У яйцеклетки наличие особых оболочек, у сперматозойда акросомальный аппарат. По количеству желтка
Биология. Жизнь. Происхождение жизни
Термин биология (от греч БИОС-жизнь, логос-наука) введен в начале 19 в. Ламарком и Тревиранусом. БИОЛОГИЯ, наука о жизни, включающая все знания о природе, структуре, функциях и поведении живых существ. Биология имеет дело не только с великим множеством форм различных организмов, но также с их эволюцией, развитием и с теми отношениями, которые складываются между ними и окружающей средой. Жизнь- процесс идущий с затратой энергии из внешней среды заключающийся в развитии, саморегуляции и самовоспроизведении. Св-ва жизни-1)обмен веществ (ассимиляция-образование и обновление структур организма; диссимиляция- расщепление органических веществ с целью обеспечить необходимыми веществами и энергией)
2)Без воздействия извне процессы в системах идут в одном направлении: теплота переходит к более холодному. 3) раздражимость, возбудимость 4)продолжительность жизни ограничена, поэтому сохранение жизни зависит от размножения –Включенность организмов в процесс эволюции 4) существование отдельных организмов лишь во взаимодействии с другими в составе сообщест –биоценозов (совокупность растений, животных, микроорганизмов, населяющих участок суши или водоёма и характеризующихся определёнными отношениями как между собой, так и с абиотическими факторами среды). 5) рост развитие 6)Изменчивость 7)Клеточное строение. Происхождение жизни: 1) Возможность абиогенного образования органических веществ, включая мономеры биологических полимеров, в условиях, бывших на Земле около 4 млрд. лет назад, доказана опытами химиков. В лабораторных условиях при пропускании электрических разрядов через различные газовые смеси, напоминающие примитивную атмосферу планеты, а также при использовании других источников энергии ученые получали среди продуктов реакций аминокислоты (аланин, глицин, аспарагиновую кислоту), янтарную, уксусную, молочную кислоты, мочевину, азотистые основания (аденин, гуанин), АДФ и АТФ. Низкомолекулярные органические соединения накапливались в водах первичного океана в виде первичного бульона или же адсорбировались на Поверхности глинистых отложений. Последнее повышало концентрацию этих веществ, создавая тем самым лучшие условия для полимеризации.
|
|
|
2) Возможность полимеризации низкомолекулярных соединений с образованием полипептидов и полинуклеотидов (определяющая следующий этап на пути возникновения жизни)
Ученые предполагают, что образование полипептидов и полинуклеотидов могло происходить в пленке из низкомолекулярных органических соединений в безводной среде, например на склонах вулканических конусов, покрытых остывающей лавой. Это предположение находит подтверждение в опытах. Выдерживание в течение определенного времени при 130°С сухой смеси аминокислот в сосудах из кусков лавы приводило к образованию полипептидов.
|
|
|
Образующиеся описанным образом биополимеры смывались ливневыми потоками в первичный бульон, что защищало их от разрушающего действия УФ-излучения, которое в то время из-за отсутствия в атмосфере планеты озонового слоя было очень жестким.
По мере повышения концентрации полипептидов, полинуклеотидов и других органических соединений в первичном бульоне сложились условия для следующего этапа — самопроизвольного возникновения предбиологических форм сложного химического состава, или протобионтов. Предположительно они могли быть представлены коацерватами (А. И. Опарин) или микросферами (С. Фоке). Это коллоидные капли с уплотненным поверхностным слоем, имитирующим мембрану, содержимое которых составляли один или несколько видов биополимеров. Возможность образования в коллоидных растворах структур типа коацерватов или микросфер доказана опытным путем.
3) гипотеза по которой жизнь занесена на Землю в виде спор микроорганизмов инопланетными существами –панспермия
|
|
|
2. 2) Живая система – определение и свойства. Уровни организации живых систем.
Экосистема – это функциональная система, состоящая из живых и неживых компонентов, объединенных круговоротом веществ и трансформацией энергии. Живое без неживого существовать не может. Даже организм как наиболее организованная живая система включает неорганические компоненты и беспрерывно поглощает и выделяет неживые вещества. Следовательно, все экологические системы, в том числе и биосфера, - это системы живые, они подчиняются законам движения живой материи. Уровни организации:1) молекулярно-генетический - Элементарной единицей на молекулярно-генетическом уровне служит ген — фрагмент молекулы нуклеиновой кислоты, в котором записан определенный в качественном и количественном отношении объем биологической (генетической) информации. Элементарное явление заключается прежде всего в процессе конвариантной редупликации, или самовоспроизведении, с возможностью некоторых изменений в содержании закодированной в гене информации. Путем редупликации ДНК происходит копирование заключенной в генах биологической информации, что обеспечивает преемственность и сохранность (консерватизм) свойств организмов в ряду поколений. Редупликация, таким образом, является основой наследственности.
|
|
|
2) клеточный уровень. клетка, служащая элементарной структурой клеточного уровня. Элементарное явление представлено реакциями клеточного метаболизма, составляющими основу потоков энергии, веществ и информации. Благодаря деятельности клетки поступающие извне вещества превращаются в субстраты и энергию, которые используются (в соответствии с имеющейся генетической информацией) в процессе биосинтеза белков и других соединений, необходимых организму. Таким образом, на клеточном уровне сопрягаются механизмы передачи биологической информации и превращения веществ и энергии. Элементарное явление на этом уровне служит энергетической и вещественной основой жизни на всех других уровнях ее организации.
3) Элементарной единицей организменного уровня является особь в ее развитии от момента зарождения до прекращения существования в качестве живой системы, что позволяет также назвать этот уровень онтогенетическим. Закономерные изменения организма в индивидуальном развитии составляют элементарное явление данного уровня. Эти изменения обеспечивают рост организма, дифференциацию его частей и одновременно интеграцию развития в единое целое, специализацию клеток, органов и тканей. В ходе онтогенеза в определенных условиях внешней среды происходит воплощение наследственной информации в биологические структуры и процессы, на основе генотипа формируется фенотип организмов данного вида.
