Дифференцировочные антигены Т-лимфоцитов.
Для клинической практики большое значение имеет определение разных маркеров лимфоцитов. Основная концепция дифференцировки лейкоцитов основана на существовании специфических мембранных рецепторов.
Так как такие рецепторные молекулы могут выступать в роли антигенов, существует возможность их выявления с помощью специфических антител, которые реагируют лишь с одним антигеном клеточной мембраны. В настоящее время существует огромное количество видов моноклональных антител к дифференцировочным антигенам лейкоцитов человека.
В связи с их важностью и для улучшения диагностики необходимы стандартизация специфичностей дифференцировочных антигенов.
В 1986 году предложена номенклатура дифференцировочных антигенов лейкоцитов человека. Это СД-номенклатура (cluster of differentiation – кластер дифференцировки). Она базируется на способности моноклональных антител реагировать с определенными дифференцировочными антигенами. СД-группы нумеруются.
На сегодняшний день имеются моноклональные антитела к целому ряду дифференцировочных антигенов Т-лимфоцитов человека.
При определении общей популяции Т-клеток используются моноклональные антитела специфичности СД2, 3, 5, 6 и 7.
СД2. моноклональные антитела специфичности СД2 направлены против антигена, который идентичен “рецептору эритроцитов барана”. Способность Т-лимфоцитов образовывать розетки с эритроцитами брана обеспечивает простую и надежную идентификацию этих клеток. СД2 обнаруживается на всех зрелых периферических Т-лимфоцитах, на большинстве тромбоцитов, а также на определенных популяциях клеток – О-лимфоцитов (ни Т- ни В-лимфоциты).
|
|
|
СД3. моноклональные антитела этого класса реагируют с тримолекулярным белковым комплексом, который ассоциирован с антигенспецифическим рецептором Т-клетки, являющийся основным функциональным маркером этой популяции. СД3 используется для идентификации зрелых Т-клеток.
СД5. антиген представляет собой гликопротеин, выявляемый на всех зрелых Т-клетках. Определяется на поздних стадиях дифференцировки клеток в тимусе. Часто маркер выявляется на клетках больных с В-клеточным типом хронического лимфолейкоза.
СД6. антитела специфичности СД6 реагируют с высокомолекулярным гликопротеином, присутствующим на мембране всех зрелых Т-клеток. Антиген выявляется также на небольшой части периферических В-клеток и присутствует у большинства лейкозных клеток В-клеточного типа хронического лимфолейкоза.
СД7. выявляется у 85 % зрелых Т-клеток. Присутствует также и на тимоцитах. Он считается наиболее надежным критерием диагностики острых Т-клеточных лейкозов.
|
|
|
Помимо этих основных Т-клеточных маркеров известны и другие дифференцировочные антигены Т-клеток, которые характерны либо для определенных стадий онтогенеза, либо для различающихся по функциям субпопуляций. Среди них наиболее широко распространены СД4 и СД8.
СД4. зрелые СД4+ Т-клетки включают Т-лимфоциты, обладающие хелперной активностью и индукторы. Особое значение имеет то, что СД4 связывается с вирусом СПИДа, что приводит к проникновению вируса внутрь клеток этой субпопуляции.
СД8. Субпопуляция СД8+ Т-клеток включает цитотоксические и супрессорные Т-лимфоциты.
Виды антигенной специфичности.
Специфичностью называют способность антигена избирательно реагировать со строго определенными антителами или клонами лимфоцитов. Специфичность зависит от наличия в структуре молекулы вещества тех или иных антигенных детерминант.
К специфичности АГ относят динамику поступления в организм и его выведения.
На иммунный ответ влияет количество поступающего антигена: чем его больше, тем более выражен иммунный ответ. Однако передозировка АГ вызывает обратную реакцию - иммунологическую толерантность.
По специфичности АГ подразделяют:
|
|
|
видовые - у всех особей вида;
типовые - вариантные;
групповые - общие для разных видов и родов;
стадийные - появляются на определенных стадиях развития.
Аутоантигены - антигены собственного организма. В нормальных условиях они не вызывают реакции иммунной системы вследствие либо иммунологической толерантности, либо недоступности для контакта с факторами иммунитета - это так называемые забарьерные антигены. При срыве толерантности или нарушении целостности биологических барьеров компоненты иммунной системы начинают специфически реагировать на аутоантигены выработкой специфических факторов иммунитета.
Ксеногенные антигены - общие для организма, стоящих на разных ступенях биологической классификации, например, относящихся к разным родам и видам - это обнаружил Д. Форсман (1911). Ученый иммунизировал кролика суспензией органов морской свинки. Оказалось, что полученная в ходе эксперимента иммунная сыворотка была способна не только взаимодействовать с антигенами морской свинки, но и аглютинировать эритроциты барана. Позже было установлено, что морская свинка и баран имеют ряд структурно сходных детерминант, дающих перекрестное реагирование. Эти антигены получили обобщенное название "антигены Форсмана".
|
|
|
"Аллогенные антигены" - (комплексные, групповые) - общие для генетически не родственных организмов, относящихся к одному виду. По ним из общей популяции можно выделить отдельные группы организмов. Примером таких антигенов у людей являются антигены группы крови, раковоэмбриональные антигены и др. Аллогенные ткани при трансплантации иммунологически не совместимы - они отторгаются микроорганизмом.
Изоантигены - антигены, общие только для однояйцевых близнецов, животных инбредных линий. Изотрансплантаты обладают практически полной иммунологической совместимостью и не отторгаются при пересадке.
