Суммарная реакция фотосинтеза
СО2 +Н2О = (СН2О) + О2.
Химию фотосинтеза описывают следующими уравнениями:

Фотосинтез – 2 группы реакций:
· световая стадия(зависят отосвещенности)
· темновая стадия (зависит от температуры).
Обе группы реакций протекают одновременно
Фотосинтез происходит в хлоропластах зеленых растений.

Фотосинтез начинается с улавливания и поглощения света пигментом хлорофиллом, содержащимся в хлоропластах клеток зеленых растений.
Хлорофилл растений представлен двумя формами – a и b.
Молекула хлорофилла поглощает фотоны в фиолетовой и синей, а затем в красной части спектра, и не взаимодействует с фотонами в зеленой и желтой части спектра.
Пигменты организованы в фотосистемы (антенные поля по улавливанию света) – содержащие по 250–400 молекул разных пигментов.
Фотосистема состоит из:
· реакционного центра фотосистемы (молекула хлорофилла а),
· антенных молекул
· фотосистема 1 (максимум поглощения света на длине волны 700 нм - Р700)
· фотосистема 2 (максимум поглощения света на длине волны 680 нм - Р680
Различия в оптимумах поглощения обусловлены небольшими различиями в структуре пигментов.
Две системы работают сопряженно, как конвейер, состоящий из двух частей и называющийся нециклическим фотофосфорилированием.
Суммарное уравнение для нециклического фотофосфорилирования:
|
|
|
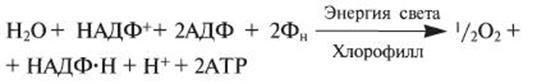
Ф - условное обозначение остатка фосфорной кислоты

Цикл начинается с фотосистемы 2.
1) антенные молекулы улавливают фотон и передают возбуждение молекуле активного центра Р680;
2) возбужденная молекула Р680 отдает два электрона кофактору Q при этом она окисляется и приобретает положительный заряд;
Кофактор (cofactor). Кофермент или любое другое вещество, необходимое для выполнения ферментом его функции
Коферменты (коэнзимы) [от лат. co (cum) — вместе и ферменты], органические соединения небелковой природы, участвующие в ферментативной реакции в качестве акцепторов отдельных атомов или атомных групп, отщепляемых ферментом от молекулы субстрата, т.е. для осуществления каталитического действия ферментов. Эти веществава, в отличие от белкового компонента фермента (апофермента), имеют сравнительно небольшую молекулярную массу и, как правило, термостабильны. Иногда под Коферментами подразумевают любые низкомолекулярные вещества, участие которых необходимо для проявления каталитического действия фермента, в т. ч. и ионы, напр. К+, Mg2+ и Мn2+ . Располагаются оферменты. в активном центре фермента и вместе с субстратом и функциональными группами активного центра образуют активированный комплекс.
|
|
|
Для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Исключение составляют гидролитические ферменты (например, протеазы, липазы, рибонуклеаза), выполняющие свою функцию в отсутствие кофермента.
Молекула восстанавливается Р680 (под действием ферментов),. При этом вода диссоциирует на протоны и молекулярный кислород, т.е. вода является донором электронов, который обеспечивает восполнение электронов в Р680.
ФОТОЛИЗ ВОДЫ — расщепление молекулы воды, в частности в процессе фотосинтеза. Вследствие фотолиза воды образуется кислород, выделяющийся зелеными растениями на свету.
Общий ход фотолиза воды.

