Размножение - универсальное свойство живого, обеспечивающее материальную непрерывность в ряду поколений. Эволюция размножения, формы размножения.
Размножение – это способность организмов производить себе подобных особей того же вида. Существует два типа размножения: половое и бесполое.
Древнейшим способом размножения на Земле было бесполое размножение.
Бесполое размножение.
Бесполое размножение происходит без образования специальных клеток, в нём участвует один организм, одна особь, при этом размножении образуются идентичные потомки. Единственным источником генетической изменчивости являются случайные мутации. Цитологической основой бесполого размножения является митоз. Молекулярной основой бесполого размножения является репликация ДНК. Бесполое размножение у различных живых организмов может происходить по-разному. Формы бесполого размножения:
1. Почкование – это форма бесполого размножения при которой новая особь образуется в виде выростов (почки) на теле родительской особи, а затем отделяется от неё и превращается в самостоятельную особь (гидра, дрожжи).
2. Фрагментация – это разделение особи на две или более частей, каждая из которых растёт и образуется отдельная особь (высшие растения, губка, дождевой червь).
3. Образование спор. Спора – это одноклеточная репродуктивная единица, состоящая из ядра и небольшого количества цитоплазмы под плотной оболочкой. Из споры образуется новая особь (низшие растения).
4. Деление. Бинарное деление клетки на две части. Ядро родительской особи один или несколько раз делится митозом, при этом образуется два или несколько дочерних ядер. Каждое из них окружается цитоплазмой и развивается в самостоятельный организм.
5. Шизогония – это множественное деление клетки. Сначала в клетке многократно делится ядро, затем вокруг каждого ядра обособляется участок цитоплазмы, который окружается плазматической мембраной. Затем происходит распад на отдельные клетки (малярийный плазмодий).
6. Вегетативное размножение. Осуществляется формирование дочернего организма из группы клеток материнского организма. У растений это размножение происходит за счёт вегетативных органов: корневищ, луковиц, клубней, усов.
В результате бесполого размножения образуются генетически идентичные особи. Скорость размножения очень высокая и в постоянных условиях организма быстро захватывают экологическую нишу.
Половое размножение.
Появилось половое размножение более 3 млрд. лет назад. Сущность полового размножения в перекомбинации генетического материала родительских особей. В результате дочерние особи становятся более разнообразными, и естественный отбор выбирает из них наиболее приспособленные. При половом размножении потомство получается в результате слияния гаплоидных клеток – гамет. При оплодотворении образуется зигота. Из которой развивается новый организм.
Оплодотворение – это процесс слияния сперматозоида с яйцеклеткой с последующим слиянием их ядер и образованием диплоидной зиготы. Биологическое значение этого процесса состоит в том, что при слиянии мужских и женских гамет образуется новый организм, несущий признак обоих родительских организмов.
Гаметы гаплоидны, они содержат половинный набор хромосом и образуются в результате мейоза.
Одной из модификаций полового размножения является партеногенез.
Партеногенез – это процесс, при котором женская гамета развивается в новую особь без оплодотворения (встречается у животных (пчёлы) и растений). Преимущество в том, что увеличивается скорость размножения.
Эволюция размножения стала важной движущей силой общей эволюции царства растений и привела к возникновению новых специализированных органов. Бесполое размножение — наиболее древний и, несомненно, изначальный способ размножения. Оно представлено у всех групп растений, включая покрытосеменные
Наиболее примитивный способ бесполого размножения — вегетативное. Его наблюдают на самых ранних этапах развития жизни — у доклеточных. У многих предъядерных (например, у бактерий) это и сейчас единственный способ размножения. Каждая крупная естественная группа растений нередко обладает свойственными только ей формами вегетативного размножения.
Так, у сине-зеленых водорослей это гормогонии, у лишайников — изидии и соредии, у грибов — хламидоспоры и почкование, у некоторых моховидных— выводковые тельца. Наиболее разнообразны формы вегетативного размножения у высших растений, особенно у покрытосеменных. Велико разнообразие спор собственно бесполого размножения. В процессе эволюции они утратили способность воспроизводить новую особь, сходную с материнской (спорофит), а дают начало половому поколению (гаметофиту). Половое размножение возникло на очень ранних этапах эволюции, но время его появления точно не установлено.
У некоторых современных примитивных групп растений половой процесс неизвестен. Не вызывает сомнений, что такие природные группы, как сине-зеленые водоросли и большинство бактерий, никогда не имели полового размножения. Другие, например некоторые грибы, возможно, утратили его. Половой процесс утрачен и у некоторых высокоорганизованных растений, например у покрытосеменных (партеногенез). В настоящее время наука еще не располагает достаточными данными для воспроизведения полной картины эволюции полового процесса у растений.