Изоантигены являются мукополисахаридными антигенами - это единственная группа антигенов, против которых постоянно имеются натуральные антитела (агглютинины). По антигенам к А- и В-изоантигенам определяются четыре основные группы крови.
Антигены гистосовместимости.
В настоящее время признана и доказана концепция наличия у человека гена иммунного ответа (JR-гена), который детерминирует силу иммунного ответа на антиген. Установлено, что ген иммунного ответа связан HLA - комплексом и может быть картирован на интервале В-D, в непосредственный близи к сублокусу HLA-D. Из того, что различные группы болезней связаны не с одним, а с несколькими аллелями HLA-B и HLA-D, можно предположить, что JR-система является многоаллельной.
Иллюстративный материал
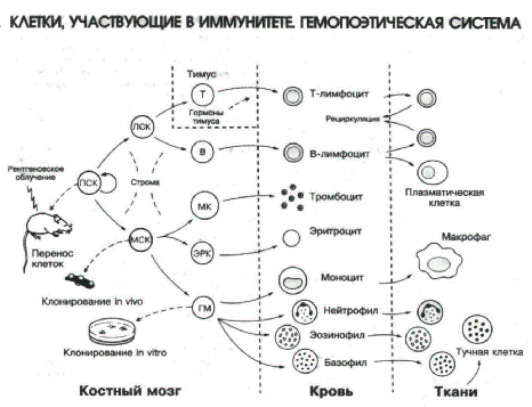
Слайд-рисунок 1.
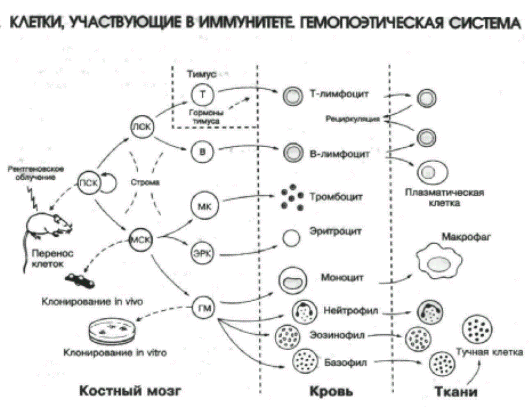
Слайд-рисунок 2.
КОМПОНЕНТЫ ИММУННОЙ СИСТЕМЫ
2.1. Лимфоидная ткань
Анатомический синоним иммунной системы — лимфоидная система. Однако понять устройство и функционирование иммунной системы можно, только проследив конкретные взаимосвязи лимфоидной системы с другими системами организма, по крайней мере с системой клеток крови и кровеносных сосудов, а также покровными тканями (слизистыми оболочками и кожей). Эти системы — ближайшие партнёры, на которые в своей работе опирается система лимфоцитарного иммунитета. Более того, гистогенетически собственно лимфоциты — компонент системы крови: лимфоциты происходят из общей для всех клеток крови стволовой кроветворной клетки. Как мы увидим дальше, нельзя избежать рассмотрения иммунных подсистем барьерных тканей — кожи и слизистых оболочек, печени, что и понятно, учитывая такие функции иммунитета, как защиту от инфекций и реакции на пищевые и ингаляционные внешние вещества.
В организме взрослого здорового человека содержится около 1013 лимфоцитов, т.е. примерно каждая 10-я клетка тела — лимфоцит. Как они расположены в организме? Анатомо-физиологический принцип устройства иммунной системы — органно-циркуляторный. Это значит, что есть ряд специализированных органов с организованной внутренней структурой. При этом лимфоциты не «сидят» в лимфоидных органах постоянно (в отличие от, например, гепатоцитов в печени), а интенсивно рециркулируют между лимфоидными органами и нелимфоидными тканями через лимфатические сосуды и кровь: через один лимфатический узел за один час проходит »109 лимфоцитов. Из общего количества лимфоцитов организма в каждый момент времени в крови находится только 0,2–2%. Миграция лимфоцитов из крови в ткани и из тканей в кровь происходит сквозь стенку сосудов за исключением эфферентных лимфатических сосудов лимфоидных органов, и механизм этой миграции включает в себя специфические взаимодействия определённых молекул на мембране лимфоцита с определёнными молекулами на мембране клеток эндотелия стенки сосудов (такие молекулы называют адгезинами, селектинами, интегринами, хоминг–Рц (от англ. home — «дом» в смысле место, предназначенное данному лимфоциту природой). Эти взаимодействия происходят не в каком придётся месте сосуда, а в определённых местах, например в лимфатических узлах — это эндотелий посткапиллярных венул. Процесс миграции лимфоцитов, конечно же, не носит характер случайного передвижения, а строго регулируется рядом факторов, зависящих от местных тканевых и системных физиологических «задач» организма.
à Кроветворный костный мозг — место обитания стволовых кроветворных клеток (СКК).
à Инкапсулированные органы: тимус, селезёнка, лимфатические узлы, печень.
à Неинкапсулированная лимфоидная ткань.
Ä Лимфоидная ткань слизистых оболочек (MALT — Mucosal–Аssociated Lymphoid Tissue). В том числе:
Ú Лимфоидная ткань, ассоциированная с ЖКТ (GALT — Gut–Associated Lymphoid Tissue) — миндалины, аппендикс, пейеровы бляшки, а также субпопуляция внутриэпителиальных лимфоцитов слизистой оболочки ЖКТ.
Ú Лимфоидная ткань, ассоциированная с бронхами и бронхиолами (BALT — Bronchus–Associated Lymphoid Tissue), а также внутриэпителиальные лимфоциты слизистой оболочки дыхательной системы.
