Основной мишенью в клетке является клеточное ядро, а точнее — молекулы ДНК.
Кривые выживаемости клеток. Принцип попадания и теория мишени Дозовые зависимости гибели и выживаемости многоклеточных организмов, как мы это уже рассматривали на примере млекопитающих, принято изображать соответственно в координатах "% погибших организмов — доза облучения" и "% выживших организмов — доза облучения": В отличие от этого в клеточной радиобиологии применяют обычно только кривые выживаемости, причем построенные в полулогарифмическом масштабе, а именно с использованием логарифмической шкалы ординат. Кривая выживаемости клеток в обычных (т.е. линейных) координатах: Кривая выживаемости клеток в полулогарифмическом масштабе (с использованием логарифмической шкалы ординат): В общем виде кривая выживаемости клеток в полулогарифмическом масштабе состоит из двух участков: т.н. "плеча" и линейного участка. Величина плеча зависит от 1) вида излучения и 2) типа клеток. Типичные кривые выживаемости могут наблюдаться при облучении только редкоионизирующим излучением (рентгеновским или γ-излучении). При облучении плотноионизирующим излучением кривые выживаемости имеют слабовыраженное плечо или вовсе его не имеют (например, в случае α-облучения): в обычных (т.е. линейных) координатах: в полулогарифмическом масштабе: Такие же кривые выживаемости, не имеющие плеча, наблюдаются и при облучении некоторых типов клеток редкоионизирующим излучением. Количественно кривые выживаемости, не имеющие плеча, описываются уравнением: или , где N0 — исходное количество клеток, N — количество клеток, выживших (т.е. сохранивших колониеобразующую способность) после облучения в дозе D, k — коэффициент, определяющий наклон прямой (чем выше k, тем круче наклон). В случае наличия плеча кривые выживаемости описываются уравнением: , где n — т.н. экстраполяционное число — значение ординаты в месте пересечения с экстраполированным прямолинейным участком кривой выживаемости: Наличие подобного рода зависимости выживаемости клеток от дозы облучения было продемонстрировано еще в 20-е годы ХХ столетия. Уже в то время было отмечено, что кривые выживаемости клеток после воздействия излучения отличаются от кривых выживаемости клеток после воздействия каких-либо токсических агентов химической природы, например ядов. Кривые выживаемости клеток, получаемые при действии химических агентов, характеризуются наличием некоторой пороговой концентрации, ниже которой все клетки выживают, а после превышения которой происходит резкое снижение доли выживших. Переход от 100%-ной выживаемости до нулевой выживаемости происходит в очень узком диапазоне концентраций химического агента. S-образную форму кривой выживаемости клеток после действия агентов химической природы объясняют обычно с т.н. "биологических" позиций, а именно исходя из наличия индивидуальных различий клеток в клеточной популяции, подвергнутой этому воздействию. Чем меньше выражены индивидуальные различия, тем круче наклон кривой выживаемости. Однако, на кривых выживаемости клеток, подвергнутых облучению, отсутствует четкий порог: гибель части клеток наблюдается даже при очень низких дозах облучения, а гибель другой части клеток той же популяции — лишь при значительно более высоких дозах. Объяснение подобной формы кривой выживаемости облученных клеток с "биологических" позиций звучало в начале прошлого века весьма неубедительно, т.к. трудно было представить, что внутри генетически однородной популяции могут находиться клетки, различающиеся по устойчивости к облучению в сотни и тысячи раз. Поэтому в 20-е годы прошлого века в противовес "биологическому" подходу сформировался "физический" (или "биофизический") подход к объяснению и интерпретации кривых выживаемости клеток, подвергнутых действию ионизирующего излучения. В основе это направления лежала "теория точечного тепла", разработанная Дессауэром (Dessauer) в 1922 г. Эта теория опиралась на следующие основные идеи: Ø несмотря на то, что плотность поглощенной энергии в облучаемом объекте в среднем очень низкая, энергия самих актов поглощения весьма велика и поэтому в микрообъемах вещества, в которых произошли акты поглощения энергии, происходит его локальное нагревание (т.е. появление т.н. "точечного тепла"), в результате чего вещество претерпевает значительные локальные изменения, обусловленные разрывом химических связей или активацией химических реакций; Ø клетка гетерогенна (т.