Механизмы биоэлектрогенеза и его роль в возбуждении. Физико-химические свойства биоэлектрогенеза (БЭГ)
Живая ткань обладает не только пассивными, но и активными электрическими свойствами, являясь источниками электромагнитной энергии. Природа БЭГ была установлена только после появления теории электрической диссоциации, которая была разработана в 1887 г. Аррениусом. Живые ткани обладают свойствами электролита, которые диссоциируются на катионы и анионы. В этой связи, самой простой моделью источника электромагнитной энергии биологических тканей служит концентрационный элемент Нернста. В нем растворы соли разной концентрации разделены мембраной, имеющей неодинаковую проницаемость для катионов и анионов, на которые диссоциирует данная соль. Вследствие существования разности концентрации, электролит стремится диффундировать через мембрану. Но, в силу неодинаковой проникающей способности образующих его ионов, один из них преодолевает мембрану, а другой задерживается ею. В результате, на мембране образуется двойной слой зарядов. При этом, более разбавленный раствор принимает заряд того иона, который лучше проникает сквозь мембрану. Если это катион, то электрический ток во внешней цепи концентрационного электролита течет от электрода, погруженного в раствор с меньшей концентрацией, к электроду, находящемуся в более крепком растворе. При одинаковой проникающей способности катионов и анионов, на которые диссоциирует электролит, электрическая энергия не генерируется, как бы ни был высок концентрационный градиент на мембране.
|
|
|
Схематически электролит Нернста можно представить:

 - проницаемость катионов
- проницаемость катионов
 - проницаемость анионов
- проницаемость анионов
Электродвижущая сила, возникающая в концентрированном электролите, который образован раствором одной соли, определяется из уравнения Нернста:

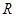 - универсальная газовая постоянная;
- универсальная газовая постоянная;
 - температура;
- температура;
 - валентность ионов;
- валентность ионов;
 - число Фарадея;
- число Фарадея;
 - концентрация анионов в первой области;
- концентрация анионов в первой области;
- проницаемость для катионов через мембрану;
 - концентрация катионов во второй области;
- концентрация катионов во второй области;
 - концентрация анионов в первой области;
- концентрация анионов в первой области;
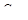 - концентрация анионов во второй области;
- концентрация анионов во второй области;
 - проницаемость для анионов через мембрану.
- проницаемость для анионов через мембрану.
Уравнение Нернста явилось результатом математической обработки многочисленных результатов экспериментального исследования функциональной зависимости ЭДС концентрационного элемента от соотношения концентрации соли в растворах, разделяемых мембраной. По Нернсту, десятикратная разность концентраций однозарядного иона создает ЭДС примерно 60 мВ (а двухзарядные ионы создают ЭДС примерно 30 мВ), следовательно, одновалентные ионы вносят наибольший вклад в БЭГ.
|
|
|
Наличие концентрационных градиентов является необходимым, но недостаточным условием БЭГ. Он обусловливается также тем, что сквозь БМ катионы проникают лучше, чем анионы. Концентрационные градиенты стремятся выровнять содержание всех ионов по обе стороны БМ. Однако, БМ препятствует этому процессу, поддерживая тем самым ионную асимметрию. Важный вклад в поддержание ионной асимметрии и соответствующих концентрационных градиентов (например, ионов 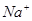 и
и  ) на плазмолемме любой клетки вносит их активный транспорт.
) на плазмолемме любой клетки вносит их активный транспорт.
Таким образом, при наличии БЭГ всегда соблюдается два обязательных условия:
. Существование концентрационных градиентов электролитов на клеточной мембране.
. Наличие неодинаковой проницаемости этой мембраны для катионов и анионов, на которые диссоциируют электролиты в живой клетке (внутри и вне клетки).
При рассмотрении живых тканей в качестве концентрационного элемента, необходимо учитывать диффузию через БМ не одного иона, а всех, концентрации которых неодинаковы внутри и вне клетки, и которые способны при этом проникать через БМ.
35. Потенциал покоя
Уравнение Гольдмана позволяет рассчитывать разность потенциалов, существующую у разных клеток между цитоплазмой и межклеточной средой в состоянии покоя и в состоянии возбуждения. Это уравнение имеет вид:
|
|
|