Ú Лимфоидная ткань, ассоциированная с женскими половыми путями (VALT — Vulvovaginal–Associated Lymphoid Tissue), а также внутриэпителиальные лимфоциты их слизистой оболочки.
Ú Лимфоидная ткань, ассоциированная с носоглоткой (NALT — Nose–Associated Lymphoid Tissue), а также внутриэпителиальные лимфоциты её слизистой оболочки.
Ä Субпопуляции лимфоцитов печени, которая в качестве лимфоидного барьера «обслуживает» кровь воротной вены, несущей все всосавшиеся в кишечнике вещества.
Ä Лимфоидная подсистема кожи (SALT – Skin-Associated Lymphoid Tissue) диссеминированные внутриэпителиальные лимфоциты и региональные лимфатические узлы и сосуды лимфодренажа.
à Периферическая кровь — транспортно–коммуникационный компонент иммунной системы.
Кроветворный костный мозг и тимус называют центральными органами иммунной системы потому, что на их территории происходит дифференцировка лимфоцитов из стволовой кроветворной клетки, так называемый лимфопоэз. Лимфопоэз — это дифференцировка лимфоцитов от стволовой кроветворной клетки до зрелого неиммунного лимфоцита. Зрелые неиммунные лимфоциты локализуются в периферических лимфоидных органах и циркулируют между ними через кровь. На территории периферических лимфоидных органов зрелые неиммунные лимфоциты вступают в контакты с АПК (АПК). Если антигенраспознающий Рц лимфоцита связывает комплементарный Аг на территории периферических лимфоидных органов, где в норме создаются все необходимые условия (корецепторные взаимодействия) для начала развития иммунного ответа, то лимфоцит вступает на путь додифференцировки в режиме иммунного ответа, т.е. начинает пролиферироватъ и продуцировать эффекторные молекулы (цитокины, перфорин, гранзимы и др. в зависимости от субпопуляции лимфоцита). Дифференцировку лимфоцитов на периферии после распознавания Аг называют иммуногенезом. Зрелые неиммунные лимфоциты в англоязычной литературе называют naive (наивные),или virgine (девственные).
| Схема кроветворения | |||||||||||||||||||||||||
| Стволовая кроветворная клетка, ростки | |||||||||||||||||||||||||
| Эрит-роид-ный | Миелоидный | Мега-карио-цитарный | Клетка-редшественник B1–лимфоцитов (автономизируется в эмбриональном периоде) | Общая лимфоидная клетка–предшественница | |||||||||||||||||||||
|
| Коммитированные клетки–предшественницы для дифференцировки в: | ||||||||||||||||||||||||
| Эритробласты, незрелые эритроциты | Общая клетка–предшественница фагоцитов — нейтрофилов и моноцитов | Общая клетка–предшественница эозинофилов и базофилов | Клетка-предшест–венница тучных клеток | Мега-карио-бласты | — | T | В2 | ДК2 | NK | ||||||||||||||||
|
|
|
| T | T |
|
|
| ||||||||||||||||||
| Зрелые клетки в периферической крови | |||||||||||||||||||||||||
| Эритроциты | Нейтро-филы | Моноциты | Эози-но-филы | Базо-филы | ? | Тромбоциты | В1 –лимфоциты (мало) | T | T | В2 | ДК 2–лимфоидные дендритные клетки | субпопуляция NK крови | |||||||||||||
| Оседлые потомки в тканях (в норме, вне воспаления) | |||||||||||||||||||||||||
| — | — | Макрофаги | DC1-миелоидные дендритные клетки | — | — | Тучные клетки | — | В1–лимфоциты в брюшной и плевральной полостях | T | T | Плазмоциты (в лимфоузлах, костном мозге, слизистых) | ДК 2 (интердигитирующие клетки в тимусе, в лимфоузлах; клетки Лангерганса в коже) | NK тканевые (в печени, в деци-дуальной оболочке матки) | ||||||||||||
| Схема лимфопоэза | |||||||||||||||||||||||||
| Стволовая кроветворная клетка (костный мозг)
| |||||||||||||||||||||||||
| Коммитированная к лимфопоэзу клетка–предшественница (общая для всех лимфоцитов)
| |||||||||||||||||||||||||
| 1 ДК 2 крови, стромы тимуса, клетки Лангерганса | 2 В2 | 3 T–лимфоциты
| 4 Нормальные киллеры
| ||||||||||||||||||||||
| АПК ДК 2 | В2 (в костном мозге) | T | T |
| |||||||||||||||||||||
|
|
| CD4+ | CD8+ | CD4– CD8– | ЦТЛ | Th(?) |
| ||||||||||||||||||
| Функционально зрелые субпопуляции иммунных лимфоцитов и дендритные клетки | |||||||||||||||||||||||||
| Активированные АПК | Продуценты IgM IgG IgA IgE | NK1.1 + Th0 Тh1 Th2 (Th3) тчцтл | ЦТЛ | Ранние продуценты ИЛ–4 | ЦТЛ; [Th0, Тh1, Th2, (Тh3)] (?) | Клетки–эффекторы IgG–опосредованной АЗКЦТ | |||||||||||||||||||
|
|
|
|
| ||||||||||||||||||||||
Обязательным процессом в начале иммуногенеза в периферических лимфоидных органах является пролиферация клонов лимфоцитов, распознавших Аг. В результате иммуногенеза развиваются клоны иммунных или эффекторных лимфоцитов, которые в англоязычной литературе называют armed (вооружённые) или effector (эффекторные) лимфоциты. Иммунные лимфоциты распознают Аг и организуют деструкцию различных периферических тканей организма, где этот Аг присутствует.
