Естественный отбор в дикой природе
За отбором гораздо легче следить в теории с помощью математических расчетов, чем в реальной ситуации в дикой природе. Кроме сложностей, связанных с контролем внешних условий, существует два основных осложняющих фактора, влияние которых ощущается очень сильно. Первый фактор – время. Если процессы протекают слишком медленно, то за время проведения эксперимента никаких изменений зафиксировать не удается. Вторая сложность – количество необходимых измерений. Для детекции слабых селективных преимуществ или недостатков необходим большой объем данных.
Второй фактор связан с вероятностью и статистикой. Если различие в приспособленности двух форм одного и того же вида очень незначительное, то для преодоления ошибки эксперимента и случайных вариаций необходимо провести измерения на большом количестве особей. Проиллюстрируем это на простом примере.
Предположим, мы хотим определить, имеет ли у какого‑то вида животных один вариант окраски преимущество над другим. Сколько животных нужно осмотреть, чтобы выявить отклонение от некоего ожидаемого соотношения? Допустим, речь идет о многочисленной популяции, скажем, о популяции рыб, которых легко выловить и посчитать. Теория вероятности гласит, что чем большее число особей мы изучим, тем точнее полученный результат будет отражать реальное количество каждого варианта окраски рыб в популяции. Сколько нам нужно рыб, чтобы получить результат с доверительной вероятностью 95 % (что означает, что в 95 случаях из 100 истинное значение оцениваемой величины попадет в определенный нами интервал)? Как видно из представленной ниже таблички, погрешность эксперимента уменьшается при увеличении числа измерений.
|
|
|
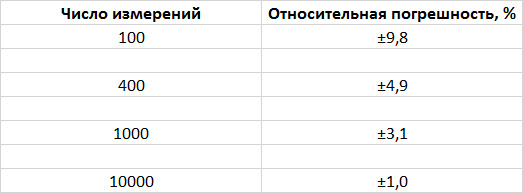
Если мы осмотрим всего 100 рыб, погрешность может достигать 10 %. При такой высокой вероятности ошибки слабых изменений не различить. (Такая же проблема возникает при опросах на выходе с избирательных участков: выборка слишком мала, и поэтому предсказания результатов иногда бывают ошибочными.)
Поскольку выявить слабые различия среди диких животных так сложно, большинство известных примеров касается действия очень сильного и, следовательно, очень быстрого отбора. Самый известный пример – меланизм у березовой пяденицы. Индустриальная революция привела к загрязнению отдельных районов Англии и Северной Америки, что затруднило рост лишайников и изменило цвет стволов деревьев, на которых пяденица скрывается от птиц. В результате произошло очень резкое и быстрое увеличение численности темноокрашенных форм бабочек и такое же быстрое сокращение численности светлоокрашенных форм. Всего за 50 лет, примерно с 1848 по 1896 г., количество темноокрашенных особей, изначально очень редких, в некоторых районах достигло 98 %. Холдейн проанализировал статистические данные по двум формам бабочек и обнаружил, что коэффициент отбора светлых бабочек на темных деревьях составил примерно ‑0,2. Может показаться, что преимущество в 20 % – не так уж много, но, нарастая год за годом подобно кумулятивной процентной ставке, такое преимущество обеспечивает очень быстрое распространение формы в популяции. Во второй половине XX в. были приняты законы об охране окружающей среды, отбор изменил свое направление, и в некоторых районах было зарегистрировано сокращение доли темноокрашенных особей березовой пяденицы от 90 до 10 % (рис. 2.2).
|
|
|

Рис. 2.2. Сокращение численности темноокрашенной формы березовой пяденицы в результате улучшения качества воздуха. При изменении внешних условий доля темноокрашенного варианта пяденицы значительно сократилась как в США [черные квадратики], так и в Великобритании [черные кружочки]. Светлые кружочки соответствуют теоретической кривой изменений численности при значении коэффициента отбора s = ‑0,15. Рисунок взят из статьи B. S. Grant et at., 1996, Journal of Heredity 87: 3551 (с изм.).
|
|
|
Естественный отбор березовой пяденицы осуществляют птицы. И это вводит в нашу задачу еще одну переменную, которая усложняет изучение естественного отбора в дикой природе. Может потребоваться анализ не только большого количества бабочек, но и разнообразных факторов отбора, действующих на них. Если хищников несколько и если условия изменяются в зависимости от региона, времени суток и т. д., возникают дополнительные трудности. В случае березовой пяденицы быстрое изменение численности двух форм, происходившее одновременно на двух континентах и связанное с активным развитием промышленности, ясно указывает на то, что в данном случае шел естественный отбор по окраске.
История с березовой пяденицей – лишь один из известных примеров. Естественный отбор животных с различной окраской был описан для улиток, божьих коровок, мешотчатых прыгунов и других видов животных; факторы отбора в одних случаях удалось установить точно, в других – предположительно. Для некоторых из этих видов коэффициент отбора определенного типа окраски достаточно высокий (от 0,01 до 0,5).
Важно понять, что длительные исследования естественного отбора в дикой природе требуют героического упорства ученых, постоянной финансовой поддержки и благосклонности самой природы. Можно сказать, что в полевых условиях происходит естественный отбор среди ученых: работать остаются лишь самые стойкие.
|
|
|
Недавние исследования взаимоотношений соколов и одичавших голубей, длившиеся семь лет, свидетельствуют о том, какое необыкновенное терпение требуется для изучения естественного отбора в дикой природе. В районе города Дэвис в Калифорнии встречаются голуби с шестью вариантами окраски оперения, и эти голуби являются излюбленной пищей сокола‑сапсана. Все голуби имеют серо‑голубое оперение, но у одного варианта этих птиц в нижней части спины выше хвоста есть белое пятно, которого нет у других (рис. 2.3).