4) Элементарной единицей популяционно-видового уровня служит популяция — совокупность особей одного вида. Объединение особей в популяцию происходит благодаря общности генофонда, используемого в процессе полового размножения для создания генотипов особей следующего поколения. Популяция в силу возможности межпопуляционных скрещиваний представляет собой открытую генетическую систему. Действие на генофонд популяции элементарных эволюционных факторов, таких, как мутационный процесс, колебания численности особей, естественный отбор, приводит к эволюционно значимым изменениям генофонда, которые представляют элементарные явления на данном уровне.
5) Организмы одного вида населяют территорию с известными абиотическими показателями (климат, химизм почв, гидрологические условия) и взаимодействуют с организмами других видов. В процессе совместного исторического развития на определенной территории организмов разных систематических групп образуются динамичные, устойчивые во времени сообщества — биогеоценозы, которые служат элементарной единицей биогеоценотического (экосистемного) уровня. Элементарное явление на рассматриваемом уровне представлено потоками энергии и круговоротами веществ. Ведущая роль в этих круговоротах и потоках принадлежит живым организмам. Биогеоценоз — это открытая в вещественном и энергетическом плане система. Биогеоценозы, различаясь по видовому составу и характеристикам абиотической своей части, объединены на планете в единый комплекс — область распространения жизни, или биосферу.
3. 3) Молекулярно-генетический уровень организации жизни. Нуклеиновые кислоты и белки, их строение и значение. Генетический код и его свойства.
Элементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, является ген . Под признаком понимают отдельное качество или свойство, по которому они отличаются друг от друга. сложные признаки, формирование которых требует синтеза многих веществ, в первую очередь белков со специфическими свойствами. Свойства белковой молекулы определяются аминокислотной последовательностью ее полипептидной цепи, которая прямо задается последовательностью нуклеотидов в ДНК соответствующего гена и является элементарным, или простым, признаком.
Нуклеиновые кислоты состоят из нуклеотидов, которые включают в себя сахар(пентозу), фосфат и азотистое основание (пурин или пиримидин). РНК и ДНК-по принципу комплементарности и антипараллельности. Есть 3 структуры: первичную структуру —полинуклеотидную цепь, вторичную структуру—две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями, и третичную структуру — трехмерную спираль с приведенными выше пространственными характеристиками.
Генетический код-это способ записи информации о структуре белков в генетическом материале. Св-ва генетического кода:1) триплетность-аминокислоты шифруются 3 рядом стоящими нуклеотидами 2) специфичность- каждый триплет кодирует одну аминокислоту 3) избыточность (вырожденность)-большинство аминокислот кодируется несколькими триплетами. 4. Универсальность всех живых организмов населяющих Землю одни и те же аминокислоты шифруются одинаковыми триплетами 5. Непрерывность и неперекрываемость при расшифровке информации
4. 4) Клеточный уровень организации жизни, происхождение и эволюция клеток. Принципы структурно-функциональной организации про- и эукариотических клеток.
клетка, служащая элементарной структурой клеточного уровня. Элементарное явление представлено реакциями клеточного метаболизма, составляющими основу потоков энергии, веществ и информации. Благодаря деятельности клетки поступающие извне вещества превращаются в субстраты и энергию, которые используются (в соответствии с имеющейся генетической информацией) в процессе биосинтеза белков и других соединений, необходимых организму. Таким образом, на клеточном уровне сопрягаются механизмы передачи биологической информации и превращения веществ и энергии. Элементарное явление на этом уровне служит энергетической и вещественной основой жизни на всех других уровнях ее организации.
симбиотическая гипотеза происхождения эукариотических клеток, согласно которой (рис. 1.4) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза.
Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли.
митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот (рис. 1.4). Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран.
Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки.
История показала, что эволюционные возможности клеток эукариотического типа несравнимо выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по размерам геном прокариот. Количество генов у бактерии и в клетке человека, например, соотносится как 1: (100-1000). Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов. Это расширяет масштабы мутационной изменчивости без угрозы резкого снижения жизнеспособности, эволюционно значимым следствием чего является образование резерва наследственной изменчивости.
При переходе к эукариотическому типу усложняется механизм регуляции жизнедеятельности клетки, что на уровне генетического материала проявилось в увеличении относительного количества регуляторных генов, замене кольцевых «голых» молекул ДНК прокариот хромосомами, в которых ДНК соединена с белками. В итоге стало возможным считывать биологическую информацию по частям с разных групп генов в разном их сочетании в различных типах клеток и в разное время. В бактериальной клетке, напротив, одновременно считывается до 80—100% информации генома. В клетках взрослого человека в разных его органах транскрибируется от 8—10% (печень, почка) до 44% (головной мозг) информации. Использованию биологической информации частями принадлежит исключительная роль в эволюции многоклеточных организмов, так как именно это позволяет разным группам клеток специализироваться по различным функциональным направлениям.
Большое значение при переходе к многоклеточности имело наличие у эукариотических клеток эластичной оболочки, что необходимо для образования устойчивых клеточных комплексов.
В природе существует значительное разнообразие конкретных клеточных форм. Вместе с тем число основных типов клеточной организации ограниченно. Выделяют прокариотический и эукариотический типы с подразделением второго на подтип клеток простейших организмов и подтип клеток многоклеточных (рис. 1).
Клетки прокариотического типа имеют особенно малые размеры — не более 0,5—3,0 мкм в диаметре. У них нет морфологически обособленного ядра, так как ядерный материал в виде ДНК не отграничен от цитоплазмы оболочкой. В клетке отсутствует развитая система мембран. Генетический аппарат образован единственной кольцевой хромосомой, которая лишена основных белков — гистонов. Гистоны являются белками клеточных ядер. Благодари значительному количеству основных аминокислот аргинина и лизина они имеют щелочной характер. В хромосомах эукариотических клеток гистоны, соединяясь с ДНК, подавляют процесс считывания биологической информации. Различия прокариотических и эукариотических клеток но содержанию гистонов указывают на разные механизмы регуляции функции наследственного материала.
5) Временная организация клетки. Понятие о клеточном и митотическом цикле. Характеристика фаз митотического цикла
Клеточный цикл — это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Важным компонентом клеточного цикла является митотический (пролиферативный) цикл —комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления.