В костном мозге проходит дифференцировка всех лейкоцитов крови. По выходе из костного мозга в периферические ткани лейкоциты в норме уже никогда не будут пролиферировать (в отличие от лимфоцитов). В ответ на адекватные сигналы извне они лишь будут активированы к выполнению своих предопределённых в процессе дифференцировки функций.
Строма костного мозга поддерживает пролиферацию и дифференцировку эритроидного (в итоге эритроциты), миелоидного (лейкоциты) и мегакариоцитарного ростков кроветворения. На территории костного мозга из стволовой кроветворной клетки образуется общая клетка — предшественник всех лимфоцитов, из которой также на территории костного мозга проходят поэз 3 из 4 её потомков: В2–лимфоциты, нормальные киллеры (NK) и дендритные клетки (ДК). Четвертый потомок, коммитированный (запрограммированный) к дифференцировке в T–лимфоциты, мигрирует для прохождения поэза из костного мозга в тимус, и какая-то часть — в слизистые оболочки, в основном ЖКТ. Схема кроветворения и лимфопоэза представлена в табл. 2.1.
Из табл. 2.1 видно, какие клетки входят в состав иммунной системы. Собственно иммуноциты — это все варианты лимфоцитов — T, B, NK. Непосредственные клетки — сотрудники лимфоцитов — все варианты лейкоцитов —·дендритные клетки, нейтрофилы, моноциты/макрофаги, эозинофилы, базофилы, тучные клетки. И даже эритроциты вносят свой вклад в деструктивное завершение иммунного ответа — транспортируют иммунные комплексы «Аг+АТ+комплемент» (на эритроцитах есть Рц для комплемента) в печень и селёзенку для фагоцитоза и разрушения.
Кроме названных клеток мезенхимного происхождения, в состав лимфоидных органов входят клетки стромы, это преимущественно эпителиальные клетки эктодермального и энтодермального происхождения, а также эндотелий сосудов, с которым взаимодействуют все клетки — участники иммунного ответа как целого и сквозь который происходит экстравазация лимфоцитов и лейкоцитов при их миграциях.
Таблица 2.1. Схема кроветворения из стволовой кроветворной клетки и схема лимфопоэза
Помимо клеток, «иммунологическая материя» представлена растворимыми молекулами — гуморальными факторами. Это продукты B–лимфоцитов — АТ (они же иммуноглобулины) и растворимые медиаторы межклеточных взаимодействий — цитокины. Цитокины — это та молекулярная «материя», посредством которой лимфоцитарный иммунитет «встроен», интегрирован в организм в целом. Цитокины вырождены относительно тканевой дифференцировки клеток–продуцентов, т.е. одни и те же гены цитокинов могут экспрессироваться в клетках разной дифференцировки. Цитокины (от греческих корней cyto — клетка, kinos — движение) — молекулы, секретируемые клетками во внеклеточную среду с целью воздействовать на другие клетки или на себя же, подать сигнал к запуску тех или иных процессов в клетках–мишенях. Цитокины — молекулярный «язык» межклеточного общения, для большинства цитокинов — локального, близкодействующего взаимодействия, ещё один «язык» наряду с нейромедиаторами и эндокринными гормонами (последние два — дистантного действия).
Кроветворение — дифференцировка клеток крови, или гемопоэз, в раннем эмбриогенезе млекопитающих начинается в кровяных островках мезодермы, затем «перемещается» в печень, селёзенку и костный мозг эмбриона. После рождения гемопоэз поддерживается в течение всей жизни в костном мозге плоских костей — грудине, ребрах, крыльях подвздошной кости, костях черепа и в позвонках. Все клетки крови происходят из общей клетки–предшественницы — стволовой кроветворной клетки (СКК). Относительно параметров состояния пула СКК есть две гипотезы. Строгие доказательства правильности какой–либо одной из них отсутствуют, и получить их, очевидно, не просто. Одна из
гипотез подразумевает, что популяция стволовых клеток делится в течение всей жизни. Вторая гипотеза предполагает, что у взрослых организмов пул истинно стволовых клеток не пополняется за счёт их пролиферации, а только расходуется в течение всей жизни на гемопоэз. Пролиферируют уже коммитированные потомки СКК на определённых этапах своей дифференцировки. Если принять вторую гипотезу, то можно сделать один рабочий вывод: следует быть осторожным с терапевтическими вмешательствами, способными несоразмерно стимулировать гемопоэз или отдельные его ростки (они, кстати, регулируются независимо друг от друга, кроме точек ветвления на альтернативную дифференцировку, например, нейтрофилы и моноциты, эозинофилы и базофилы), чтобы не израсходовать досрочно «золотой запас» СКК.
На территории костного мозга проходит полный «курс» эритропоэза (заканчивается эритроцитами), миелопоэза (заканчивается нейтрофилами, моноцитами, эозинофилами, базофилами), мегакариоцитопоэза (заканчивается тромбоцитами), дифференцировки дендритных клеток и, вероятно, NK, а также лимфопоэза В2–лимфоцитов («классических», в смысле «давно известных»). Для дефинитивных потомков В2–лимфоцитов — плазматических клеток костный мозг является и «периферическим» лимфоидным органом. Значительная часть В2–лимфоцитов, пройдя «курс» иммуногенеза в периферических лимфоидных органах (лимфатических узлах, селёзенке) и превратившись в плазмоциты, возвращается на территорию костного мозга, и там плазмоцит вырабатывает свой ресурс по продукции больших количеств АТ в течение периода от нескольких дней до месяца.