Уравнение фотолиза воды
hv
2Н20=4Н+ 4е- +02
Для создания одной молекулы кислорода нужно восстановить две молекулы Р680, потерявшие в сумме четыре электрона, при этом образуются четыре протона.
3) эти протоны образуются во внутреннем пространстве тилакоида, где создается избыточная концентрация протонов по сравнению с окружающим пространством (т. е. более кислая среда), (формируются - протонный градиент и мембранный потенциал). АТФ-синтетаза парами выпускет протоны наружу и синтезирует АТФ из АДФ.
|
|
|
4) два электрона, поступившие к кофактору Q, передаются далее по цепочке белков, которая очень похожа на цепь переноса электронов.
5) электроны поступают от пластоцианина к реакционному центру фотосистемы 1 – молекуле Р700.
Фотосистема 1:
1) реакционный центр Р700 отдает 2 e акцепторному железосодержащему белку (Р430), окисляется и приобретает положительный заряд;
2) эта молекула восстанавливается и теряет заряд, получив два «успокоившихся» (но не до исходного состояния – их энергия еще не до конца израсходована!) электрона, изначально поступивших от фотосистемы 2. В этом случае необходимости в фотолизе нет и его не происходит;
3) Р430 отдает электроны белку ферродоксину;
4) получив электроны, этот белок восстанавливает кофермент НАДФ+ до НАДФ∙Н. Данный кофермент представляет собой фосфорилированный НАД. Процесс происходит на внешней мембране тилакоида.
Таким образом, по цепи белков передаются электроны, в результате чего во внутреннем пространстве тилакоида создается избыточная концентрация протонов, а на мембране – разность потенциалов.
Возникающая потенциальная энергия используется для синтеза АТФ за счет движения протонов по градиенту, осуществляемого АТФ-синтетазой. (т.о. энергия потока используется для получения АТФ)
|
|
|
Для возбуждения электронов используется свет
НАДФ + используется в темновой стадии фотосинтеза (а может быть далее использован и митохондриях).
В целом получается, что протоны образуются во внутреннем пространстве тилакоида при фотолизе воды, закачиваются туда же в ходе работы фотосистемы 2 и черпаются из внешнего пространства тилакоида для восстановления НАДФ+ до НАДФ-Н (т.е. электроны, движущиеся вниз по градиенту энергии к НАДФ вдоль цепи переноса электронов, взаимодействуют с ионами водорода (из воды), образуя восстановленный НАДФ-)
Фотосистема 1 может работать автономно.
Циклическое фотофосфорилирование
При этом используется обходной путь переноса электронов от возбужденного реакционного центра – а именно та же цепь переноса электронов, которая ведет из фотосистемы 2. Электроны проходят по ней возвращаются обратно к реакционному центру фотосистемы 1 – Р700. Т.о. электроны возвращаются обратно к Р700 по другой цепи переноса электронов. Как и при нециклическом фосфорилировании энергия возбуждения электронов, перемещающихся вдоль этой цепи, направляется на получение АТФ.
Оно может идти параллельно с нециклическим. Кроме того, оно используется некоторыми фотосинтетическими бактериями, которые в процессе фотосинтеза не выделяют кислорода.

Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами:
1) синтезом АТФ;
2) образованием НАДФ·Н2;
3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
количество АТФ, синтезируемое АТФ-синтетазой, не связано жесткой зависимостью от количества электронов, пропущенных по цепочке белков в фотосистеме II.
Темновая фаза
Протекает в строме хлоропласта.
Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте.
Реакции темновой фазы представляют цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
В настоящее время различают два типа фотосинтеза: С3- и С4-фотосинтез. Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин).
Характерные особенности С3-фотосинтеза:
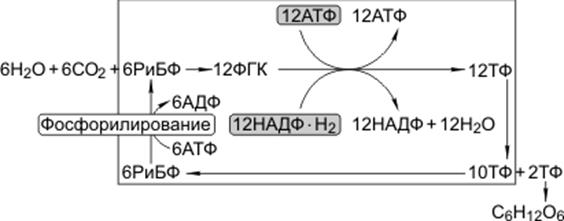
1) Первая реакция в этой цепочке - фиксация углекислого газа. Акцептором углекислого газа является рибулозодифосфат,
2) реакцию карбоксилирования рибулозодифосфата катализирует рибулозодифосфат-карбоксилаза,
3) в результате карбоксилирования рибулозодифосфат образуется шестиуглеродное соединение, которое распадается на две фосфоглицериновой кислоты.
фосфоглицериновая кислота восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию рибулозодифосфата, часть превращается в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу;
цикл этих реакций получил название цикл Кальвина:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды.
С3-фотосинтез
Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом.
К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ.
АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
Бактериальный фотосинтез
Фотосинтез бактериальный — фотосинтез, осуществляемый бактериальными микроорганизмами.
типично водные микроорганизмы, распространенные в пресных и соленых водоемах. Особенно часто они встречаются в местах, где есть сероводород, как в мелководье, так и на значительной глубине. В почве фототрофных бактерий мало, но при затоплении ее водой они могут расти весьма интенсивно
пурпурные и зеленые бактерии — наиболее древние фотосинтезирующие организмы
Всего сейчас описано около пятидесяти видов пурпурных и зеленых бактерий.
Содержат развитую систему мембран, некоторые из них выполняют функцию фотосинтезирующего аппарата (хроматофоры или тилакоидами
В отличие от растений, фотосинтез у пурпурных и зеленых бактерий
не сопровождается выделением кислорода
Донорами водорода при ассимиляции углекислоты служит не вода, а другие вещества:
· сероводород (2C02+H2S+2H20 -> 2(CH20)+H2S04),
· молекулярная сера,
· тиосульфат,
· сульфит,
· молекулярный водород,
· органические соединения.
Обязательными компонентами сред, кроме источников углерода и азота, естественно, являются фосфор, сера, калий и магний в виде минеральных солей. Установлено также, что эти микроорганизмы нуждаются в довольно большом количестве железа, но по сравнению с растениями проявляют значительно меньшую потребность в марганце. Рост их зависит от концентрации кальция и наличия в небольших количествах таких элементов, как Mo, Co, Zn, Си и, видимо, других.
У многих пурпурных и зеленых бактерий установлена способность фиксировать молекулярный азот.
У синезеленых водорослей (цианобактерий), как и у растений, в процессе бактериального фотосинтеза в качестве донора водорода образуется вода и выделяется кислород.
Дата добавления: 2018-10-26; просмотров: 481; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!