Твердо установлено, что у растений, стоящих на более низкой ступени эволюции, половой процесс протекает в более примитивной форме, чем у растений, стоящих на более высокой ступени эволюции. Один из показателей уровня эволюции — специализация гамет. Наиболее примитивен изогамный половой процесс. Более высокий уровень — гетерогамия.
Наиболее специализированным половым процессом считают оогамию. Мужская гамета теряет подвижность, приспособившись к наземным условиям жизни, и доставляется к яйцеклетке пыльцевой трубкой. Таким образом, у семенных растений половой процесс не связан с наличием водной среды, это — приспособление к сухопутному образу жизни. Половое размножение развивалось не изолированно от бесполого, а во взаимной связи с ним. Каждая природная группа растений в жизненном цикле имеет свою закономерность смены бесполого и полового размножения. У многих низших растений смена ядерных фаз не выражена, спорофит почти полностью выпадает из жизненного цикла (диплоидна лишь зигота).
В жизненном цикле некоторых низших растений и всех высших существует чередование спорофита и гаметофита. Степень их морфологической разнородности и преобладания в жизненном цикле служит важным показателем эволюционного уровня изучаемой группы растений. На основании работ В. Гофмейстера (1849, 1851) установлена важная общая закономерность: в процессе эволюции спорофит приобретает всё большее развитие и самостоятельность; гаметофит, напротив, все более редуцируется, полностью теряет свою самостоятельность и всецело зависит от спорофита, что и наблюдают у наиболее высокоорганизованных семенных растений — голосеменных и покрытосеменных. В. Гофмейстер, изучая жизненные циклы различных природных групп растений, показал, что между высшими споровыми и покрытосеменными нет такого большого разрыва, как предполагали ранее.
100. Пути происхождения различных групп паразитов. Принципы взаимодействия паразита и хозяина на уровне особей. Пути морфофизиологической адаптации паразитов.
Наиболее просто объясняется происхождение эктопаразитизма. Один из путей к этому — через увеличение количества источников питания с последующей их сменой. Так, многие насекомые имеют колюще-сосущий ротовой аппарат, питаясь соками растений. Но питание за счет прокалывания ткани и всасывания жидкости и есть способ поглощения пищи всеми кровососущими членистоногими, ряд которых, потребляя кровь человека и теплокровных животных, продолжает пользоваться также и соками растений, s
Другой путь, ведущий к эктопаразитизму, — хищничество. Активные хищники, осваивающие для питания все более крупные жертвы, становятся вначале временными, а затем и постоянными эктопаразитами за счет удлинения контактов с организмом хозяина. Так, многие пиявки, ведущие себя как хищники по отношению к мелким организмам, становятся паразитами более крупных животных, питаясь их кровью. Увеличение продолжительности питания — основное направление перехода от временного к постоянному эктопаразитизму. Действительно, из большого количества кровососущих форм членистоногих наиболее длительное питание на хозяине характерно именно для постоянных паразитов, степень контакта которых с хозяевами наиболее высока.
Иной путь возникновения эктопаразитизма — через усиление контакта так называемых гнездовых паразитов с поверхностью тела хозяина. Животные, обитающие в убежище другого вида, могут питаться его перьями, волосами и отпадающими чешуйками кожного эпидермиса. Переход к постоянному обитанию на поверхности тела хозяина дает паразиту большие преимущества. Возможно, так возник паразитизм пухоедов, власоедов птиц и млекопитающих и группы клещей — обитателей эпидермиса животных и человека.
Основная масса случаев эндопаразитизма в полостных органах, имеющих связь с внешней средой, представляет собой явление, развившееся в результате случайного заноса в организм цист, яиц или личинок свободноживущих видов, предварительно имеющих адаптации к обитанию в почве или в воде, содержащей избыток органического вещества. Примером является угрица кишечная, которая в своем развитии сохранила возможность обитать и размножаться как в почве, так и в организме человека (см. разд. 20.2.1.1).
Возможен переход к паразитированию в одном хозяине после предварительной адаптации к обитанию в другом, служащем источником питания первого. Так, известен целый ряд гельминтов, которые, обитая в кишечнике рыбы, не перевариваются в пищеварительной системе хищников, съевших паразитов вместе с хозяином и продолжающих паразитировать в кишечнике или тканях уже нового вида (см. разд. 20.1.2.1).
Не исключается и вариант перехода к полостному паразитизму видов, предварительно адаптированных к эктопаразитизму. Этим путем, вероятно, эволюционируют некоторые насекомые, большую часть цикла развития проводящие в ротовой полости птиц, но выходящие для размножения на перьевой покров их головы.