Интерфаза (постмитотический, пресинтетический или G1 период) длится от нескольких часов до нескольких суток. Дочерние клетки растут, увеличивается Объем цитоплазмы, идет активный синтез белка, углеводов, липидов, ферментов. В синтетическом периоде удваивается количество наследственного материала. Постсинтетическом периоде (предмитотический G2) интенсивный синтез РНК и особенно белка. Завершается удвоение массы цитоплазмы.
| Фаза митоза | Содержание изменений |
| Профаза Метафаза Анафаза Телофаза | Хромосомы спирализуются и приобретают вид нитей. Ядрышко разрушается. Распадается ядерная оболочка. В цитоплазме уменьшается количество структур шероховатой сети. Резко сокращается число полисом. Центриоли клеточного центра расходятся к полюсам клетки, между ними микротрубочки образуют веретено деления Заканчивается образование веретена деления. Хромосомы выстраиваются в экваториальной плоскости клетки (метафазная пластинка). Микротрубочки веретена деления связаны с кинетохорами хромосом. Каждая хромосома продольно расщепляется на две хроматиды (дочерние хромосомы), соединенные в области кинетохора Связь между хроматидами нарушается, и они в качестве самостоятельных хромосом перемещаются к полюсам клетки со скоростью 0,2—5 мкм/мин. По завершении движения на полюсах собирается два равноценных полных набора хромосом Реконструируются интерфазные ядра дочерних клеток. Хромосомы де-спирализуются. Образуются ядрышки. Разрушается веретено деления. Материнская клетка делится на две дочерние |
6) Самовоспроизводство генетического материала, репликация, репликон. Особенности репликации и про- и эукаритор
Одним из основных свойств материала наследственности является его способность к самокопированию —репликация. Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали. Такой способ удвоения молекул, при котором каждая дочерняя молекула содержит одну материнскую и одну вновь синтезированную цепь, называют полуконсервативным. Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул.
Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации — репликон.
Инициация репликации- точки ori (ори), состоящих из 300 пар нуклеотидов, от них в противоположном направлении расходяться репликационные вилки, образуется репликационный глазок. Белок геликаза разрывает водородные связи. Белок топоизомераза разрывает одну из цепе ДНК. Полимераза присоединяет очередной нуклеотид к OH-группе в 3’ положении, цепь удлиняется на этом конце. Для того чтобы полимераза начала синтезировать новую полинуклеотидную цепь нужна затравка (короткая последовательность РНК, образуемые при участии белка праймазы. Это означает что матрицей при репликации может служить цепь, которая имее свободный 3’ OH конец. На второй цепи синтез осуществляется фрагментом ОКАЗАКИ, также от 5 к 3 концу. Из двух синтезируемых цепей одна строится непрерывно, ее синтез идет быстрей –лидирующая цепь, а другая отстающая. Результат репликации-две молекулы ДНК, нуклеотдиная последовательность которой индетична материнской. Репликация у про- и эукариот протекает практически индетично, одна скорость синтеза у эукариот ниже, чем у прокариот. Днк прокариот имеет один локус ori и представляет собой целиком отдельные репликоны. Эукариотические хромосомы содержат большее колличсетво репликонов(репликация начинается в нескольких точкаХ)
7) Размножение как свойство жизни. Сравнительная характеристика и многообразие форм бесполого и полового размножения. Биологическое значение размножения.
Размножение-свойство живых организмов передача в ряду поколений генетического материала (ДНК), т.е. определенной, специфичной для данного вида биологической информации. Бесполое размножение возникло первей. Существуют различные формы бесполого размножения: множественное деление (шизогония) например малярийный плазмодий, почкование- сначало на родительской особи формируется выступ, который потом отшнуровывается, деление на двое бывает продольным -медузы, поперечное-кольчатые черви, фрагментация распад тела многоклеточного организма на части, которые далее превращаются в самостоятельных особей(иглокожие, плоские черви), размножение спорами. Различия: для размножения Б. нужна одна особь, для П 2 особи; Клеточный источник наследственной информации для развития потомка Б. многоклеточные-одна или несколько соматических клеток, одноклеточные -клетка организм как целое, П родители образуют гаметы, родитель представлен в потомке исходно одной клеткой. Главный клеточный механизм: Б-митоз, П-мейоз. Эволюционное значение: Б. способствует поддержанию наибольшей приспособленности в маломеняющихся условиях обитания, усиливает роль стабилизирующего естественного отбора. П за счет генетического разнообразия создает предпосылки к освоению разнообразных условий обитания, способствует осуществлению творческой роли естественного отбора. Б 2n, П-n, Б сохраняет вид без изменений, П-появление внутривидовой изменчивости.
Биологическая роль размножения состоит в том что обеспечивает смену поколений, с его помощью сохраняются во времени биологические виды и жизнь. Также увелечение числа особей, воспроизведение себе подобнового.
8)Митоз, его биологическое значение, нарушение митоза и их роль в возникновении соматических мутаций.
5 вопрос. Нарушение фаз митоза приводит паталогичискем изменениям клеток. Отклонение от нормального течения процесса спирализации может привести к набуханию и слипанию хромосом. Иногда наблюдается отрыв участка хромосомы, который, если он лишен центромеры, не участвует в анафазном перемещении к полюсам и теряется. Отставать при движении могут отдельные хроматиды, что приводит к образованию дочерних ядер с несбалансированными хромосомными наборами. Повреждения со стороны веретена деления приводят к задержке митоза в метафазе, рассеиванию хромосом. При изменении количества центриолей возникают многополюсные или асимметричные митозы. Нарушение цитотомии приводит к появлению дву- и многоядерных клеток.
Отклонение от нормального течения процесса спирализации может привести к набуханию и слипанию хромосом. Иногда наблюдается отрыв участка хромосомы, который, если он лишен центромеры, не участвует в анафазном перемещении к полюсам и теряется. Отставать при движении могут отдельные хроматиды, что приводит к образованию дочерних ядер с несбалансированными хромосомными наборами. Повреждения со стороны веретена деления приводят к задержке митоза в метафазе, рассеиванию хромосом. При изменении количества центриолей возникают многополюсные или асимметричные митозы. Нарушение цитотомии приводит к появлению дву- и многоядерных клеток.
