Особенность белкового обмена в эритроцитах
В зрелом эритроците белки не синтезируются, т.к. у него нет рибосом, ЭПР, аппарата Гольджи и ядра. Однако в цитоплазме синтезируется пептид глутатион.
Биосинтез глутатиона осуществляется в 2 стадии:
1). АТФ + глутаминовая кислота + цистеин ® γ-глутамилцистеин + АДФ + Фн
2). АТФ + γ-глутамилцистеин + глицин ® глутатион + АДФ + Фн
Первая стадия катализируется γ-глутамилцистеинсинтетазой, вторая стадия – глутатионсинтетазой.
Катаболизм белков в эритроците неферментативный. Белки разрушаются и инактивируются в эритроците под действием неблагоприятных факторов: СРО, гликозилирования, взаимодействия с тяжелыми металлами и токсинами.
Особенность обмена нуклеотидов в эритроцитах
В зрелом эритроците:
1. из ФРПФ (из рибозо-5ф) и аденина может синтезироваться АМФ.
2. АМФ с участием АТФ превращается в АДФ.
3. В реакциях субстратного фосфорилирования (гликолиз) АДФ превращается в АТФ.
4. В гликолизе НАД+ восстанавливается в НАДН2, который используется для регенерации гемоглобина из метгемоглобина.
5. В ПФШ НАДФ+ восстанавливается в НАДФН2, который используется для функционирования антиоксидантной системы.
Особенность липидного обмена в эритроцитах
В зрелом эритроците липиды не синтезируются, однако эритроцит может обмениваться липидами с липопротеинами крови. Катаболизм липидов неферментативный, повреждение и разрушение липидов происходит в реакция ПОЛ.
|
|
|
Особенность углеводного обмена в эритроцитах
В зрелых эритроцитах углеводы не синтезируются. Катаболизм углеводов происходит на 90% в анаэробном гликолизе и на 10% в ПФШ, основной субстрат – глюкоза. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Наряду с глюкозой эритроцит может использовать фруктозу, маннозу, галактозу, а также инозин, ксилит и сорбит.
В процессе гликолиза с участием фосфоглицераткиназы и пируваткиназы образуется АТФ, а с участием 3-ФГА дегидрогеназы восстанавливается НАДН2. В окислительной стадии ПФШ с участием глюкозо-6-фосфат дегидрогеназы и 6-фосфоглюконат дегидрогеназы восстанавливается НАДФН2.
Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и направляется преимущественно в печень для глюконеогенеза.
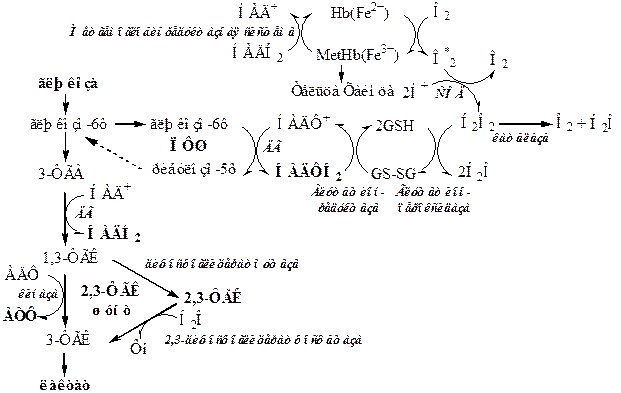
Энергетический обмен в эритроцитах
Образующаяся в анаэробном гликолизе АТФ используется для функционирования транспортных АТФаз, для работы цитоскелета и синтеза некоторых веществ. За 1 час все эритроциты крови потребляют 0,7г глюкозы.
Генетический дефект любого фермента гликолиза приводит к уменьшению образования АТФ, в результате падает активность Na+,К+-АТФ-азы, повышается осмотическое давление и возникает осмотический шок.
|
|
|
Для оценки эффективности работы транспортных систем определяют осмотическую резистентность эритроцитов. Осмотическая резистентность эритроцитов в свежей крови в норме составляет 0,20-0,40% NaCl.
Обезвреживание активных форм кислорода в эритроцитах
Высокое содержание О2 в эритроцитах является причиной образования большого количества активных форм кислорода. Постоянным источником активных форм кислорода в эритроцитах является неферментативное окисление гемоглобина в метгемоглобин: 1). Hb (Fe2+) ® Met Hb (Fe3+) +e- 2). e- + O2 → О∙2
Также СРО в эритроците стимулируют различные окислители - нитраты, сульфаниламиды, противомалярийное лекарство примахин.
Образующиеся активные формы кислорода запускают реакции СРО, которые приводят к разрушению липидов, белков, углеводов и др. органических молекул и являются причиной старения и гемолиза эритроцита.
Для сдерживания СРО в эритроците функционирует ферментативная антиоксидантная система. Для ее работы необходим глутатион и НАДФН2.
Супероксиддисмутаза (Cu2+ и Zn2+) превращает супероксидные анионы в перекись водорода: 2О∙2 + 2H+ → H2O2+ O2
Каталаза - геминовый фермент, разрушает перекись водорода до воды и кислорода: 2Н2О2 → H2O+ O2
|
|
|
Глутатионпероксидаза (селен) при окислении глутатиона разрушает перекись водорода и гидроперекиси липидов до воды:
Н2О2 + 2 GSH → 2 Н2О + G-S-S-G.