ЭДС в состоянии покоя называют потенциалом покоя (ПП),
в состоянии возбуждения - потенциалом действия (ПД). Результаты расчетов довольно близко совпадают с результатами экспериментов. Наилучшее совпадение имеет место для ПП.
Анализ результатов эксперимента показал, что в состоянии покоя проницаемость для 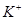 больше проницаемости для
больше проницаемости для  и
и  . Это позволяет считать проницаемости для ионов и анионов бесконечно малыми величинами по отношению к проницаемости для . В этом случае численное значение потенциала покоя можно рассчитывать по формуле:
. Это позволяет считать проницаемости для ионов и анионов бесконечно малыми величинами по отношению к проницаемости для . В этом случае численное значение потенциала покоя можно рассчитывать по формуле:

Необходимо отметить, что ПП возник не за счет более быстрой диффузии ионов по сравнению с 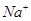 . Разница между проницаемостями и определяет только направление катионного потока, который создает ПП на БМ. Поскольку содержание в цитоплазме больше, чем в межклеточной среде, то катионный поток направлен из клетки наружу (выходящий ток). Соотношение концентрации ионов в межклеточной среде и внутри клетки определяет амплитуду ПП. Причиной возникновения трансмембранной разности потенциалов в состоянии покоя, является скорость диффузии через БМ ионов и анионов высокомолекулярных органических веществ, находящихся в цитозоли. Катионы проходят сквозь плазмолемму, а анионы задерживаются ею, что приводит к образованию не плазмоалемме двойного электрического слоя зарядов. При этом, межклеточная среда, как более разбавленный раствор, приобретает "+" заряд, а цитоплазма "-".
. Разница между проницаемостями и определяет только направление катионного потока, который создает ПП на БМ. Поскольку содержание в цитоплазме больше, чем в межклеточной среде, то катионный поток направлен из клетки наружу (выходящий ток). Соотношение концентрации ионов в межклеточной среде и внутри клетки определяет амплитуду ПП. Причиной возникновения трансмембранной разности потенциалов в состоянии покоя, является скорость диффузии через БМ ионов и анионов высокомолекулярных органических веществ, находящихся в цитозоли. Катионы проходят сквозь плазмолемму, а анионы задерживаются ею, что приводит к образованию не плазмоалемме двойного электрического слоя зарядов. При этом, межклеточная среда, как более разбавленный раствор, приобретает "+" заряд, а цитоплазма "-".
|
|
|
Расчеты экспериментальных данных свидетельствуют о том, что все клетки организма в состоянии покоя характеризуются определяемым потенциалом (поляризацией). Клеточная мембрана всегда заряжена и всегда на ее внутренней поверхности поддерживается "-" заряд (разность потенциала), а на внешней - "+" заряд. Трансмембранная разность потенциалов в разных клетках различна, но всегда составляет несколько десятков мВ. Так, например, для гигантского аксона кальмара потенциал покоя составляет примерно -85 мВ, в то время, как для мембран эпителиальных клеток ПП - в 2-3 раза ниже. Незначительная на первый взгляд величина потенциала покоя создает высокий градиент потенциала на плазмолемме, так как толщина БМ составляет примерно 10 нм, следовательно, напряженность электрического поля на плазмолемме может достигать  В/м.
В/м.


Избирательная проницаемость плазмолеммы каждой клетки обеспечивает стабильное разобщение катионов и анионов, вследствие чего, на ней стойко поддерживается высокий градиент потенциала, и она обладает ЭДС. Эта ЭДС направлено против причины ее вызывающей и противодействует дальнейшему разобщению зарядов на БМ. Существование ионных градиентов на БМ связано с работой системы активного транспорта и является энергоемким процессом. В этой связи, потенциал покоя частично экономит энергетические затраты свободной энергии на поддержание этих градиентов, так как потенциал покоя действует в противоположном направлении концентрационному градиенту.
. Потенциал действия
При действии на живые ткани раздражителей, изменяются условия генерации ЭДС в клеточных концентрационных элементах, что выражается в возникновении ПД. Согласно натриевой теории (гипотезе), при возбуждении нервных или мышечных волокон, изменяется проницаемость плазмолеммы клеточных структур для ионов и . Мгновенно, на короткий промежуток времени, проницаемость для ионов и становится равным, как 1:30.

