ГЛАВА 6. ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ (эндокринные железы) 4 страница
ж
Пути рециркуляции лимфоцитов
Рисунок 386. Взаимосвязи органов иммунной системы. Показаны взаимоотношения центральных и периферических органов иммунной системы и направления миграции лимфоцитов в процессе созревания и рециркуляции (указано стрелками). В — В-лимфоциты; М — моноциты; Г — гранулоциты; Т — Т-лимфоциты.
| Ж |
Органы иммунной системы. Хотя клетки, выполняющие иммунологические функции, рассеяны по всему организму, к иммунной системе, строго говоря, относятся лишь лимфоидные органы и лимфоидные скопления, т.е. органы и структурные образования, основу которых составляют лимфоциты. Лимфоидные органы разделяют на центральные (первичные) и периферические (вторичные). Такими органами являются красный костный мозг, тимус, лимфатические узлы, селезенка и кишечные пластинки Пэйе.
| Таблица 17. Органы и другие структурные образования иммунной системы
|
|
|
|
Иммунная система не только подвижная, но и непрерывно обновляющаяся. В то же время постоянство ее состава поддерживается достаточно строго. Хотя принципы этого гомеостаза еще недостаточно понятны, очевидно, что регуляция осуществляется на уровне отдельных разделов (компартментов) системы: численность и состав клеток костного мозга, тимуса и периферических лимфоидных органов регулируются самостоятельно и относительно независимо друг от друга, причем периферические лимфоидные органы регулируются как единое целое.
Уникальная особенность лимфоидных клеток состоит в том, что для их терминального созревания во внутренней среде организма в норме отсутствуют необходимые стимулы: для этого требуется поступление извне или образование внутри (вследствие патологического процесса) чужеродной субстанции — антигена. Лишь при появлении последнего во внутренней среде организма лимфоциты превращаются в эффекторные клетки, способные выполнять иммунологические функции, которые и служат конечными стадиями их развития.
|
|
|
Костный мозг. В костном мозгу (рис. 387) образуются все форменные элементы крови, включая лейкоциты — клетки, непосредственно связанные с иммунной системой, костный Часть клеток, развивающихся из костномозговых предшественников и мозг относящихся к иммунной системе, практически не определяется в крови, таковы дендритные и тучные клетки. Кроветворная ткань представлена в костном мозгу цилиндрическими скоплениями вокруг артериол, образующими шнуры, отделенные друг от друга венозными синусоидами. Они расположены радиально и впадают в центральный синусоид. Клетки различных типов располагаются в \ кроветворной ткани островками. Наибольшее число стволовых элементов iif^ сосредоточено в периферической части просвета костномозгового канала. Рядом с ними вокруг разветвлений артериол располагаются лимфоидные и моноцитарные Рисунок элементы, тогда как предшественники нейтрофильных гранулоцитов сосредото -
387. чены в центре гемопоэтических долек. Размножающиеся и созревающие кроветворные клетки располагаются в петлях, которые образуют ретикулярные клетки. По мере созревания клетки (в частности, лимфоидные) перемещаются из периферической части просвета к центру, где они проникают в синусоиды и поступают в кровоток.
|
|
|
На долю миелоидных элементов в костном мозгу человека приходится 60—65 % клеток, на долю лимфоидных — около 10-15 % кариоцитов. Примерно 60 % лимфоидных клеток находится в процессе созревания, остальные — зрелые клетки, готовые к эмиграции из костного мозга или, наоборот, мигрировавшие в костный мозг из крови. Доля ежедневно эмигрирующих лимфоцитов составляет у мышей около 50 % от общего числа лимфоидных элементов в костном мозгу (180 из 230-320 млн клеток). Обратный приток лимфоцитов из крови в костный мозг в 10 раз меньше (20 млн клеток в день). Содержание в костном мозгу лимфоцитов В-ряда выше, чем Т-клеток и их предшественников. В костном мозгу содержится 1-2 % плазматических клеток, что можно рассматривать как отражение иммунных реакций, реализуемых в костном мозгу.
