеток протащила в мирный союз налаженную машину смерти, вредную для обоих членов союза; несмотря ни на что, эта машина сохранялась на протяжении нескольких сотен миллионов лет, пока наконец ей не нашлось применение. Можно ли найти логичное обоснование этому безумному сценарию? Да, можно, но только если мы готовы на уступку. Машина смерти не всегда вызывала смерть. В один прекрасный день она вызвала появление пола.
Секс и смерть
Давайте рассмотрим первых эукариот с точки зрения водородной гипотезы, предполагающей мирное сосуществование. Во введении к части 5 книги мы обсуждали, что естественный отбор может действовать на разных уровнях — на уровне особи, ее клеток, митохондрий и, конечно, генов. Мы видели, что когда речь идет о клетках, которые, как бактерии, размножаются бесполым путем, не всегда правильно считать, что естественный отбор действует на уровне генов. В таких случаях отбор в основном работает на уровне отдельных клеток, которые и есть истинные самовоспроизводящиеся единицы. Эти соображения сейчас нам очень пригодятся, потому что мы должны по отдельности рассмотреть интересы митохондрий и интересы содержащих их клеток на заре существования эукариотического симбиоза. Тогда и тех и других можно было считать отдельными клетками (в следующих главах мы увидим, что такой подход может оказаться полезным, даже когда речь идет о современности).
Так какими же были частные интересы протомитохондрий и клеток-хозяев? И как, учитывая сочетание автономии и неустойчивой взаимозависимости, они вообще могли действовать в своих интересах? Убедительный ответ на этот вопрос предложили в 1999 г. двое ученых: Нейл Блэкстоун, один из самых плодотворных мыслителей в области эволюционной биохимии (Университет Северного Иллинойса, США), и Дуглас Грин, один из первых исследователей высвобождения цитохрома с при апоптозе (Калифорнийский университет в Сан-Диего, США).
Митохондрии, как и все клетки, должны пролиферировать, то есть размножаться с увеличением числа клеток. После того как они связали свою судьбу с судьбой хозяина, убивать его и находить следующего не было смысла, так как вне клетки они бы не выжили. Кроме того, есть предел размножения митохондрий в одной клетке: митохондриальный «рак» погубил бы и клетку и митохондрии. Таким образом, успешное размножение митохондрий возможно только параллельно с размножением клетки-хозяина. Когда она делится, популяция митохондрий должна удвоиться, чтобы обеспечить митохондриями дочернюю клетку. Конечно, клетка-хозяин тоже спит и видит, как бы ей поделиться, поэтому ее интересы и интересы митохондрий совпадают. В противном случае этот союз вряд ли продолжался бы два миллиарда лет. Он бы распался в самом начале, и мы бы сейчас не ломали голову над этой проблемой, потому что нас не было бы.
Но интересы митохондрий и клетки-хозяина не всегда совпадают. Что произойдет, если клетка-хозяин по какой-то причине откажется делиться? Ясно, что ее митохондрии тоже не смогут размножаться (ну, то есть они могли бы, но только до определенного предела, а потом разрушили бы клетку и себя самих вместе с ней). Исход зависит от того, почему клетка-хозяин отказалась делиться. Скорее всего, причиной было отсутствие пищи. В части 3 мы говорили о том, что, несмотря на огромный потенциал в плане размножения, бактерии проводят большую часть жизни в заторможенном состоянии. То же самое, скорее всего, относилось и к ранним эукариотам. Если так, им оставалось только подтянуть кушаки и ждать обеда, после которого можно снова начинать размножаться. Интересы митохондрий и клетки-хозяина снова совпадали: если митохондрии будут вынуждать хозяина делиться в условиях нехватки ресурсов, они погибнут вместе с ним. Куда лучше направить оставшиеся ресурсы на повышение устойчивости к вероятному в период лишений физическому стрессу — жаре, холоду, ультрафиолету. В таких случаях многие клетки образуют устойчивую спору, переживая неблагоприятные условия в состоянии покоя и возвращаясь к жизни, когда все налаживается.
Кроме того, клетке могло помешать делиться повреждение, в том числе повреждение ядерной ДНК. Тут интересы хозяина и митохондрии начинают расходиться. Предположим, что пищи много, но клетка-хозяин тем не менее не делится. Я прямо вижу, как митохондрии кричат, прижимаясь лицом к решетке: «Выпустите нас! Мы не сделали ничего плохого!» Тем временем соседние клетки, посмеиваясь, продолжают делиться, и их митохондрии радостно делятся вместе с ними. Что же делать митохондриям, которые оказались в ловушке? Убивать хозяина невыгодно, тогда они погибнут сами. Хорошо бы заставить клетку-хозяина слиться с другой клеткой и рекомбинировать ДНК с ДНК партнера. Рекомбинация ДНК обычна у бактерий и является основой пола у эукариот. Слившаяся клетка получает шанс на новую жизнь, а митохондрии — место, где можно размножаться.
Учитывая, что половой процесс — крайне затратная вещь, ожесточенные споры о том, почему он возник, бушуют до сих пор. Скорее всего, вклад внесли несколько факторов. Уильям Хамильтон предполагал, что половой процесс «маскирует» повреждения ДНК (поврежденный ген, вероятно, окажется в паре с неповрежденной копией того же гена), а генерируемая рекомбинацией изменчивость может давать клетке преимущества в борьбе с паразитами. Последние данные говорят о том, что ни одной из этих причин по отдельности недостаточно, чтобы объяснить возникновение полового процесса. Однако они не противоречат друг другу, а преимущества полового процесса, по всей видимости, разнообразны. С другой стороны, его происхождение окутано тайной. У бактерий есть рекомбинация, но нет слияния клеток. Напротив, у большинства эукариот при половом размножении происходит слияние двух клеток, затем слияние их ядер и, наконец, рекомбинация их генов (самое «обязывающее» из всех этих действий). Почему эукариотические клетки вообще стали сливаться? Понятно, что утрата бактериями неудобной клеточной стенки сделала сам физический акт слияния в принципе возможным, но потребности сливаться это не объясняет. Может быть, это митохондрии заставили эукариотические клетки сливаться друг с другом? Может быть, половое слияние — это тоже митохондриальная диверсии? Том Кавалье-Смит, с которым мы встречались в части 1 книги, говорил, что у первых эукариот слияние клеток, скорее всего, было обычным делом. Он утверждает, что деление клетки при половом процессе (мейозе), когда число хромосом сначала удваивается, а потом снова делится пополам, произошло за счет нескольких простых шагов как способ восстановления исходного числа генов и ядер после слияний клеток. Тогда слияние должно было бы произойти в любом случае, а митохондрии могли просто агитировать за него.
Могут ли митохондрии манипулировать клеткой, в которой находятся? Это серьезный вопрос. Мы знаем, что современные митохондрии могут, ведь они вызывают апоптоз. Но могли они манипулировать эукариотической клеткой на заре ее существования? Нейл Блэкстоун предположил, что они делали это очень хитрым способом, который объясняет и неудержимую тягу к слиянию, и происхождение апоптоза.
Дата добавления: 2019-02-13; просмотров: 327; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!