Рис. 2.3. Варианты окраски хвостовой части оперения у голубей. В популяции городских голубей имеются особи с белыми перьями в нижней части спины (слева); такая окраска дает им определенное преимущество при нападении сокола‑сапсана. Рисунок Джейми Кэрролл.
Группа исследователей под руководством Альберто Паллерони из Гарвардского университета в мельчайших деталях изучила воздушный бой между соколом и голубем. За семь лет ученые стали свидетелями 1485 (!) нападений пяти взрослых соколов на голубей (рис. 2.4). Ученые проанализировали связь окраски голубей с количеством и успешностью нападений соколов. Для этого они пометили и описали 5235 голубей – это очень большая выборка.

Рис. 2.4. Охота. При нападении сапсан может развивать скорость до 70 м/с. В данном случае голубю явно угрожает опасность. Фотография Роба Пальмера, печатается с любезного разрешения Альберто Паллерони.
Исследователи обнаружили, что соколы нападают на голубей с белым пятном на спине реже, чем на других голубей, и вероятность удачной охоты на них ниже. В этом регионе голуби с белым пятном составляют 20 % всех голубей, а среди жертв соколов на их долю приходится всего 2 %.
Паллерони с коллегами объяснили подобное преимущество голубей с белыми перьями на спине их поведением при нападении сокола. Когда при виде добычи сокол разгоняется до 70 м/с, голубь складывает крылья и «скатывается» с траектории движения сокола. Биологи предположили, что белое пятно отвлекает нападающего сокола, когда голубь начинает свой кувырок, и это дает голубю дополнительную долю секунды, чтобы увернуться от хищника.
Чтобы проверить свою гипотезу, исследователи изменили окраску 756 голубей, «пересадив» белое пятно на голубей без пятна (для этого они выстригали белые перья у голубей с пятном и приклеивали их серым голубям). Затем птиц выпускали и следили за их судьбой при нападении сокола. Ситуация изменилась кардинальным образом. Голуби, которые раньше имели пятно, теперь становились добычей соколов не реже, чем все остальные, а сизые голуби с приклеенными белыми перьями оказались защищенными.
В ходе этого потрясающего исследования ученые отметили постоянное увеличение доли голубей с белым пятном на спине среди всей популяции голубей. Они в реальном времени наблюдали эволюцию этих птиц в условиях естественного отбора.
Исследование естественного отбора в дикой природе не сводится исключительно к анализу таких простых признаков, как окраска. Прекрасный пример – изучение эволюции трехиглой колюшки. Судя по геологическим данным, при отступлении ледников в конце последнего ледникового периода морская колюшка, заселившая реки и озера, оказалась в изоляции. Эти пресноводные популяции стали постепенно отличаться от своих морских предков. Морская форма колюшки обычно имеет на боках непрерывный ряд из 30 и более костных пластинок. Но у многих пресноводных популяций количество костных пластинок сократилось до 0–9. Селективное преимущество от сокращения числа пластинок для пресноводных рыб, по‑видимому, объясняется большей гибкостью тела и повышенной маневренностью.
Ученые следили за эволюцией колюшки и в режиме реального времени. Озеро Лоуберг на Аляске было заселено морской колюшкой после проведения в 1982 г. программы химического уничтожения местной популяции колюшек. Регулярный анализ, проводившийся с 1990 по 2001 г., показал, что всего за 12 лет доля океанической формы колюшки сократилась со 100 до 11 %, а доля форм с малым количеством костных пластинок достигла 75 %; еще одну небольшую часть составляют различные промежуточные формы (рис. 2.5). Эволюция колюшки в озере Лоуберг показывает, что эта рыба способна адаптироваться к жизни в пресной воде всего за десять лет после переселения из океана. Поскольку колюшка так быстро адаптируется к новым условиям обитания, она является прекрасной моделью для изучения эволюции, и мы вновь обратимся к ней в восьмой главе.

Рис. 2.5. Быстрая эволюция костных пластинок у трехиглой колюшки. В североамериканских озерах океаническая колюшка адаптировалась к новым условиям жизни путем сокращения числа и размера защитных костных пластинок на боках. Из статьи M. A. Belletal., 2004, Evolution 58:814.
Приведенные примеры иллюстрируют быстроту эволюции и силу естественного отбора в отношении уже существующих в популяции вариантов. Но откуда берутся эти варианты? И что происходит, если нужные варианты отсутствуют? Как долго популяция может «ждать» появления новых вариантов?
Дата добавления: 2018-09-22; просмотров: 325; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!