В-росток лимфопоэза отличается большой интенсивностью функционирования. За сутки у мыши образуется 20-50 млн В-клеток, что значительно превосходит число клеток, требующихся для обновления популяции В-лимфоцитов. Полагают, что значительная часть образующихся клеток гибнет на стадии преВ-лимфоцитов. В настоящее время признано, что костный мозг служит основным местом дифференцировки В-лимфоцитов у млекопитающих (хотя признается роль в В-лимфопоэзе групповых лимфатических фолликулов толстой кишки у овец и некоторые другие особые случаи экстрамедуллярной дифференцировки В-клеток). Небольшое количество (7-8%) кариоцитов костного мозга приобретает маркеры Т-клеток под влиянием дифференцировочных факторов, т.е. представляют собой предшественники Т- лимфоцитов. Часть из них экспрессирует маркеры Т-лимфоцитов — CD7, 2 и 5.
|
|
|
Формирование этих маркеров зависит от гормонов тимуса, приносимых в костный мозг с кровью. У клеток-предшественников, испытавших воздействие этих гормонов, повышается способность к миграции в тимус, в котором они продолжают свое развитие. На
костномозговой стадии развития предшественники Т-лимфоцитов стимулируют (при участии выделяемого ими гуморального фактора) пролиферацию стволовых клеток.
До 4% клеток костного мозга (20-30% от общего числа его лимфоидных клеток) несут маркеры зрелых Т-лимфоцитов, в частности CD3. Часть из них — Т-клетки, мигрировавшие из крови, — является представителями рециркулирующего пула лимфоцитов. Для этих клеток характерно преобладание СD8+-лимфоцитов над СD4+-лимфоцитами: соотношение CD4+/CD8+ составляет в костном мозгу 0,5-1,0 (в крови 1,5-2,0). Миграция зрелых Т- лимфоцитов, а также части тимоцитов в костный мозг усиливается под влиянием гормонов коры надпочечников. В связи с этим содержание Т-клеток в костном мозгу увеличивается при стрессе. Физиологический смысл этого явления неясен; возможно, это связано с усилением процесса образования антител в костном мозгу вследствие миграции сюда Т-клеток, но это объяснение имеет отношение лишь к миграции СD4+-клеток.
В то же время для костного мозга характерна субпопуляция Т-клеток, имеющих фенотип CD3+CD4"CD8". На их долю приходится 60% СDЗ+-клеток. Среди них 70-80% несут рецептор ав-типа, а 20-30% — у5-типа. Первые похожи на аналогичные клетки печени, но лишены (у мышей) маркера В220. Полагают, что они дифференцируются локально, минуя тимус.
Тимус (вилочковая железа) (рис. 388). В отличие от костного мозга, совмещающего функции кроветворного органа и центрального органа иммунной системы, в котором детерминируется развитие лимфоидных клеток и созревают В-лимфоциты, тимус специализирован исключительно на развитии Т-лимфоцитов (а также, как выяснилось в последнее время, миелоидных элементов собственного микроокружения). Это отражает особую сложность процесса развития Т-лимфоцитов.
|
|
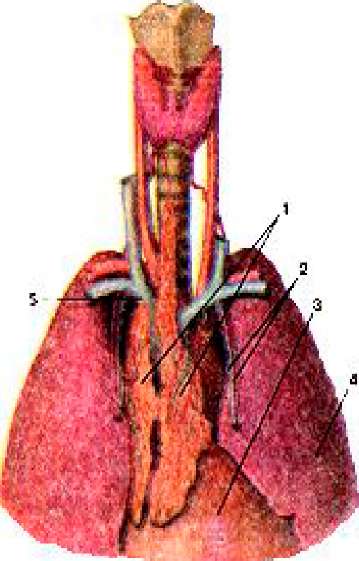
Рисунок 388. Тимус (Thimus). Положение тимуса в грудной полости. Вид спереди. 1 - тимус (правая и левая доли); 2 - внутренние грудные артерия и вена; 3 - перикард; 4 - левое лёгкое; 5 - плечеголовная вена (левая).
Тимус представляет собой лимфоэпителиальный орган. Эпителиальная часть органа является стабильной составляющей, имеющей местное происхождение, а лимфоидные элементы (собственно тимоциты) являются транзиторными: их предшественники мигрируют в тимус из костного мозга (в эмбриональном периоде из печени), а большая часть созревших в тимусе Т-лимфоцитов эмигрирует в периферический отдел иммунной системы, где они включаются в функциональный рециркулирующий пул Т-клеток. Основное назначение тимуса и состоит в формировании данного пула, что включает в себя:
— созревание Т-лимфоцитов, в частности появление у них антигенраспознающих рецепторов;
— дифференцировку Т-клеток на субпопуляции;
— отбор (селекцию) клонов Т-лимфоцитов, способных распознавать чужеродные пептиды в комплексе с аутологичными продуктами МНС.