Клетки–предшественницы второй субпопуляции B–лимфоцитов — В1 ещё в периоде эмбриогенеза отселяются из костного мозга в брюшную и плевральную полости и там поддерживают дифференцировку этих «полостных» В1–лимфоцитов в течение всей жизни уже автономно от СКК костного мозга. Если по какой-то причине предшественники В1–лимфоцитов повреждаются, то регенерация популяции В1–лимфоцитов не поддерживается за счёт СКК костного мозга.
Другие ветви дифференцировки общей лимфоидной клетки–предшественницы, а именно предшественники T–лимфоцитов, для прохождения своего лимфопоэза выселяются из костного мозга в другие органы и ткани (тимус и слизистую оболочку ЖКТ).
2.2. Тимус
Тимус (thymus) — специализированный лимфоидный орган, в котором проходит лимфопоэз бoльшая часть T–лимфоцитов организма (аббревиатура «Т» происходит от «Thymus»). Тимус расположен в переднем верхнем средостении, за грудиной, над сердцем. Тимус состоит из двух больших долей, которые фрагментированы на множество долек, разделённых фиброзными перегородками. Эти дольки и являются структурными единицами строения тимуса (рис. 2.2). В каждой дольке чётко различимы две гистологические зоны: по периферии — корковая, в центре — мозговая. Строма тимуса эпителиальная. Полагают, что эпителий разных его зон происходит из различных зародышевых листков. Эпителий корковой зоны эктодермального происхождения, в эмбриогенезе развивается из 3–го и 4–го глоточных выпячиваний эктодермы. Эпителий мозговой зоны энтодермального происхождения и закладывается из 3–го и 4–го глоточных карманов.
Есть в тимусе и такие необычные клетки, как миоидные . Они происходят, вероятно, из нервного гребня. В норме их немного. Эти клетки содержат белки, характерные для мышечных клеток (актин, миозин и др.). У мышей морфогенез тимуса различимо проходит с 9-х по 11-е сутки эмбрионального развития.
Эпителиальные клетки тимуса имеют особую морфологию. Эпителиальные клетки коры своими отростками «обнимают и баюкают» лимфоциты тимуса (тимоциты), поэтому они названы nurse cells (клетки–"сиделки», «медсёстры», «нянечки»).
Эпителиальные клетки тимуса продуцируют цитокины ИЛ–1, ИЛ–3, ИЛ–6, ИЛ–7, LIF, GM–CSF. На клетках эпителия тимуса экспрессированы такие молекулы адгезии, как LFA–3 и ICAM–1, которые комплементарны молекулам адгезии на тимоцитах — соответственно CD2 и LFA–1. Эти взаимодействия и удерживают развивающиеся тимоциты на территории тимуса на необходимое для дифференцировки время.
Клетки мезенхимного, или костномозгового происхождения в тимусе представлены тимоцитами (это лимфоциты тимуса), а также дендритными клетками тимуса и макрофагами. Дендритные клетки расположены преимущественно в зоне, переходной между корковой и мозговой. Макрофаги присутствуют в корковой зоне, пограничной и мозговой.
Тимоциты дифференцируются из общей стволовой кроветворной клетки. На клетках–предшественницах тимоцитов ещё вне тимуса у человека экспрессированы известные молекулы клеточных мембран CD7, CD2, CD34 и цитоплазматическая форма CD3. У мыши на претимоцитах обнаружены маркёры Thy–1, HSA, Pgp–1, H–2, Sca–1 (Ly–6 A/E), немного CD4. Клетки–предшественницы приходят в тимус через стенку венул с высоким эндотелием в корково–мозговой области и оттуда мигрируют в субкапсулярную зону.
У молодой мыши в возрасте 5–8 нед в тимусе содержится »2´108 тимоцитов. При этом ежедневно в результате митозов вновь образуется 5´107 тимоцитов. Но в периферические лимфоидные органы выходит около 106 зрелых неиммунных T–лимфоцитов, т.е. меньше 1% от общего числа тимоцитов. Остальные 99% тимоцитов погибают в тимусе по механизму апоптоза и устраняются фагоцитозом макрофагами. Правда, гистологических признаков столь массовой гибели клеток именно на территории тимуса нет. Поэтому более вероятно, что запрограммированные на гибель тимоциты ещё живыми уходят из тимуса и попадают напрямую в органы, специализированные на катаболизме и выведении из организма, — в печень, селёзенку, кишечник. Там эти клетки и разрушаются. Таковы вкратце закономерности дифференцировки T–лимфоцитов.
В мозговой зоне долек имеются плотные образования из скрученных эпителиальных клеток — тельца Гассаля (тельца вилочковой железы). Вероятно, это места компактного скопления дегенерирующих эпителиальных клеток.
Тимус интенсивно васкуляризирован. Стенки капилляров и венул — это гематотимический барьер на входе в тимус и, возможно, на выходе из него. Выход зрелых лимфоцитов из тимуса либо свободен (каждая долька имеет эфферентный лимфатический сосуд, выносящий лимфу в лимфатические узлы средостения, оттуда в грудной лимфатический проток и через него в системную циркуляцию), либо происходит путём экстравазации через стенку посткапиллярных венул с высоким эндотелием в корково–мозговой области и/или через стенку капилляров.