Глутатионредуктаза восстанавливает окисленный глутатион с участием НАДФН2:
GS-SG + НАДФН2 → 2 GSH + НАДФ+.
Недостаток глутатиона и НАДФН2 в эритроцитах приводит к снижению АОА, активации ПОЛ и может стать причиной гемолитической анемии. Различные окислители - нитраты, сульфаниламиды, противомалярийное лекарство примахин, усиливают гемолиз эритроцитов.
Дефицитглутатиона может быть обусловлен действием токсических веществ, например ионами тяжелых металлов или наследственным недостатком глутатионредуктазы.
Дефицит НАДФН2 возникает при наследственной недостаточности (аутосомно-рецессивный тип) первого фермента ПФШ глюкозо–6–фосфатдегидрогеназы. Не менее 100 млн человек являются носителями около 3000 генетических дефектов глюкозо-6-фосфатдегидрогеназы.
Для оценки эффективности работы антиоксидантных систем определяют перекисную резистентность эритроцитов.
Обмен метгемоглобина
В течение суток до 3% гемоглобина может спонтанно окисляться в метгемоглобин:
|
|
|
Hb (Fe2+) ® Met Hb (Fe3+) +e-
Восстановление метгемоглобина до гемоглобина осуществляет метгемоглобинредуктазная система. Она состоит из цитохрома b5 и цитохром b5 редуктазы (флавопротеин), донором водорода служит НАДН2, образующийся в гликолизе.
1). Цитохром b5 восстанавливает Fe3+ метгемоглобина в Fe2+ гемоглобина:
MetHb(Fe3+) + цитb5 восст → Hb(Fe2+) + цит b5 окисл
2). Окисленный Цитохром b5 восстанавливается цитохром b5 редуктазой:
цит b5 окисл + НАДН2 → цитb5 восст + НАД+
Восстановление метгемоглобина может осуществляться также неферментативным путём, например, за счёт витамина В12, аскорбиновой кислоты или глутатиона.
У здорового человека концентрация метгемоглобина в крови не превышает 1%.
Генетический дефект ферментов гликолиза и метгемоглобинредуктазной системы приводит к накоплению метгемоглобина и увеличению образования активных форм кислорода. Активные формы кислорода вызывают образование дисульфидных мостиков между протомерами метгемоглобина, что приводит к их агрегации с образованием телец Хайнца. Последние способствуют разрушению эритроцитов при попадании их в мелкие капилляры. Накопление метгемоглобина в крови из-за нарушения транспорта кислорода ведет к гипоксии.
Дифосфоглицератный шунт
Кроме традиционного ПФШ, у гликолиза эритроцитов многих млекопитающих есть свой специфический шунт - 2,3–дифосфоглицератный.
В эритроцитах имеется дифосфоглицератмутаза, которая позволяет обходить в гликолизе фосфоглицераткиназную реакцию. Дифосфоглицератмутаза катализирует превращение 1,3–ФГК в 2,3–ФГК. Ее стимулирует дефицит кислорода. В условиях гипоксии до 20% глюкозы идет по этому пути. Образующаяся 2,3–ФГК встраивается в молекулу гемоглобина и аллостерически уменьшает его сродство к кислороду. Кривая диссоциации оксигемоглобина смещается вправо, что способствует переходу кислорода из оксигемоглобина в ткани.
Под действием 2,3–дифосфоглицератфосфатазы (принято считать, что этой активностью обладает фосфоглицератмутаза) 2,3–ФГК превращается в 3–ФГК, которая возвращается в реакции гликолиза.
При 2,3–дифосфоглицератном шунте в гликолизе не синтезируется АТФ, а свободная энергия 1,3–ФГК, рассеивается в форме теплоты. В этом может заключаться определённое преимущество, поскольку даже в тех случаях, когда потребности в АТФ минимальны, гликолиз может продолжаться.
Образование эритроцитов
Эритроциты, так же как и другие клетки крови, образуются из полипотентных стволовых клеток костного мозга. Стволовая клетка превращается в эритроцит за две недели.
Размножение и превращение начальной клетки эритроидного ряда в унипотентную стимулирует ростовой фактор интерлейкин-3 (цитокин), который синтезируется Т-лимфоцитами и клетками костного мозга.
Дальнейшую пролиферацию и дифференцировку унипотентной клетки эритроидного ряда регулирует гормон эритропоэтин, который синтезируется в почках. Образование эритропоэтина в почках стимулирует недостаток кислорода. Хроническая почечная недостаточность подавляет образование эритропоэтина, что ведет к развитию анемии.
На стадии эритробласта происходят интенсивный синтез гемоглобина, конденсация хроматина, уменьшение размера ядра и его удаление. Образующийся ретикулоцит ещё содержит глобиновую мРНК и активно синтезирует гемоглобин. Циркулирующие в крови ретикулоциты лишаются рибосом, ЭР, митохондрий и в течение двух суток превращаются в эритроциты.
Дата добавления: 2019-02-12; просмотров: 269; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!