Удвоение ДНК клетки не всегда сопровождается ее разделением на две. Поскольку механизм такого удвоения совпадает с предмитотической редупликацией ДНК и оно сопровождается кратным увеличением количества хромосом, это явление получило название эндомитоза. Другое явление, сходное по результату и названное политенией, заключается в кратном увеличении содержания ДНК в хромосомах при сохранении их диплоидного количества. Эндомитоз и политения приводят к образованию полиплоидных клеток, отличающихся кратным увеличением объема наследственного материала. В таких клетках в отличие от диплоидных гены повторены более чем два раза. Пропорционально увеличению числа генов растет масса клетки, что повышает ее функциональные возможности. В организме млекопитающих полиплоидизация с возрастом свойственна печеночным клеткам.
9)Мейоз, его биологическое значение, паталогии мейоза и их роль в возникновении генеративных мутаций.
При последующем оплодотворении гаметы формируют организм нового поколения с диплоидным кариотипом (пс + пс == 2n2c). В этом заключается важнейшее биологическое значение мейоза. Мейоз состоит из двух быстро следующих одно за другим делений, происходящих в периоде созревания. Удвоение ДНК для этих делений осуществляется однократно в периоде роста. Второе деление мейоза следует за первым практически сразу так, что наследственный материал не синтезируется в промежутке между ними
Первое мейотическое деление называют редукционным, так как оно приводит к образованию из диплоидных клеток (2п2с) гаплоидных клеток п2с. Такой результат обеспечивается благодаря особенностям профазы первого деления мейоза. В профазе I мейоза, так же как в обычном митозе, наблюдается компактная упаковка генетического материала (спирализация хромосом). Одновременно происходит событие, отсутствующее в митозе: гомологичные хромосомы конъюгируют друг с другом, т.е. тесно сближаются соответствующими участками.
В результате конъюгации образуются хромосомные пары, или биваленты, числом п. Так как каждая хромосома, вступающая в мейоз, состоит из двух хроматид, то бивалент содержит четыре хроматиды. Формула генетического материала в профазе I остается 2n4c. К концу профазы хромосомы в бивалентах, сильно спирализуясь, укорачиваются. Так же как в митозе, в профазе I мейоза начинается формирование веретена деления, с помощью которого хромосомный материал будет распределяться между дочерними клетками (рис. 5.5).
Процессы, происходящие в профазе I мейоза и определяющие его результаты, обусловливают более продолжительное течение этой фазы деления по сравнению с митозом и дают возможность выделить несколько стадий в ее пределах (рис. 5.5).
Лептотена —наиболее ранняя стадия профазы I мейоза, в которой начинается спирализация хромосом, и они становятся видимыми в микроскоп как длинные и тонкие нити. Зиготена характеризуется началом конъюгации гомологичных хромосом, которые объединяются синаптонемальным комплексом в бивалент (рис. 5.6). Пахитена — стадия, в которой на фоне продолжающейся спирализации хромосом и их укорочения, между гомологичными хромосомами осуществляется кроссинговер — перекрест с обменом соответствующими участками. Диплотена характеризуется возникновением сил отталкивания между гомологичными хромосомами, которые начинают отдаляться друг от друга в первую очередь в области центромер, но остаются связанными в областях прошедшего кроссинговера — хиазмах (рис. 5.7).
Диакинез — завершающая стадия профазы I мейоза, в которой гомологичные хромосомы удерживаются вместе лишь в отдельных точках хиазм. Биваленты приобретают причудливую форму колец, крестов, восьмерок и т.д. (рис. 5.8).
Таким образом, несмотря на возникающие между гомологичными хромосомами силы отталкивания, в профазе I не происходит окончательного разрушения бивалентов. Особенностью мейоза в овогенезе является наличие специальной стадии — диктиотены, отсутствующей в сперматогенезе. В метафазе I мейоза завершается формирование веретена деления. Его нити прикрепляются к центромерам хромосом, объединенных в биваленты, таким образом, что от каждой центромеры идет лишь одна нить к одному из полюсов веретена. В результате нити, связанные с центромерами гомологичных хромосом, направляясь к разным полюсам, устанавливают бивалентны в плоскости экватора веретена деления.
В анафазе I мейоза ослабляются связи между гомологичными хромосомами в бивалентах и они отходят друг от друга, направляясь к разным полюсам веретена деления. При этом к каждому полюсу отходит гаплоидный набор хромосом, состоящих из двух хроматид (см. рис. 5.5).
В телофазе I мейоза у полюсов веретена собирается одинарный, гаплоидный набор хромосом, каждая из них содержит удвоенное количество ДНК.
Формула генетического материала образующихся дочерних клеток соответствует п2с.
Второе мейотическое (эквационное) деление приводит к образованию клеток, в которых содержание генетического материала в хромосомах будет соответствовать их однонитчатой структуре пс (см. рис. 5.5). Это деление протекает, как митоз, только клетки, вступающие в него, несут гаплоидный набор хромосом. В процессе такого деления материнские двунитчатые хромосомы, расщепляясь, образуют дочерние однонитчатые.
Одна из главных задач мейоза — создание клеток с гаплоидным набором однонитчатых хромосом —достигается благодаря однократной редупликации ДНК для двух последовательных делений мейоза, а также благодаря образованию в начале первого мейотического деления пар гомологичных хромосом и дальнейшего их расхождения в дочерние клетки.
Кроссинговер обеспечивает перекомбинацию отцовских и материнских аллелей в группах сцепления. Паталогии:1)неравноценный кроссинговер, приводящий к мутациям 2)полное не расхождение хромосом во время мейоза, приводящие к последующей полиплодии организма 3)нерасхождение отдельных пар гомологичных хромосом из-за полного сцепления гомологов, это приводит к различным вариантам ангуэплоидии.
10) Мейоз и оплодотворение как механизмы, обеспечения поддержки постоянства кариотипа в ряду поколений. Комбинативная изменчивость
Биологическое значение М. заключается в поддержании постоянства кариотипа в ряду поколений организмов данного вида и обеспечении возможности рекомбинации хромосом и генов при половом процессе. М. — один из ключевых механизмов наследственности и наследственной изменчивости. Диплойдное ядро зиготы. Таким образом мейоз и последующее оплодотворение обеспечивают сохранение у нового поколения организмов диплойдного кариотпиа, присущего всем особям данного вида.Благодаря особенностям мейоза образуются клетки, несущие полноценный геном, в котором каждая группа сцепления представлена в единственном экземпляре (гаплойдный набор хромосом). Ядра гамет сливаются и образуется Комбинативная изменчивость возникает при свободных скрещиваниях в популяциях или при искусственной гибридизации. В результате рождаются особи с новыми сочетаниями признаков и свойств, которые отсутствовали у родителей. Комбинативная изменчивость, проявляющаяся в генотипическом разнообразии особей, повышает выживаемость вида в изменяющихся условиях его существования.