От других лимфоидных органов тимус отличает особая постнатальная динамика его морфогенеза в зависимости от возраста. К моменту рождения тимус полностью сформирован. Он густо заселен лимфоцитами (тимоцитами) в течение всего детства и до момента полового созревания. После пубертатного периода тимус начинает уменьшаться в размерах, сморщиваться. Удивительно, но тимэктомия у взрослых не приводит к серьёзным дефектам в иммунитете, как если бы в детстве и подростковом возрасте был создан необходимый и достаточный пул периферических T–лимфоцитов на всю оставшуюся жизнь. Казалось бы, анатомия тимуса и его возрастная инволюция давно известны, но где хранится, как расходуется и как регенерирует множество T–лимфоцитов на протяжении жизни человека — это некоторая почти разгаданная загадка.
2.3. Лимфатические узлы
Лимфатические узлы — множественные, симметрично расположенные, инкапсулированные периферические лимфоидные органы бобовидной формы, размером от 0,5 до 1,5 см в длину (вне воспаления). Лимфатические узлы через афферентные лимфатические сосуды (которых несколько на один узел) дренируют тканевую жидкость. Лимфатические узлы расположены регионарно и называются в соответствии с частью тела, которую они «обслуживают»: околоушные, заднешейные, подмышечные, подколенные, паховые, брыжеечные и т.д. Таким образом, лимфатические узлы — это «таможня» для всех веществ (в том числе Аг), попадающих во внутреннюю среду организма.
Афферентные лимфатические сосуды впадают в субкапсулярный синус лимфатического узла. Из анатомических ворот узла параллельно с артерией и веной выходит единственный эфферентный сосуд, несущий лимфу в конечном итоге в грудной лимфатический проток, который впадает в нижнюю полую вену и таким образом лимфа вливается в системный кровоток.
Лимфатический узел состоит из коркового и мозгового вещества.
B–клеточная зона. Корковое вещество лимфатического узларазделено соединительнотканными перегородками на радиальные секторы и содержит лимфоидные фолликулы, B–лимфоцитарная зона. Строма фолликулов содержит фолликулярные дендритные клетки (ФДК), формирующие то особое микроокружение, в котором происходит уникальный для B–лимфоцитов процесс соматического гипермутагенеза вариабельных сегментов генов иммуноглобулинов и отбор наиболее аффинных вариантов АТ («созревание аффинности АТ»). Лимфоидные фолликулы проходят 3 стадии развития. Первичный фолликул — мелкий фолликул, состоящий из неиммунных B–лимфоцитов. После того как B–лимфоцит распознаёт Аг и получит все необходимые костимулирующие сигналы, он вступит в иммуногенез, строго необходимым этапом которого является пролиферация клона B–лимфоцитов. Фолликул, содержащий интенсивно пролиферирующие B–лимфоциты, называют герминативным центром (центр размножения). Первичный фолликул преобразуется в активный, т.е. содержащий центр размножения в течение примерно 1 нед после активной иммунизации. По завершении процесса иммуногенеза лимфоидный фолликул существенно уменьшается в размере, это вторичный фолликул.
T –клеточная зона. В паракортикалъной зоне лимфатического узла расположены T–лимфоциты и посткапиллярные венулы, через стенку которых происходит миграция лимфоцитов из крови в лимфатический узел. Это T–зависимая зона. В активном состоянии посткапиллярные венулы имеют не плоский, а высокий эндотелий. В T–зависимой зоне содержится много интердигитирующих дендритных клеток. Это совсем другие дендритные клетки, чем фолликулярные дендритные клетки.
Интердигиталъные дендритные клетки — клетки костномозгового происхождения, мигрировавшие в узел с тканевой жидкостью по афферентным лимфатическим сосудам из покровных тканей (в коже эти клетки называют клетками Лангерганса, или белыми отростчатыми эпидермоцитами), это АПК для T–лимфоцитов.
Под паракортикальной зоной расположены мозговые тяжи, в которых много макрофагов, и, если узел находится в состоянии активного иммунного ответа, в этих тяжах можно видеть зрелые B–лимфоциты (плазматические клетки). Тяжи впадают в синус мозгового вещества, из которого выходит эфферентный лимфатический сосуд.
2.4. Селезёнка
Селезёнка — относительно большой непарный орган, с массой в среднем 150 г у взрослого человека. Лимфоидную ткань селезёнки называют белой пульпой. Селезёнка — лимфоцитарная «таможня» для Аг, попавших в системную циркуляцию. Лимфоциты селезёнки локализованы вокруг артериол в виде так называемых периартериолярных муфт.
2.5. Печень
О том, что печень имеет уникальные функции в иммунной системе организма, свидетельствуют следующие факты.
Ú В эмбриональном периоде развития печень является мощным органом кроветворения и лимфопоэза; есть или нет хоть в какой-то мере лимфопоэз в печени у взрослых — вопрос открытый, и возможно что есть.
Ú Аллогенные трансплантаты печени не отторгаются или почти не отторгаются (по крайней мере не требуют столь жёсткой иммуносупрессивной химиотерапии, как трансплантаты других органов).
Ú Толерантность на вводимые перорально Аг можно индуцировать только при нормальном физиологическом кровоснабжении печени и не удаётся индуцировать после хирургической операции по созданию портокавальных анастомозов.
Ú Печень синтезирует белки острой фазы — C–реактивный белок и связывающий маннозу лектин, которые являются растворимыми Рц, непосредственно связывающими микробные продукты.
Ú Печень синтезирует белки системы комплемента, в том числе C1. C1q тоже непосредственно связывает бактериальные продукты и инициирует активацию всей системы комплемента. Без активации системы комплемента невозможен адекватный гуморальный иммунный ответ в виде синтеза АТ в B–лимфоцитах.