Эти процессы осуществляются путем воздействия на предшественники Т-лимфоцитов и созревающие тимоциты клеточных и гуморальных факторов микроокружения, создаваемого элементами стромы тимуса.
Тимус состоит из двух долей, объединенных друг с другом. Каждая доля ограничена капсулой, от которой внутрь ткани отходят перегородки, делящие ее на дольки на уровне наружной части органа — коры. Внутренняя часть органа — мозговая — едина для каждой доли.
Тимус как бы разделен на два пространства (компартмента). Одно из них представлено обычной соединительнотканной стромой, сопровождающей сосуды и нервы. Строма образована фибробластами, эндотелием капилляров, волокнами; в периваскулярном пространстве присутствуют макрофаги, реже — плазматические и тучные клетки. Большая же часть объема органа приходится на второй — эпителиальный (внутритимусный) — компартмент, образованный трехмерным каркасом из эпителиальных клеток и ограниченный этими же клетками снаружи.
Периферические лимфоидные органы. Как отмечалось, периферический отдел иммунной системы включает систему органов (лимфатические узлы, селезенка, лимфоидные структуры и диффузная лимфоидная ткань, связанные со слизистыми оболочками и кожей), объединенных системой рециркуляции с относительной автономией субсистем, которые связаны с различными отделами слизистых оболочек (желудочно-кишечного тракта, бронхолегочного аппарата, урогенитального тракта) и кожи.
Лимфатические узлы дренируют лимфу с определенных регионов тела и контролируют появление в них чужеродных объектов — антигенов и их носителей. В некоторых отделах организма, например вдоль крупных сосудов на брыжейке, узлы располагаются цепочками или образуют конгломераты. Они имеют бобовидную форму и размеры от зерна до миндального ореха. Приносящие лимфатические и кровеносные сосуды проникают в узел с его выпуклой стороны и впадают в краевой синус, через который лимфа проникает в ткань коры. Узел покрыт капсулой, от которой внутрь органа отходят тонкие соединительнотканные перегородки — трабекулы. Эфферентные сосуды выходят с вогнутой поверхности узла.
Лимфатический узел (рис. 389) содержит наружную, корковую и более глубокую, медуллярную, зоны. В наружной части коры имеются фолликулы, которые служат средоточием В-лимфоцитов; среди стромальных элементов фолликулов находятся фолликулярные дендритные клетки.
|
|
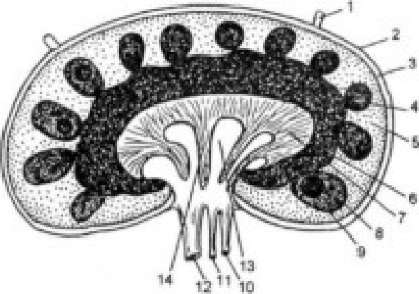
Рисунок 389. Строение лимфатического узла. 1 - афферентный лимфатический сосуд; 2 - капсула; 3 - краевой синус; 4 - первичный лимфоидный фолликул; 5 - наружные слои коры (перифолликулярное пространство); 6 - паракортикальная зона; 7 - мозговой слой; 8 - вторичный фолликул; 9 - зародышевый центр; 10 - артерия; 11 - вена; 12 - эфферентный лимфатический сосуд; 13 - медуллярный синус; 14 - медуллярный шнур.
При развитии иммунного ответа в фолликулах появляются центры размножения (содержащий их фолликул называется вторичным). При этом дендритные клетки длительное время сохраняют на своей поверхности антиген, что является условием для формирования в зародышевом центре клеток памяти. В пространстве, окружающем фолликулы, содержатся как В-, так и Т-лимфоциты. Фолликулярные дендритные клетки отсутствуют у мышей с разрушенными генами лимфотоксинов а и в; параллельно блокируются развитие лимфатических узлов и формирование центров размножения в фолликулах селезенки. Очевидно, «выпадающие» морфогенетические процессы контролируются лимфотоксинами.