11) Сравнительная характеристика митоза и мейоза. Биологическое значение этих форм клеток деления.
Митоз обеспечивает размножение самотических клеток, дочерние клетки индетичны материнской, протекает в одну фазу 2n2c. Мейоз-размножение половых клеток, с его помощью происходит перекомбтнация наследственного материала, обеспечивает комбинативную изменчивость, 2 фазы, длится дольше чем митоз, nc. Второе деление мейоза соответствует делению митоза.
12) Гаметогенез, его биологическое значение. Периоды гаметогенеза, отличия ово и сперматогенеза.
Гаметогенез — процесс образования яйцеклеток (овогенез) и сперматозоидов (сперматогенез) — подразделяется на ряд стадий.
В стадии размножения диплоидные клетки, из которых образуются гаметы, называют сперматогониями и овогониями. Эти клетки осуществляют серию последовательных митотических делений, в результате чего их количество существенно возрастает. Сперматогонии размножаются на протяжении всего периода половой зрелости мужской особи. Размножение овогоний приурочено главным образом к периоду эмбриогенеза.
Овогонии и сперматогонии, как и все соматические клетки, характеризуются диплоидностью. Если в одинарном, гаплоидном наборе число хромосом обозначить как n, а количество ДНК — как с, то генетическая формула клеток в стадии размножения соответствует 2n2с до 5-периода и 2n4с после него.
На стадии роста происходит увеличение клеточных размеров и превращение мужских и женских половых клеток в сперматоциты и овоциты I порядка. Важным событием этого периода является редупликация ДНК при сохранении неизменным числа хромосом. Последние приобретают двунитчатую структуру, а генетическая формула сперматоцитов и овоцитов I порядка приобретает вид 2п4с.
Основными событиями стадии созревания являются два последовательных деления: редукционное и эквационное,— которые вместе составляют мейоз. После первого деления образуются сперматоциты и овоциты II порядка (формула п2с), а после второго — сперматиды и зрелая яйцеклетка (пс).
В результате делений на стадии созревания каждый сперматоцит I порядка дает четыре сперматиды, тогда как каждый овоцит I порядка — одну полноценную яйцеклетку и редукционные тельца, которые в размножении не участвуют. Благодаря этому в женской гамете концентрируется максимальное количество питательного материала — желтка.
Процесс сперматогенеза завершается стадией формирования, или спермиогенеза. Ядра сперматид уплотняются вследствие сверхспирализации хромосом, которые становятся функционально инертными. Пластинчатый комплекс перемещается к одному из полюсов ядра. Центриоли занимают место у противоположного полюса ядра, причем от одной из них отрастает жгутик, у основания которого в виде спирального чехлика концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, так что головка зрелого сперматозоида практически ее лишена.
Отлиие: О.-начало в пренатальном периоде. С.-постнатальном.2)С-образование 4х клеток.О.-одна яйцеклетка и 3 тельца. 3)О.-фаза роста выражена, фаза формирования отсутствует
За счет генетического разнообразия половое размножение создает предпосылки к освоению разнообразных условий обитания; дает эволюционные и экологические перспективы; способствует осуществлению творческой роли естественно отбора.
13)Строение и функции половых клеток. Типы яйцеклеток.
У большинства видов по структурным и функциональным признакам половые клетки делятся на материнские (яйцеклетки) и отцовские (сперматозоиды). Как правило, яйцеклетки и сперматозоиды вырабатываются разными организмами — женскими (самки) и мужскими (самцы). В подразделении гамет на яйцеклетки и сперматозоиды, а особей на самок и самцов заключается явление полового диморфизма. В сравнении с другими линиями соматических клеток (эпителиальные, нервные, мышечные) гаметы характеризуются рядом отличий. Важнейшее из них — гаплоидный набор хромосом в ядрах, что обеспечивает воспроизведение в зиготе типичного для организмов данного вида диплоидного числа хромосом.
Гаметы отличаются необычным для других клеток значением ядерно-цитоплазматического отношения. У яйцеклеток оно снижено благодаря увеличенному объему цитоплазмы, в которой размещен питательный материал (желток) для развития зародыша. У сперматозоидов благодаря малому количеству цитоплазмы ядерно-цитоплазматическое отношение высокое. Это находится в соответствии с главной функциональной задачей мужской гаметы — транспортировкой наследственного материала к яйцеклетке.
Половые клетки отличаются низким уровнем обменных процессов, близким к состоянию анабиоза. Мужские гаметы не вступают в митотический цикл. У яйцеклеток эта способность восстанавливается при оплодотворении или действии фактора, активирующего партеногенез.
У яйцеклетки наличие особых оболочек, у сперматозойда акросомальный аппарат. По количеству желтка
- Полилецитальные — содержат большое количество желтка (членистоногие, рыбы, кроме осетровых, рептилии, птицы).
- Мезолецитальные — содержат среднее количество желтка (осетровые рыбы, амфибии).
- Олиголецитальные — содержат мало желтка (моллюски, иглокожие).
- Алецитальные — не содержат желтка (плацентарные млекопитающие).
По расположению желтка
- Телолецитальные — желток смещён к вегетативному полюсу яйцеклетки. Противоположный полюс называется анимальным. Сюда относятся некоторые полилецитальные (рыбы, кроме осетровых, рептилии, птицы) и все мезолецитальные яйца (осетровые рыбы, амфибии).
- Гомо (изо)- лецитальные — желток распределён равномерно. Сюда относятся олиголецитальные ядра (моллюски, иглокожие, млекопитающие).Из гомолицетальных яйцеклеток, в ходе эволюции появились алицетальные яйцеклетки — без желтка. Такие клетки характерны для человека.
- Центролецитальные — желток расположен в центре яйцеклетки. Сюда относятся некоторые полилецитальные яйца (членистоногие). Это совершенно особый тип яиц. Анимально-вегетативная полярность этих яиц не выражена, так как место выделения редукционных телец может быть различным. Вместо анимального и вегетативного полюсов у этих яиц говорят о переднем и заднем полюсах. В центре яйца расположено ядро, а по периферии — ободок свободной от желтка цитоплазмы. Оба этих района — центр и периферия яйца — связаны тонкими цитоплазматическими мостиками, а всё промежуточное пространство заполнено желтком.