Ú В печени содержатся собственные субпопуляции лимфоцитов, в том числе уникальные лимфоциты, сочетающие в себе признаки T– и NK–лимфоцитов.
Клеточный составпечени известен из гистологии.
1. Паренхима печени — гепатоциты — крупные полиморфные клетки (30 мкм в диаметре), у взрослых 25% гепатоцитов тетраплоидны. На гепатоцитах содержится очень мало молекул MHC класса I и в здоровой печени нет молекул MHC II, которые, однако, могут экспрессироваться при заболеваниях этого органа.
2. Купферовские клетки — макрофаги печени. Они составляют около 15% от общего числа клеток печени и 80% всех макрофагов тела (по приблизительной оценке). Плотность макрофагов выше в перипортальных областях печени, чем в среднем по органу.
3. Эндотелиальные клетки синусоидов печени особенные — под ними нет базальной мембраны.
4. Звездчатые клетки, или клетки Ито, — уникальные клетки в пространствах Диссе (перисинусоидальные пространства). Эти клетки ещё называют липоцитами, так как они содержат жировые вакуоли с витамином А. Но они содержат также контрактильные белки — a-актин и десмин, характерные для гладкомышечных клеток, — и могут трансформироваться в миофибробласты. Клетки Ito также являются основными синтезаторами коллагена в печени.
5. Лимфоидная система печени, помимо лимфоцитов, содержит ещё и анатомический субстрат циркуляции лимфы — это пространства Диссе. Эти пространства непосредственно открыты для содержимого портального тракта с одной стороны и также без всяких барьеров для ткани печени с другой стороны. Лимфоток в печени составляет, по-видимому, не менее 15–20% всего лимфотока в организме (т.е. больше, чем в любом другом органе тела).
Достоверно подсчитать количество лимфоцитов в печени — за пределами возможностей существующих методов, так как при выделении лимфоцитов в суспензию потери велики и не контролируемы. Работ по подсчёту субпопуляций лимфоцитов в печени крайне мало. Соответственно результаты практически не проверяемы. Но те, что есть, показывают следующее распределение (по данным проточной цитофлюорометрии):
а) на 1 г ткани печени приходится 2–15´106 лимфоцитов;
б) внутрипечёночные лимфоциты составляют примерно 25% непаренхиматозных клеток печени (купфферовские клетки тоже 25%);
в) лимфоциты NK составляют 20% всех лимфоцитов;
г) лимфоциты Tab составляют 20% всех лимфоцитов, из них CD8+ в 5–2 раза больше, чем CD4+ (соотношение CD8:CD4 = 4:1, по данным другой работы — 2:1), в любом случае в периферической крови наоборот — CD4:CD8=2:1.
д) лимфоциты Tgd составляют около 3% всех лимфоцитов;
е) B–лимфоцитов не более 5%, на большинстве из них экспрессированы IgA.
Есть обоснованное предположение, что лимфоциты печени обеспечивают постоянное поддержание иммунологической толерантности к пищевым веществам. Хотя, конечно, в какой-то мере иммунный ответ на пероральные вещества может развиваться, но он более выражен при патологии ЖКТ, или/и при аллергических заболеваниях, нежели в норме.
2.6. Лимфоидная ткань слизистых оболочек и кожи
Неинкапсулированная лимфоидная ткань слизистых оболочек — представлена глоточным лимфоидным кольцом Пирогова, пейеровыми бляшками тонкой кишки, лимфоидными фолликулами аппендикса, а также лимфоидной тканью слизистых оболочек желудка и кишечника, бронхов и бронхиол, слизистых оболочек мочеполовой системы и всех остальных слизистых оболочек.
Пейеровы бляшки (по новой номенклатуре — групповые лимфатические фолликулы) располагаются в l . propria тонкой кишки. Каждая бляшка примыкает к эпителию кишки под так называемыми М–клетками («М» от membranous, эти клетки в отличие от энтероцитов не имеют ворсинок), являющимися «входными воротами» в пейерову бляшку. Основная масса лимфоцитов бляшки — это B–клеточный фолликул с зародышевым центром посередине. T–клеточные зоны окружают фолликул ближе к слою энтероцитов. B–лимфоциты составляют 50–70%, T–лимфоциты — 10–30% всех клеток пейеровой бляшки. Основная функциональная нагрузка пейеровых бляшек — поддержание иммуногенеза B–лимфоцитов и их дифференцировка в плазмоциты, продуцирующие АТ — иммуноглобулины секреторных классов А и Е (IgA и IgE). Продукция IgA в слизистой оболочке кишки составляет более 70% общей ежедневной продукции иммуноглобулинов в организме: у взрослого человека около 3 г IgA каждый день. Более 90% всего синтезируемого организмом IgE, оказывается, тоже экскретируется через слизистую оболочку кишки.
Организованная лимфоидная ткань слизистых оболочек, пусть даже и без капсул, это не единственное место пребывания лимфоцитов в слизистых оболочках. Есть ещё (и немало) лимфоциты, диссеминированные по одному среди эпителиальных клеток. Их назвали внутриэпителиальными лимфоцитами. Это T–лимфоциты. На их поверхности экспрессирована особая молекула, обеспечивающая адгезию этих лимфоцитов к энтероцитам — HML–1. У мышей большинство, а у человека не менее 10–50% внутриэпителиальных лимфоцитов составляют Tgd/CD8aa+.