Мозговая зона лимфатических узлов содержит мякотные шнуры, образованные ретикулиновыми волокнами, лимфоцитами обоих классов (с преобладанием В-лимфоцитов) и плазматическими клетками, число которых особенно увеличивается при иммунном ответе. Между мякотными шнурами находятся медуллярные синусы — скопления лимфы, собираемой в эфферентные лимфатические сосуды. В узлах преобладают Т-лимфоциты. Хотя в них развивается как гуморальный, так и клеточный ответ на антигены, полагают, что второй тип ответа в большей степени отражает специфику иммунологической функции лимфатических узлов. В основе этого лежат особенности микроокружения лимфатических узлов, которые в условиях антигенной стимуляции способствуют дифференцировке CD4+-клеток преимущественно в направлении «воспалительных» хелперов типа ТЫ. Последние обеспечивают развитие клеточных иммунных реакций. Отличия в структуре и функции лимфатических узлов, локализующихся в различных отделах организма, невелики. Наибольшим своеобразием отличаются брыжеечные лимфатические узлы, в которых несколько выше процент В- лимфоцитов, особенно несущих мембранный IgA. Кроме того, они занимают особое место в рециркуляции лимфоцитов.
Селезенка расположена на гематогенных путях распространения антигенов, что отличает ее от лимфатических узлов, контролирующих лимфатические пути, и обусловливает единичность этого органа (в отличие от множественности лимфатических узлов, дренирующих определенные регионы тела). Селезенка обладает более комплексными функциями, чем лимфатический узел: у многих животных в ней осуществляется гемопоэз, она служит фильтром для старых эритроцитов, резервуаром, регулирующим объем циркулирующей крови и т.д.
Селезенка (рис. 390) окружена плотной капсулой, от которой внутрь отходят трабекулы, несущие сосуды (как и в лимфатических узлах). Содержимое селезенки представляет собой массу красного цвета (красная пульпа), в которую вкраплены белесоватые зерновидные тельца (белая пульпа).
Белая пульпа связана с артериолами селезенки и имеет наиболее прямое отношение к иммунологической функции органа. Артериолы, отходящие от трабекулярных
| I |
ССЛвЭЁНМЭ артерий, окружены лимфоидными скоплениями, муфтами, образованными в
у^в основном Т-лимфоцитами и являющимися тимусзависимыми зонами селезенки.
Муфты соседствуют с фолликулами, аналогичными фолликулам лимфатических узлов и образованными В-лимфоцитами. В фолликулах от артериол отделяются капилляры, открывающиеся в ткань, которая окружает фолликулы, и муфты — маргинальную зону. Последняя отделяет белую пульпу от красной, в которую Рисунок 390. переходит артериола. В маргинальной зоне В-лимфоциты преобладают, но здесь присутствуют и Т-клетки. Плотность лимфоцитов в маргинальной зоне меньше, чем в фолликулах и муфтах. Структура стромы тимусзависимых и тимуснезависимых зон селезенки напоминает таковую аналогичных зон лимфатических узлов и характеризуется теми же особыми типами клеток.
В красной пульпе наряду с элементами крови, сосредоточенными в расширенных сосудах — синусоидах, содержатся губчатые скопления ткани, в которой присутствуют макрофаги, плазматические клетки и лимфоциты обоих классов. Плазматические клетки обнаруживаются в селезенке даже вне явно выраженных иммунных реакций. Это связано с тем, что в организме всегда присутствуют антигены (например, пищевые), на которые реагирует прежде всего селезенка. «Спонтанные» антителообразующие клетки селезенки и являются отражением этого фонового уровня иммунных реакций организма, тем более что селезенка служит тем
органом периферического отдела иммунной системы, в котором существуют оптимальные условия именно для развития гуморального ответа.