14) Наследственная изменчивость как свойство живого. Структурно-функциональные уровни генетического материала. Строение и функции ДНК. Процессы в которых участвует ДНК.
Непрерывность существования и историческое развитие живой природы обусловлены двумя фундаментальными свойствами жизни: наследственностью и изменчивостью На клеточном и организменном (онтогенетическом) уровнях организации живого под наследственностью понимают свойство клеток или организмов в процессе самовоспроизведения передавать новому поколению способность к определенному типу обмена веществ и индивидуального развития, в ходе которого у них формируются общие признаки и свойства данного типа клеток и вида организмов, а также некоторые индивидуальные особенности родителей. На популяционно-видовом уровне организации жизни наследственность проявляется в поддержании постоянного соотношения различных генетических форм в ряду поколений организмов данной популяции (вида). На биоценотическом уровне продолжительное существование биоценоза обеспечивается сохранением определенных соотношений видов организмов, образующих этот биоценоз.
В ходе возникновения и развития жизни на Земле наследственность играла решающую роль, так как закрепляла в ряду поколений биологически полезные эволюционные приобретения, обеспечивая определенный консерватизм организации живых систем. Наследственность является одним из главных факторов эволюции.
Продолжительное существование живой природы во времени на фоне меняющихся условий было бы невозможным, если бы живые системы не обладали способностью к приобретению и сохранению некоторых изменений, полезных в новых условиях среды. Свойство живых систем приобретать изменения и существовать в различных вариантах называется изменчивостью.
Несмотря на то что по своим результатам наследственность и изменчивость разнонаправлены, в живой природе эти два фундаментальных свойства образуют неразрывное единство, чем достигается одновременно сохранение в процессе эволюции имеющихся биологически целесообразных качеств и возникновение новых, делающих возможным существование жизни в разнообразных условиях.
ДНК состоит из нуклеотидов, в состав которых входят сахар — дезоксирибоза, фосфат и одно из азотистых оснований — пурин (аденин или гуанин) либо пиримидин (тимин или цитозин).
Особенностью структурной организации ДНК является то, что ее молекулы включают две полинуклеотидные цепи, связанные между собой определенным образом.
Ген-эллементарная единица генетического аппарата. Основные свойства которого определяются его химической организацией. Нуклеиновые кислоты – материальный субстрат наследственности и изменчивости, состоит из нуклеотидов, кот. В свою очередь состоят из пентозы, фосфата и азотистых оснований. Функции ДНК-хранение генетической информации, передача, удвоение. Репликация-удвоение ДНК, способ полуконсервативный. Репарация- исправление ошибок. Есть дореплекативная и постреплекативная(световая репарация, SOS-система). Транскрипция переписывание информации с ДНК на иРНК с помощбю ДНК-полимеразы. Мутирование-нескоректированные изменения химеческой структуры генов, воспроизведение в последующих циклах репликации и проявляющаяся у потомства в виде новых вариантов признаков.
15)
15. Генный уровень организации наследственного материала. Ген. Определение строение генов про и эукариот. Свойства гена.
на генном уровне организации наследственного материала обеспечиваются индивидуальное наследование и индивидуальное изменение отдельных признаков и свойств клеток, организмов данного вида.
Реальное существование генного уровня организации наследственного материала дало возможность исследователям при анализе характера наследования отдельных признаков открыть главные закономерности, которые легли в основу наших представлений об организации материального носителя наследственности и изменчивости.
ГЕН-лементарная единица генетического аппарата.Осн свойства гена опр-ся его хим организацией. Исследования, направленные на выяснение химической природы наследственного материала, неопровержимо доказали, что материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты, которые были обнаружены Ф. Мишером (1868) в ядрах клеток гноя. Нуклеиновые кислоты являются макромолекулами, т.е. отличаются большой молекулярной массой. Это полимеры, состоящие из мономеров — нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание (пурин или пиримидин). К первому атому углерода в молекуле пентозы С-1' присоединяется азотистое основание (аденин, гуанин, цитозин, тимин или урацил), а к пятому атому углерода С-5' с помощью эфирной связи — фосфат; у третьего атома углерода С-3' всегда имеется гидроксильная группа — ОН (рис. 3.1).
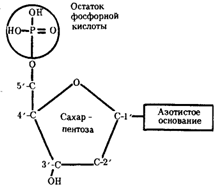
Рис. 3.1. Схема строения нуклеотида
Сборка полинуклеотидной цепи осуществляется при участии фермента полимеразы, который обеспечивает присоединение фосфатной группы следующего нуклеотида к гидроксильной группе, стоящей в положении 3', предыдущего нуклеотида. Благодаря отмеченной специфике действия названного фермента наращивание полинуклеотидной цепи происходит только на одном конце: там, где находится свободный гидроксил в положении 3'. Начало цепи всегда несет фосфатную группу в положении 5'. Это позволяет выделить в ней 5' и 3 '-концы.
Ген-участок молекулы ДНК,отвеч за синтез 1 полипептида
1. дискретность-ген опр возможность развития отдельного качества,присущего данной клетке или организму.
2. специфичным –т.к. в гене заключается информация об аминокислотной последовательности определенного полипептида,
3.плейотропность-участие его продукта-полипептида в разных биохим процессах,имеющих отношение к формированию различных сложных признаков.
4. дозированностью действия, т.е. количественной зависимостью результата его экспрессии от дозы соответствующего аллеля этого гена
5. аллельность-наличие нескольких вариантов формирования признака.
6.стабильность — способность сохранять структуру;
7.лабильность — способность многократно мутировать
У про и эукариот почти всё одинаково, кроме…
Наследственный материал прокариотической клетки содержится главным образом в единственной кольцевой молекуле ДНК. Она располагается непосредственно в цитоплазме клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты, часть из которых заключена в рибосомах. Гены прокариот состоят целиком из кодирующих нуклеотидных последовательностей, реализующихся в ходе синтеза белков, тРНК или рРНК.