Наиболее сложно и многообразно происхождение паразитов тканей внутренней среды. Один из путей — через изменение инстинкта откладки яиц и предварительных адаптации к эктопаразитизму. Таким путем, вероятно, произошел тканевой паразитизм личинок мух и оводов (см. разд. 21.2.4), откладывающих яйца на поверхности кожи и слизистых оболочек животных и человека. Личинки при этом вскоре погружаются под покровы и ведут типичный эндопаразитический образ жизни.
Многие паразиты приспособились к обитанию в тканях после освоения полостных органов, связанных с внешней средой. Так, по-видимому, шла эволюция паразитизма у ленточных червей, у трихинеллы спиральной (см. разд. 20.2.1.2). В цикле развития этих паразитов имеются формы, обитающие как в кишечнике, так и в тканях.
Некоторые паразиты внутренней среды возникли, вероятно, предварительно адаптировавшись к обитанию в пищеварительной системе членистоногих, а с переходом последних к гематофагии заселили новую и труднодоступную экологическую нишу — кровь и другие ткани мезодермального происхождения.
Таким образом, путей перехода к паразитизму у разных видов животных много, но несомненным остается одно: паразитизм — явление вторичное. Об этом свидетельствует наличие в жизненных циклах многих, даже наиболее специализированных паразитов, свободноживущих стадий, рекапитулирующих свободный образ жизни предков (см., например, разд. 20.1.1).
Переход к паразитическому образу жизни сопровождается появлением у паразитов ряда адаптации, облегчающих их существование, развитие и размножение в специфических условиях организма хозяина Разнообразие форм паразитизма, различное систематическое положение паразитов (их принадлежность к разным отрядам, классам и типам), а также обитание их в разных органах и системах хозяина обусловливают многообразие этих адаптации.
Однако некоторые приспособления являются абсолютно универсальными. К ним в первую очередь относятся высокая плодовитость иособенности половой системы. Действительно, возможность оставления потомства и попадания его в благоприятную среду — организм хозяина — у паразитов часто ничтожна. В связи с этим интенсивность размножения паразитов по сравнению со свободноживущими формами гораздо более велика. Достигается это разными способами. У многоклеточных это сильная степень развития половой системы и образование огромного количества половых продуктов. Этому способствуют первичный гермафродитизм плоских червей, изначально высокая плодовитость круглых червей и основной массы членистоногих. Нередко высокая интенсивность полового размножения дополняется размножением личиночных стадий жизненного цикла. Особенно это характерно для сосальщиков, личинки которых размножаются партеногенетически, а у некоторых ленточных червей — внутренним или наружным почкованием.

Рис. 18.4. Органы прикрепления паразитов
Присасывательные диски у инфузории (Л) и лямблии (Б) (указаны стрелками), сложная присоска (В) и кутикулярные шипики на коже (Г) у сосальщиков, крючья и сложные присоски на головках ленточных червей (Д, Е, Ж), конечности, служащие для прикрепления, у ракообразного (3) и насекомого (И)
Некоторые паразиты из типа простейших приобретают способность к множественному делению — шизогонии, когда из одного паразита может образоваться более 1000 дочерних особей, или к спорогонии, в результате которой из одной особи могут образоваться десятки тысяч организмов следующего поколения. Практически у всех эктопаразитов и паразитов, обитающих в полостных органах, имеются адаптации для прикрепления к телу хозяина. Они встречаются у простейших (присасывательные диски лямблии), у гельминтов (присоски, шипики, крючья плоских червей, хитинизированный ротовой аппарат ряда круглых червей) и паразитических членистоногих (своеобразные конечности) (рис. 18.4).
Эндопаразиты, обитающие в полостных органах, имеют покровы, обладающие антиферментными свойствами, быстро регенерирующие либо вообще непроницаемые для ферментов хозяина. Паразиты, живущие в тканях, часто там инкапсулируются (рис. 18.5).
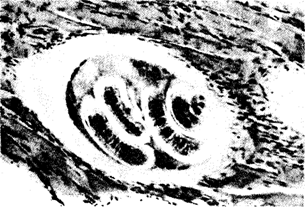
Рис. 18.5. Личинка трихинеллы, инкапсулированная в мышце
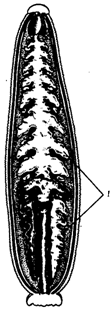
Рис. 18.6. Пищеварительная система пиявки:
1 – слепые выросты кишечника
Паразиты, питающиеся кровью (представлены в основном членистоногими), имеют колюще-сосущий ротовой аппарат, а также сильно растяжимый хитиновый покров, часто разветвленную пищеварительную трубку (рис. 18.6), антикоагулянтные свойства слюны и консервантные свойства ферментов пищеварительной системы.
Эндопаразиты, активно отыскивающие хозяина, обладают органами ориентации в среде, используемыми для поисков хозяина (светочувствительные глазки, термо- и хеморецепторы), и органами передвижения (рис. 18.7).