В l . propria присутствует много диссеминированных активированных (иммунных) CD4+ T–лимфоцитов, мигрировавших сюда из мезентериальных лимфатических узлов для осуществления эффекторных реакций. Здесь много и иммунных B–лимфоцитов, и плазматических клеток, мигрировавших сюда из пейеровых бляшек и мезентериальных лимфатических узлов для осуществления массовой продукции секреторных иммуноглобулинов.
Таким образом, вполне оправдано понятие тканевой, местной иммунной подсистемы слизистых оболочек, так же, как и печени, и кожи. В каждой ткани популяции лимфоцитов имеют свои особенности. Более того, чтобы лимфоциту мигрировать в определённую ткань, ему необходимо экспрессировать на мембране определённый так называемый хоминг –Рц (от англ. home — дом, место «прописки» лимфоцита). Для внутриэпителиальных лимфоцитов слизистой оболочки кишки это HML–1, для внутриэпителиальных лимфоцитов кожи это CLA–1. Чтобы зрелому неиммунному T–лимфоциту, вышедшему из тимуса, попасть на территорию T–зависимой зоны лимфатического узла, ему необходимо экспрессировать на своей поверхности особую молекулу — L–селектин. Это важно понимать и в теоретическом аспекте, и в прикладном. В рутинных клинико–иммунологических анализах в качестве биологического материала используют периферическую кровь из вены. Так вот, если патологический процесс затронул тканевые лимфоциты и их клетки-партнёры (эндотелий, макрофаги, другие лейкоциты), то лимфоциты, циркулирующие в крови, лишь в минимальной мере могут нести на себе (или не нести совсем) признаки тканевой патологии, и неадекватность такого анализа на практике часто дискредитирует в целом то, что называют анализом иммунного статуса.
Миграция лимфоцитовмежду лимфоидными органами и не–лимфоидными тканями, несмотря на свою высокую интенсивность, ни в коем случае не напоминает «броуновское движение» — она строго и тонко регулируется экспрессией определённых молекул адгезии и их лигандов. В результате в каждой ткани, в каждом органе имеется свой особый субпопуляционный состав лимфоцитов и их клеток — партнёров по иммунному ответу.
Иллюстративный материал
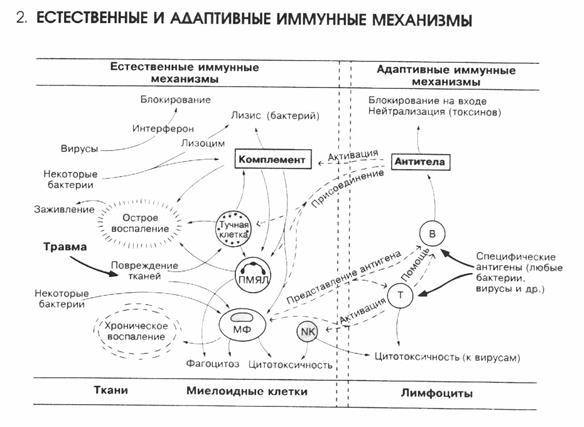
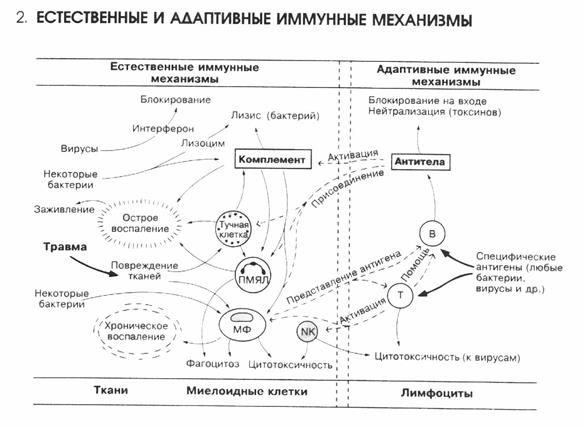
Слайд-рисунок 1.
Слайд-рисунок 2.
Рекомендуемая литература:
На русском языке
Основная:
1.Ройт А. «Иммунология», Москва, 2007
2.Хаитов Р.М. «Иммунология», Москва, 2000
3.Шортанбаев А.А., Кожанова С.В «Общая иммунология», Алматы, 2008
Дополнительная:
1. Федосеев Г.Б. «Аллергология», Санкт-Петербург, 2007
2. Хацкий С.Б. «Аллергология в схемах и таблицах», Санкт-Петербург, 2001
3. Современные аспекты клинической пародонтологии// под ред. Дмитриевой Л.А., Москва, 2001.
4. Грудянов А.И. Пародонтология. Избранные лекции. Москва, 1997
5. Медуницын Н.В. «Вакцинология», Москва, 2004
На казахском языке
основная:
1. Шортанбаев А.А., Кожанова С.В. «Жалпы иммунология», Алматы, 2008
2. К.А. Жуманбаев «Клиникалык иммунология жэне аллергология», Караганды, 2008
дополнительная:
1. Балпанова Г.Т. «Трансплантациялық иммунитет», Алматы, 2002
2. Бижигитова Б.Б. «Комплемент жүйесі мен антигенді таныстырушы жасушалардың иммунды жауаптағы маңызы», Алматы, 2006
3. Мухамбетова С.Г., Каракушикова А.С., Кожанова С.В., Садвакасова Г.С., Балпанова Г.Т. «Иммунды статусқа баға берудің қазіргі әдістері», Алматы, 2005
Контрольные вопросы (обратная связь)
1. Предмет и задачи иммунологии.
2. Основные аспекты фундаментальной иммунологии.
3. Современные определения иммунитета, понятие об иммунной реакции.
Дата добавления: 2018-09-20; просмотров: 469; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!