Наследственный материал эукариот больше по объему, чем у прокариот Он расположен в основном в особых ядерных структурах —хромосомах ,которые отделены от цитоплазмы ядерной оболочкой. Необходимый для синтеза белков аппарат, состоящий из рибосом, тРНК, набора аминокислот и ферментов, находится в цитоплазме клетки.
Значительные отличия имеются в молекулярной организации генов эукариотической клетки. В большинстве из них кодирующие последовательности экзоны(кодирующие) прерываются интронными(некодирующие) участками, которые не используются при синтезе тРНК, рРНК или пептидов.
У эукариот гены «захломлены интронами, а у прокариот» есть лиш экзоны.
16. Свойства гена как функциональной единицы наследственности и изменчивости. Понятие аллеля и множественного аллелизма.
Основные свойства гена как функциональной единицы материала наследственности и изменчивости определяются его химической организацией.
Исследования химической природы наследственного материала, неопровержимо доказали, что материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты. Это полимеры, состоящие из мономеров-нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание. Среди нуклеиновых кислот различают два вида соединений: дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК) кислоты. Более химически устойчивым компонентом является ДНК, которая и представляет собой субстрат наследственности и изменчивости. Согласно модели Д. Уотсона и Ф. Крика, молекула ДНК представляет собой замкнутую спиралевидную двойную цепь, звенья которой составляют нуклеотиды. Каждый нуклеотид состоит из сахара - дезоксирибозы, фосфорной кислоты и одного азотистого - пуринового (аденин, гуанин) или пиримидинового (цитозин, тимин) основания. Азотистое основание в каждом нуклеотиде соединено с молекулой дезоксирибозы. Все нуклеотиды связаны между собой через дезоксирибозу и остатки фосфорной кислоты. Следовательно, нить ДНК состоит из чередующихся молекул оксирибозы, фосфорной кислоты и азотистых оснований. Нити ДНК соединены друг с другом водородными атомами через азотистые основания так, что против пуринового основания всегда располагается пиримидиновое. Такими парами являются аденин и тимин, гуанин и цитозин. Эти пары оснований называются комплементарными. Одним из основных свойств материала наследственности является его способность к самокопированию - репликации.
Аллели (аллеломорфы)— различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологичных хромосом; определяют варианты проявления одного и того же признака. В диплоидном организме может быть два одинаковых аллеля одного гена, в этом случае организм называется гомозиготным, или два разных, что приводит к гетерозиготному организму.
Множественный аллелизм — это существование в популяции более двух аллелей данного гена. В популяции оказываются не два аллельных гена, а несколько. Возникают в результате разных мутаций одного локуса.
17. Этапы реализации генетической информации. Транскрипция и процессинг РНК.
Этапы реализации ген инфы:
1. Транскрипция - биосинтез молекул РНК на соответствующих участках ДНК, первый этап реализации генетической информации в живых клетках. Осуществляется с помощью специального фермента - РНК-полимеразы.
Транскрипция-переписывание наследственной информации с участка молекулы ДНК(репликона) на иРНК. Синтез начинается с обнаружения ДНК-полимеразой промотора. Синтез иРНК происходит с соблюдением комплиментарности и антипараллельности по отношению к матричной цепи ДНК(3’-5’).При встрече с терминатором РНК-полимераза отделяется от матрицы ДНК и от иРНК. Участок содержащий промотор,транскрибируемую последовательность и терминатор называют транскриптоном.Далее происходит процессинг-«созревание» иРНК. С помощью спец ферментов интроны-не несущие генной информации вырезаются, а остаются лишь экзоны.
К настоящему времени считается установленным, что процессинг мРНК включает три основных процесса: 1) кэпирование – химическая модификация 5'-концевой последовательности мРНК; 2) сплайсинг – удаление некодирующих интронных последовательностей из мРНК и сшивание образующихся экзонов; 3) полиаденилирование – химическая модификация 3'-концевой последовательности мРНК
…………….Не обязательно…..
(Стартовой точкой транскрипции служит основание ДНК, соответствующее основанию РНК, которое первым включается в транскрипт. Транскрипция мРНК продолжается до тех пор, пока РНК-полимеразе II не встретится сигнал терминации транскрипции. Первичный транскрипт мРНК расщепляется специфической эндонуклеазой сразу за последовательностью нуклеотидов АААУАА и к З'-концу мРНК пришивается с помощью поли-А-полимеразы хвост из 100—200 остатков аде-нина, которые, по-видимому, защищают мРНК от деградации во время ее передвижения из ядра в цитоплазму. Кроме того, вскоре после начала синтеза мРНК ее 5'-конец «копи-руется», т.е. к нему присоединяется модифицированный нук-леотид — 5-метилцитозин. Кэппирование происходит в месте начала транскрипции. Обычно транскрипция продолжается, и мРНК удлиняется еще на несколько тысяч пар нуклеотидов, комплементарных ДНК, после чего молекула мРНК отъединяется от матрицы ДНК и РНК-полимеразы. Прежде чем такая мРНК попадет в цитоплазму, она должна созреть (см. рис. 3.7).
Созревание, или процессинг, заключается в том, что из первичного транскрипта мРНК вырезаются (сплайси-руются) специальными ферментами участки гена, транскрибированные в мРНК, которые содержат обычно (но не всегда) некодирующие последовательности1. Эти некодирующие участки имеются практически во всех генах высших организмов и называются интронами. От молекулы мРНК отрезаются также последовательности нуклеотидов за поли-А-хвос-том. Кодирующие участки гена называют экзонами. Экзоны гена, представленные в мРНК, соединяются с помощью специальных ферментов вместе, образуя функционально зрелую мРНК. Именно последовательность кодонов мРНК кодирует последовательность аминокислот в белке, который будет создаваться на ее основе. Зрелая мРНК перемещается в цитоплазму. )
2. Трансляция в биологии, процесс биосинтеза полипептидных цепей белков в живых клетках. Заключается в "считывании" генетической информации, "записанной" в виде последовательности нуклеотидов в молекулах информационных (матричных) рибонуклеиновых кислот (иРНК, или мРНК).
18. Этапы реализация генетической информации, трансляция и посттрансляционные изменения белка.
Этапы реализации ген инфы
1) Транскрипция - биосинтез молекул РНК на соответствующих участках ДНК, первый этап реализации генетической информации в живых клетках. Осуществляется с помощью специального фермента - РНК-полимеразы.