е. неоднородна) по своему объему в отношении чувствительности к облучению и имеет как исключительно важные для жизни области, повреждение которых приводит клетку к гибели, так и области, относительно несущественные для выживания клетки; Ø пространственное распределение появления "точечного тепла" внутри облученной клетки имеет случайный характер, т.е является чисто статистической (т.е. вероятностной) функцией. Т.о. эта теория заявила, что конечный эффект в клетке (например, гибель) определяется вероятностью осуществления акта поглощения энергии (т.е. появления "точечного тепла") в жизненно важных микрообъемах (областях) клетки. При увеличении дозы облучения вероятность осуществления акта поглощения энергии в этих клеточных областях возрастает, при снижении дозы — уменьшается. Однако, даже при очень высокой дозе облучения существует определенная вероятность того, что в каком-то количестве клеток не произошло актов поглощения энергии в жизненно важных клеточных областях и поэтому эти клетки останутся неповрежденными облучением. И наоборот — даже при очень низкой дозе существует вероятность того, что в части клеток произойдут акты поглощения энергии в жизненно важных клеточных областях, что вызовет гибель этих клеток. В дальнейших работах Дж. Кроутера (J.A. Crowther; в русскоязычной литературе встречаются также и другие варианты написания фамилии этого исследователя — Краузер, Кроузер), Д. Ли (D.E. Lea), К. Циммера (K.G. Zimmer) и Н.В. Тимофеева-Ресовского в 20‑30‑е годы прошлого столетия основные идеи теории точечного тепла получили дальнейшее развитие в результате чего были сформулированы 2 фундаментальные положения в радиобиологии. Первое положение, получившее название "принцип попадания", может быть сформулировано следующим образом: Ø Поглощение энергии в облучаемом объеме происходит в результате дискретных актов взаимодействия квантов излучения с веществом — т.н. "попаданий", пространственное распределение которых в облучаемом объекте имеет случайный характер. Таким образом, "принцип попадания" отмечает дискретность и вероятностный (случайный) характер актов взаимодействия излучения с веществом. Фактически попаданием считается возникновение акта ионизации каких-либо молекул в облучаемом объеме. Второе положение, получившее название "теория мишени", может быть сформулировано следующим образом: Ø Клетка имеет в своем составе как исключительно важные для выживания области — т.н. "мишени", радиационное повреждение которых приводит клетку к гибели, так и области, относительно несущественные для выживания клетки. Таким образом, "теория мишени" отмечает биологическую гетерогенность различных областей (микрообъемов) внутри клетки в отношении чувствительности к облучению и наличие внутри клетки "мишеней" — чувствительных областей, радиационное повреждение которых приводит клетку к гибели. Поражение клеточных мишеней может происходить как при одном единственном попадании в нее — это т.н. одноударные мишени, так и при нескольких попаданиях — это т.н. многоударные мишени. Более того, в клетке возможно наличие нескольких мишеней и гибель клетки может наступить лишь после того, как все они получили определенное число попаданий. В реальности анализ многомишенных кривых гибели требует слишком много допущений и исследования влияния многих параметров и поэтому в большинстве случаев оказывается очень ненадежным. Кроме того принцип попадания и теория мишени не учитывают наличия в клетке репарационных процессов, которые существенно затрудняют интерпретацию кривых выживаемости. Возвратимся к кривым выживаемости облученных клеток и рассмотрим как можно их интерпретировать с позиций принципа попадания и теория мишени для случая одноударной и многоударной одномишенных кривых выживаемости и какими параметрами можно охарактеризовать эти кривые выживаемости. Для этого рассмотрим кривые выживаемости, имеющие плечо и неимеющие плеча. Для кривой выживаемости, неимеющей плеча, важной и фактически единственной характеристикой является величина D37 — доза, при которой выживает 37% облученных клеток. Почему именно 37%? Вернемся к уравнению, описывающему кривую выживаемости, неимеющую плеча: . В этом уравнении при некоторой дозе облучения выполняется условие kD = 1. Это соответствует случаю, когда в среднем число попаданий в одноударные мишени равно числу мишеней. В действительности же при этой дозе облучения часть попаданий происходит в однажды уже пораженные мишени, а некоторые мишени не испытали ни одного попадания. Подставив в это уравнение значение kD = 1, получаем: . Таким образом, при дозе облучения, которая приводит к выживаемости 37% клеток, число