В селезенке В-лимфоциты преобладают над Т-клетками, что также свидетельствует о преимущественной ориентации органа на развитие гуморального иммунного ответа (в противоположность лимфатическим узлам). Среди Т-лимфоцитов преобладают, как и в других периферических органах иммунной системы, Т-хелперы. Однако в красной пульпе сосредоточены почти исключительно Т-клетки с супрессорной активностью. Здесь имеются также 0-клетки (т.е. клетки, лишенные маркеров Т- и В-лимфоцитов), в том числе значительное число NK-киллеров. В селезенке много макрофагов, и уровень их функциональной активности достаточно высок. В связи с тем, что селезенка служит хранилищем крови, регулятором ее циркуляторного объема и местом, где задерживаются (при участии макрофагов) старые эритроциты, в суспензиях спленоцитов всегда имеется много красных кровяных элементов.
| Рисунок 392. Лимфоидные узелки в стенке аппендикса. Поперечный срез. 1 - стенка аппендикса; 2 - лимфоидные узелки; 3 - эпителиальный покров. |
Лимфоидные ткань и структуры, связанные со слизистыми оболочками. Лимфоидные структуры (рис. 391-394) и диффузные лимфоциты свойственны всем основным типам слизистых оболочек — в пищеварительном, бронхолегочном и урогенитальном трактах. Это огромный по объему отдел иммунной системы: площадь поверхности слизистых оболочек у человека составляет 400 м2 (для сравнения: поверхность кожи — 1,8 м2).
|
|
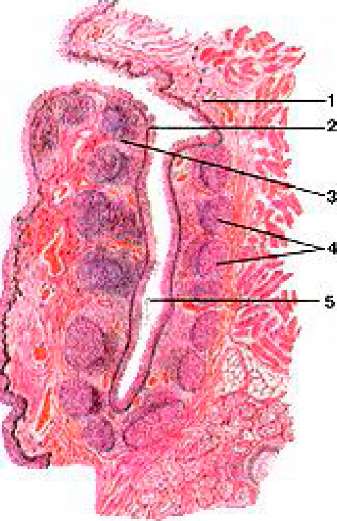
Рисунок 391. Небная миндалина (tonsilla palatina). Поперечный разрез. 1 - слизистая оболочка; 2 - многослойный плоский эпителий; 3 - околоузелковая лимфоидная ткань; 4 -
лимфоидные узелки; 5 - просвет крипты.
| 2
|
|
|
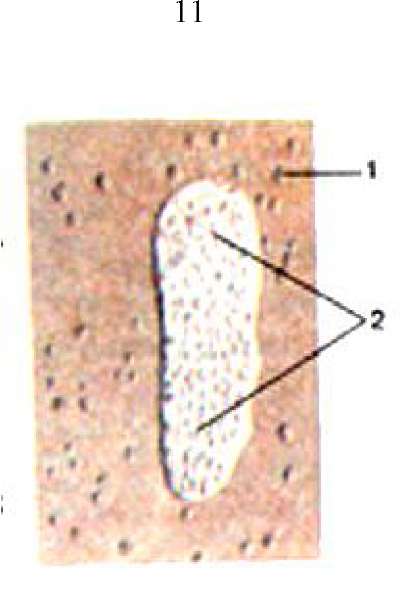
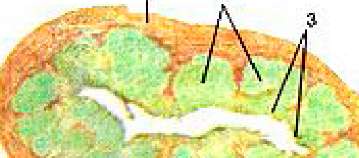
Рисунок 393. Лимфоидные узелки и лимфоидная бляшка в стенке тонкой кишки. I - лимфоидные узелки; 2 - лимфоидная бляшка
Различают структурированную и диффузную составляющие лимфоидной ткани, ассоциированной со слизистыми оболочками. Первая включает в себя единичные некапсулированные фолликулы, а также такие организованные формирования лимфоидной ткани, как миндалины, аппендикс, групповые лимфатические фолликулы (пейеровы бляшки). Вторая составляющая представлена единичными клетками, инфильтрирующими эпителиальные пласты слизистых оболочек (Т-лимфоциты) и собственную пластинку (lamina propria), а также подслизистый слой (преимущественно В-лимфоциты). Другая классификация лимфоидных образований, связанных со слизистыми оболочками, основана на ПЛаСТННЙИ учете места этих образований в иммунных процессах: их разделяют на ПеЙе индуктивные и эффекторные участки. К первым в пищеварительном тракте
Рисунок 394. относятся миндалины, групповые лимфатические фолликулы и региональные лимфатические узлы, ко вторым — межэпителиальные лимфоциты и собственная пластинка (Пейе). Первые осуществляют процесс восприятия и первичной обработки антигена, вторые реализуют иммунные реакции.
Дата добавления: 2018-09-20; просмотров: 365; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!