2) . Трансляция-осуществляется с помощью тРнк. С одной стороны к ней присоединяется антикодон, а с другой кодируемая им Аминокислота. С помощью фермента амиоацил-тРНК-синтетазы происходит переписывание информации с языка кодонов на язык Аминокислот. Процесс переписывания происходит на рт=ибосомах. Происходит сборка больших и малых субъединиц рибосом т РНК переходит последовательно из А в П участок.Можно выделить 3 фазы:1)инициации-сборка малой и большой субъединиц рибосом и присоединение к ней первой аминоацил-тРНК.2)Фаза элангации –удлинение пептида, происходит специфичное узнавание аминоацил тРНК, очередного кодона, находящегося в А участве, комплементарное взаимодействие между кодоном и антикотдоном. 3)Фаза терминации-узнавание специфическим рибосомным белком одного из терминирующих кодонов, при этом к последней аминокислоте в пептидной цепи присоединяется вода.
Наследственная информация, «записанная» в молекулах ДНК и «переписанная» на мРНК, расшифровывается в
ходе трансляции благодаря двум процессам специфического узнавания молекулярных поверхностей. Сначала
фермент аминоацил-тРНК-синтетаза обеспечивает соединение тРНК с транспортируемой ею аминокислотой.
Затем аминоацил-тРНК комплементарно спаривается с мРНК благодаря взаимодействию антикодона с
кодоном. С помощью системы тРНК язык нуклеотидной цепи мРНК. транслируется в язык аминокислотной
последовательности пептида (рис. 3.30).
Посттрансляционные преобразования белков:после трансляции белки приобретают первичную,вторичную,третичную и четвертичную структуры.Многие белки синтезируются в виде пре-белков(пре-инсулин).
19. Основные этапы реализации генетической информации. Особенности экспрессии гена у про и эукариот.
1) Транскрипция - биосинтез молекул РНК на соответствующих участках ДНК, первый этап реализации генетической информации в живых клетках. Осуществляется с помощью специального фермента - РНК-полимеразы.
2) Трансляция в биологии, процесс биосинтеза полипептидных цепей белков в живых клетках. Заключается в "считывании" генетической информации, "записанной" в виде последовательности нуклеотидов в молекулах информационных (матричных) рибонуклеиновых кислот (иРНК, или мРНК).
на каждом этапе экспрессии генетической информации можно отметить некоторые особенности течения этих процессов у про- и эукариот.
Транскрипция у про- и эукариот. Транскрипция — это синтез РНК на матрице ДНК. У прокариот синтез всех трех видов РНК катализируется одним сложным белковым комплексом — РНК-полимеразой.
Транскрипционный аппарат эукариотических клеток включает три ядерные РНК-полимеразы, а также РНК-полимеразы митохондрий и пластид. Ферментный комплекс РНК-полимеразы специфически узнает некую нуклеотидную последовательность (часто не одну), расположенную на определенном расстоянии от стартовой точки транскрипции, — промотор. Стартовой точкой считают нуклеотид ДНК, которому соответствует первый нуклеотид, включаемый ферментом в РНК-транскрипт.
После установления контакта между РНК-полимеразой и промоторным участком начинается сборка молекулы РНК,
В связи с тем что прокариотические гены целиком состоят из нуклеотидных последовательностей, участвующих в кодировании информации, транскрибированные с них РНК сразу после их синтеза способны выполнять функцию матриц для трансляции. Лишь в исключительных случаях требуется их предварительное созревание — процессинг.
В отличие от прокариотических генов большинство генов эукариотических клеток прерывисты, так как несут в своем составе неинформативные нуклеотидные последовательности — интроны, не участвующие в кодировании информации. В связи с этим первичные транскрипты, синтезированные РНК-полимеразой II, обладают большими, чем необходимо для трансляции, размерами и оказываются менее стабильными. В совокупности они образуют так называемую гетерогенную ядерную РНК(тяРНК), которая прежде чем выйти из ядра и начать активно функционировать в цитоплазме, подвергается процессингу и превращается в зрелые мРНК.
Процессинг эукариотических мРНК.преобразования первичных транскриптов эукариотических генов, обусловленные их экзонитронной организацией и необходимостью перехода мРНК из ядра в цитоплазму, определяют особенности реализации генетической информации в эукариотической клетке.
Трансляция у про- и эукариот. В прокариотических клетках процесс трансляции сопряжен с синтезом мРНК:
У эукариот трансляция осуществляется в цитоплазме, куда попадает из ядра зрелая мРНК. Синтез белка в эукариотической клетке, осуществляемый на моноцистронной мРНК, завершается после прохождения рибосомой по всей мРНК, вплоть до узнавания ею кодона-терминатора, прекращающего образование пептидных связей.
20. Генные мутации. Определение. Классификация. Мутон. Возможные механизмы возникновения и последствия генных мутаций. Примеры у человека
Генные мутации — нескорректированные изм. хим. структуры генов, воспроизводимые в последовательных циклах репликации и проявляющиеся у потомства в виде новых вариантов признаков.
Мутации по типу замена азотистых оснований. Эти мутации происходят в силу причин: одной из них может быть возникающее случайно или под влиянием конкретных хим. агентов изменения структуры основания, уже вкл. в спираль ДНК. Если такая измененная форма основания остается не замеченной ферментами репарации, то при ближайшем цикле, репликации она может присоединять к себе др. нуклеотид.
Мутации со сдвигом рамки считывания. Этот тип мутации составляет значительную долю спонтанных мутаций. Они происходят вследствие выпадения или вставки в нуклеотидную последовательность ДНК одной или нескольких пар комплементарных нуклеотидов.
Мутации по пути инверсии нуклеотидных последовательностей в гене. Данный тип мутации происходит вследствие поворота участка ДНК на 1800. Этому предшествует образование молекулой ДНК петли, в пределах которой репликация идет в направлении, обратном правильному.
Мутон-минимальное количество наследственного материала,способное,изменяясь,приводить к появлениювариантов признаков,и соответствует элементарной единице мутационного процесса.
Механизмы возникновения генных мутаций-неравноценный обмен при кроссинговере,т.е. при нарушенииирекомбинации между ними. Это приводит к сдвигу рамки считывания и приводит к нарушению синтеза пептидной цепи с заданными свойствами.
Дата добавления: 2018-08-06; просмотров: 246; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!