Передний конец тела паразитов, внедряющихся в организм хозяина, снабжен органами проникновения — специализированными железами, колющими стилетами и т.д. Это касается даже некоторых простейших, способных проникать в ткани хозяина через неповрежденные покровы.
Все паразиты, развивающиеся со сменой хозяев, используют в качестве таковых виды, связанные между собой непосредственными пищевыми взаимоотношениями или обитающие с ними в одной среде. Большинство промежуточных хозяев являются источником питания для основных. Другой распространенный путь попадания паразита в организм хозяина — это использование многочисленных переносчиков, которые обеспечивают не только постоянную циркуляцию паразитов в экологических системах, но и их широкое расселение.
Морфофизиологические и биологические адаптации паразитов
Условия сохранения жизни хозяина требуют от паразита максимальной приспособленности к хозяину при возможно меньших нарушениях его жизненных функций. Чем теснее контакт паразита и хозяина, тем сильнее выражен регресс ряда систем органов паразита, который сопровождается развитием специализированных структур (органы фиксации), усложнением отдельных систем (например, половой).
Морфофизиологические адаптации связаны с изменением внешнего и внутреннего строения паразитов и функционирования их систем органов. Они подразделяются на:
- прогрессивные адаптации: наличие органов фиксации (присоски, крючья, коготки вшей, ротовой аппарат клещей); сложное строение наружных покровов (кутикула, тегумент); молекулярная "мимикрия" (сходство структуры белков и ферментов паразита и хозяина); выделение кишечными паразитами антиферментов (зашита от переваривания соками хозяина); внутриклеточное паразитирование; иммунносупрессивное действие паразитов (эндопаразиты секретируют протеазы, разрушающие иммунные комплексы и клетки хозяина) и др.
- регрессивные: редукция органов движения и некоторых систем (кровеносной, дыхательной); упрощение строения нервной системы и органов чувств.
1) Механическое повреждение органов и тканей: мигрирующие личинки аскарид нарушают целостность стенки кишечника и альвеол легких, клубки аскарид могут вызывать непроходимость кишечника, паразитирующий в печени кошачий сосальщик - закупорку желчных ходов. Присоски ленточных червей, ущемляя слизистую кишечника, приводят к некрозу тканей. Хоботки клещей и насекомых повреждают кожные покровы.
Токсико-аллергическое действие оказывают продукты жизнедеятельности или распада погибших паразитов. Приступ малярии связан с выходом в кровь продуктов метаболизма плазмодиев при разрушении эритроцитов. Массовая гибель личинок трихинелл при введении антигельминтиков может вызвать смерть больного от анафилактического шока. При "укусах" насекомых-эктопаразитов человек ощущает зуд в результате токсического действия их слюны на нервные окончания в коже. Кожные высыпания, зозинофилия, головные боли являются следствием действия продуктов обмена циркулирующих в крови личинок гельминтов.
Поглощение питательных веществ и витаминов из организма хозяина. Гельминтозы обычно сопровождаются гиповитаминозами (А и С). Широкий лентец в организме человека избирательно поглощает витамин В12, что приводит к развитию анемии. Чем больше масса тела эндопаразита, тем большее количество питательных веществ он поглощает в организме хозяина (например, широкий лентец и бычий цепень).
4).Открытие путей для вторичной инфекции: гельминты или их личинки, нарушая целостность кожных покровов или слизистой стенки кишечника, способствуют проникновению микроорганизмов.
Нарушение всех процессов обмена веществ у хозяина (белкового, углеводного, жирового и др.), общее ослабление организма, снижение его сопротивляемости и повышение чувствительности к другим заболеваниям (аскаридоз часто сочетается с дизентерией, брюшной тиф - с гельминтозами).
Паразиты играют важную роль в стимулировании иммунной системы, поддержании ее на высоком уровне и в конечном итоге в охране гомеостаза хозяина.
Государственное бюджетное образовательное учреждение
высшего профессионального образования
«Волгоградский государственный медицинский университет»
Министерства здравоохранения Российской Федерации
Кафедра биологии
Факультет лечебный
Дисциплина «Биология»
Экзаменационный билет №33
1. Биологическая изменчивость людей и биогеографическая характеристика среды. Экологическая дифференцировка человечества. Понятие об экологических типах людей и условиях их формирования.
2. Характеристика ово- и сперматогенеза. Строение сперматозоида. Строение и типы яйцеклеток у животных и человека.
3. Эхинококк и альвеококк. Систематическое положение, морфология, циклы развития, пути заражения, диагностика, профилактика. Отличие личиночных стадий развития.
М.П. Зав. кафедрой ___________
98.
Дата добавления: 2018-09-20; просмотров: 497; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!