При этом, проницаемость для оказывается намного больше проницаемости для анионов, с которыми ион образует соли в межклеточной среде. Следовательно, в момент возбуждения, проницаемость для ионов намного больше проницаемости для ионов и анионов. Это позволяет принять проницаемости для ионов  и анионов за бесконечно малые величины. После преобразования уравнения Гольдмана, получаем формулу для расчета так называемого натриевого потенциала. Он равен превышению по абсолютной величине потенциала действия над потенциалом покоя.
и анионов за бесконечно малые величины. После преобразования уравнения Гольдмана, получаем формулу для расчета так называемого натриевого потенциала. Он равен превышению по абсолютной величине потенциала действия над потенциалом покоя.
Разность модулей ПД и ПП называют потенциалом инверсии. В аксоне он эквивалентен натриевому потенциалу, то есть:

Численное значение потенциала инверсии неодинаково в разных клетках, но всегда составляет десятки мВ, и знак этого потенциала положительный, относительно потенциала покоя:

Как правило, потенциал  меньше ПП по модулю. Так, например, в гигантском аксоне кальмара примерно равен +55 мВ, а в миакардиальных волокнах он составляет примерно +20 мВ (для сердца человека). ПД определяется суммой абсолютных значений ПП и .
меньше ПП по модулю. Так, например, в гигантском аксоне кальмара примерно равен +55 мВ, а в миакардиальных волокнах он составляет примерно +20 мВ (для сердца человека). ПД определяется суммой абсолютных значений ПП и .


Продолжительность существования ПД в большинстве нервных и мышечных волокон невелика (в пределах долей единиц мс.), причем, не требуется никаких воздействий, чтобы нейрон или мышечное волокно вернулось к исходному состоянию поляризации, то есть, приобрело обычный ПП. Возбудимый участок цитоплазмы приобретает положительный потенциал, относительно межклеточной среды, и такое состояние называют деполяризацией. Возвращение к прежней поляризации называют реполяризацией.
ПД создается за счет более быстрой диффузии сквозь плазмолемму ионов , по сравнению с анионами, которые образуют с анионами соли в межклеточной среде, следовательно, деполяризация (появление 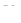 ) связана с вхождением в цитоплазму ионов .
) связана с вхождением в цитоплазму ионов .
Возникновение ПД обусловлено не нарушением ионных концентраций в цитозоли, а падением электрического сопротивления плазматической мембраны, вследствие повышения ее проницаемости для ионов . Так, при деполяризации сопротивление плазмолеммы уменьшается примерно в 40 раз. Входящий поток ионов является результатом его пассивного транспорта по мембранным каналам под действием концентрационного и электрического градиентов, а выходящий поток ионов обеспечивается работой натриевой помпы (активный транспорт). В состоянии покоя, встречные потоки ионов уравновешенны, в то время, как при возбуждении в течение времени существования ПД, система встречных потоков нарушается, и ионы устремляются из межклеточной среды внутрь клетки. При этом система активного транспорта оказывается неспособна моментально ликвидировать (компенсировать) резкое усиление входящего потока. Она делает это с некоторым запозданием, в фазу реполяризации. Мембранные потенциалы являются не равновесными, а стационарными, так как они поддерживаются в условиях существования встречных потоков через БМ. Сдвиги мембранных потенциалов связаны с нарушением действия стационарного режима, причем, возбуждение сопровождается усилением входящего и выходящего потоков ионов . Следовательно, возбуждение не включает систему активного транспорта ионов , а напротив, активизирует ее, однако, даже при максимальной активизации работы натриевой помпы, она не может воспрепятствовать кратковременному накоплению небольшого количества ионов в цитоплазме. В промежутках между последовательными генерациями ПД (в состоянии покоя)  помпа откачивает из цитоплазмы избыток ионов , который образовался при возбуждении, и ликвидирует созданный дефицит ионов .
помпа откачивает из цитоплазмы избыток ионов , который образовался при возбуждении, и ликвидирует созданный дефицит ионов .
Дата добавления: 2019-02-12; просмотров: 316; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!