попаданий в мишени в среднем равно числу мишеней. Поэтому дозу D37 называют также среднелетальной дозой (не путать с ЛД50!). Для кривой выживаемости, имеющей плечо, имеется несколько важных характеристик: Ø во-первых, это D37 — доза, при которой выживает 37% облученных клеток; Ø во-вторых, доза Dq — доза, соответствующая точке пересечения экстраполированного прямолинейного участка кривой выживаемости с осью абсцисс, проведенной на уровне 100%-ной выживаемости; Dq характеризует величину плеча; Ø в-третьих, доза D0 — определяется как приращение дозы, снижающей выживаемость в е раз на прямолинейном участке кривой выживаемости; служит исходной мерой радиочувствительности; Ø в-четвертых, экстраполяционное число n — значение ординаты в месте ее пересечения с экстраполированным прямолинейным участком кривой выживаемости; в теории мишени экстраполяционное число определяет какое количество попаданий в мишень необходимо для поражения этой мишени. Конечно, в настоящее время ясно, что интерпретация кривых выживаемости клеток чисто с позиций принципа попадания и теории мишени в большинстве случаев не является корректной, так как здесь не учитываются процессы восстановления индуцированных облучением повреждений. Тем не менее, все перечисленные выше характеристики кривых выживаемости широко используются при анализе этих кривых. В частности, величину Dq используют в качестве показателя способности клетки осуществлять репарационные (восстановительные) процессы: чем больше плечо, тем выше репарационная способность клетки. Принцип попадания и теория мишени имеют ряд очень важных практических приложений. Например, на основании их положений возможно определение размера мишени. Это возможно, если процесс инактивации клеток (или процесс инактивации какого-либо фермента при его облучении in vitro) описывается кривой, неимеющей плеча, т.е. является одноударным процессом. В этом случае массу, молекулярную массу и объем мишени можно легко рассчитать по следующим формулам: , , , где ρ — плотность мишени. Рассчитанные таким образом молекулярные массы большого числа ферментов (в данном случае речь идет не об облучении клеток, а об облучении ферментов в водных растворах) очень хорошо совпадали с их молекулярными массами, рассчитанными другими физико-химическими методами. Что же является основной клеточной мишенью при действии радиации?
|
|
|
|
|
|
|
|
|
|
|
|
Основной мишенью в клетке является клеточное ядро, а точнее — молекулы ДНК.
Тот факт, что именно ядро является основной клеточной мишенью при действии радиации, было, в частности, показано в экспериментах по определению размеров мишени в клетке. Оказалось, что размер клеточной мишени примерно совпадает с размером клеточного ядра.
Отсюда следует один из важнейших выводов из принципа попадания и теории мишени: чем выше содержание ДНК, тем чувствительнее клетка к действию ионизирующего излучения.
На представленном ниже рисунке показана корреляция между радиочувствительностью (1/D37) различных биологических объектов (вирусов и клеток) и содержанием в них нуклеиновых кислот (по числу нуклеотидов). [См. красную линию]






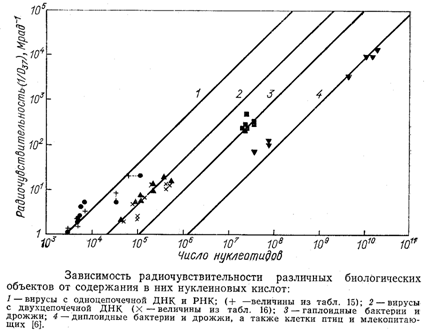
Корреляция между радиочувствительностью и количеством нуклеотидов в нуклеиновой кислоте наблюдается особенно четко, если эти биологические объекты сгруппировать в 4 группы в зависимости от структурной организации их нуклеиновых кислот и проанализировать эту корреляцию внутри каждой из них. Это следующие группы:
1) группа вирусов с одноцепочечной ДНК и РНК,
2) группа вирусов с двуцепочечной ДНК,
3) группа гаплоидных бактерий и дрожжей,
4) группа диплоидных бактерий и дрожжей, а также клеток птиц и млекопитающих.
В каждой из этих групп — чем больше число нуклеотидов, тем выше радиочувствительность.
Интересно также обратить внимание, что при одном и том же числе нуклеотидов вирусы с двуцепочечной ДНК примерно на порядок более устойчивы, чем вирусы с одноцепочечной ДНК и РНК, а диплоидные клетки примерно на порядок более устойчивы, чем гаплоидные клетки. Это, в частности говорит о том, что наличие двух цепочек в молекуле ДНК и двойного набора хромосом (т.е. двух копий каждой из молекул ДНК) повышает надежность биологического объекта и его устойчивость к облучению.
Предполагают, что второй по значению мишенью в клетке являются биологические мембраны. Однако, большинство исследователей всё же считает это утверждение недостаточно доказанным.
Дата добавления: 2018-05-12; просмотров: 668; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!